大珠母贝胰岛素相关肽受体基因的克隆与表达
2020-07-24赵子涵王姿曼邓岳文杨创业李俊辉
赵子涵,王姿曼,邓岳文,2,杨创业,2,李俊辉
大珠母贝胰岛素相关肽受体基因的克隆与表达
赵子涵1,王姿曼1,邓岳文1,2,杨创业1,2,李俊辉1
(1. 广东海洋大学水产学院 // 2. 广东省珍珠科技创新中心,广东 湛江 524025)
【】克隆大珠母贝()胰岛素相关肽受体基因,并分析其在不同组织中的表达。利用cDNA快速末端克隆技术(RACE)获得大珠母贝基因cDNA全长序列。荧光定量PCR技术分析在闭壳肌、鳃、外套膜边缘区、套膜区、中央区、足和肝胰腺等组织的表达模式。【】基因全长6 009 bp,5′非编码区(UTR)615 bp、3′UTR 764 bp、开放阅读框(ORF)长4 629 bp、编码 1 542个氨基酸。序列分析表明,含有保守结构域Fu、3个FNⅢ结构域和Tyrkc结构域,并含有信号肽和2个跨膜结构域。在大珠母贝各个组织存在差异表达,在鳃、肝胰腺和外套膜边缘区中显著高表达。
大珠母贝;胰岛素肽相关受体;基因克隆;表达模式
胰岛素样家族是动物生长、发育和代谢的重要调节因子,通过与酪氨酸激酶受体(胰岛素受体)结合,在传递跨膜信息和调节细胞功能方面发挥重要作用,参与许多生命活动[1-2]。胰岛素相关受体()是胰岛素受体()家族的成员,作为细胞表面受体,是参与调节体内酸碱平衡的细胞外碱性介质代谢传感器,在肾、胃和胰腺的某些细胞群中表达[3-4]。脊椎动物有不止一个同源的胰岛素受体家族成员,而无脊椎动物只有一个胰岛素相关肽受体(),这个单一的调节无脊椎动物的生长和代谢[1,5]。胰岛素相关肽及其受体已在许多软体动物中报道,Hamano等[6]在太平洋牡蛎中鉴定分离了胰岛素相关肽基因,并分析其结构和表达。在太平洋牡蛎中,胰岛素系统调控外套膜生长和壳形成有关的内外上皮细胞水平,还可通过饵料调控整合环境条件的变化[7-8]。马氏珠母贝()中胰岛素相关肽受体()与人重组胰岛素样生长因子I()相互作用,并刺激了卵母细胞的MAPK和PI3K信号通路调节糖原代谢[4]。Lardans等[9]也探讨了曼氏血吸虫幼虫共培养类胰岛素受体()在曼氏血吸虫幼虫胚胎细胞的激活和增殖过程中的可能意义。
大珠母贝()自然分布在澳大利亚、菲律宾、马来西亚、印度尼西亚等热带亚热带国家沿海地区,在我国广东省、广西省和海南省也有分布,是我国南方海水养殖珍珠贝类的重要组成部分之一,主要用于生产大规格珍珠[10-11]。目前大珠母贝产业主要问题为养殖期间苗种成活率偏低,无法为商业化生产提供足量母贝[12]。通过品种培育与养殖技术研究,能明显提高养殖成活率。本实验利用RACE技术克隆了基因的全长序列,并通过荧光定量PCR技术检测了在不同组织的表达模式,为进一步了解在大珠母贝中生长作用提供资料。
1 材料与方法
1.1 材料
实验用贝取自湛江市徐闻县承梧村海区,挑选无病害的2龄大珠母贝,剪取闭壳肌(A)、鳃(GI)、外套膜边缘区(ME)、套膜区(MP)、中央区(MC)、足(F)和肝胰腺(HE),所取组织放入液氮速冻,存于-80 ℃超低温冰箱,用于荧光定量分析。
RNA提取试剂Trizol,SMARTer TMRACEcDNA Amptification kit,SYBR Premix ExTaqTM购自Thermo Fisher Scientific公司,Reverse Transcriptase M-MLV (RnaseH)、rTaq和Primerstar DNA聚合酶、PDM-19T载体购自Takala Bio公司。
1.2 PmIRR基因全长的克隆
1.2.1 总RNA的提取和cDNA第一链的合成 用Trizol提取法提取大珠母贝组织总RNA,利用NanoDropND1000紫外分光光度计检测RNA浓度和纯度,1%琼脂糖凝胶电泳检测其完整性。5′RACE和3′RACE模板的获得参考SMARTerTM RACE cDNA Amolification Kit[13]。
1.2.2序列全长获取 根据大珠母贝转录组数据库中的unigene序列设计基因特异性引物,将该基因中间分为三段克隆。PCR扩增产物通过10 g/L的琼脂糖凝胶电泳检测,并用纯化回收试剂盒对目的片段进行纯化回收。目的片段通过PDM-19T载体连接,然后转入DH5a 感受态细胞中。通过LA(含氨苄青霉素 Amp+)固体平板筛选出阳性菌落,挑选单克隆[14],送至广州生工生物工程有限公司测序。利用5′端和3′端巢式引物进行扩增。目的片段的回收、转化、测序等方法同基因中间片段,然后将5′端和3′端拼接获得全长序列(表1)。
1.3 PmIRR组织差异表达
用荧光定量PCR检测基因在边缘膜、套膜、中央膜、闭壳肌、腮、肝胰腺、足组织中的表达模式。以为内参基因,配置10 µL反应体系:10 µmol/L上、下游引物各0.4 µL,灭菌的ddH2O 3.8 µL,cDNA0.4 µL,SYBR® Select Master Mix 5 µL反应程序:95 ℃下预变性 5 min,95 ℃下变性 10 s,60 ℃下退火15 s,72 ℃延伸15 s,40循环,添加 1 个溶解曲线。每个样品重复 3 次。用2-△△Ct法计算基因在各组织中的相对表达量。通过SPSS19.0对数据进行单因素方差分析,= 0.05[13]。
1.4 生物信息学分析
利用DNAMAN将中间片段、3′端序列和5′序列拼接起来,获得cDNA全长序列。用在线分析工具ORF Finder 分析开放阅读框(ORF)并推导出基因的氨基酸序列,用ProtParam (http://web.expasy.org/protparam/)分析氨基酸序列的理化性质,运用在线工具SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP/)预测其信号肽。SMART(http://smart.embl-heidelberg.de/)预测PmIRR的保守结构域,用TMHMM Server, v.2.0推测其跨膜结构域,用SOPMA(https://npsa- prabi. ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)推测蛋白质的二级结构。用MEGA6构建PmIRR进化树。用ClusterW2与其它已知物种相应的IRR氨基酸序列进行同源性对比。
表1 引物序列

Table 1 Sequences and functions of primers
2 结果与分析
2.1 基因克隆与特征分析
克隆得大珠母贝cDNA全长序列。结果显示,序列全长6 009 bp,其中 5′UTR 为615 bp;3′UTR 为764 bp,包含28 bp的ployA;ORF为4 629 bp,编码1 542个氨基酸(图1)。经预测PmIRR有一个信号肽位点(27 ~ 28 bp),为分泌蛋白(图2)。两个跨膜结构域位于12-34bp和1005-1027bp,该分子属于膜蛋白(图3)。SMART预测其有1个(Fu)结构域、3个(FN3)结构域和1个(Tyrkc)结构域(图4)。

小写字母和大写字母分别表示非编码区和编码区;加粗字体为起始密码子和终止密码子;双下划线的字母表示信号肽;斜体部分表示跨膜结构域;阴影部分表示Fu结构域;单划线为FN3结构域;方框内为Tyrkc结构域
图1基因的 cDNA 序列及编码的氨基酸序列
Fig. 1cDNA and amino acid sequence of
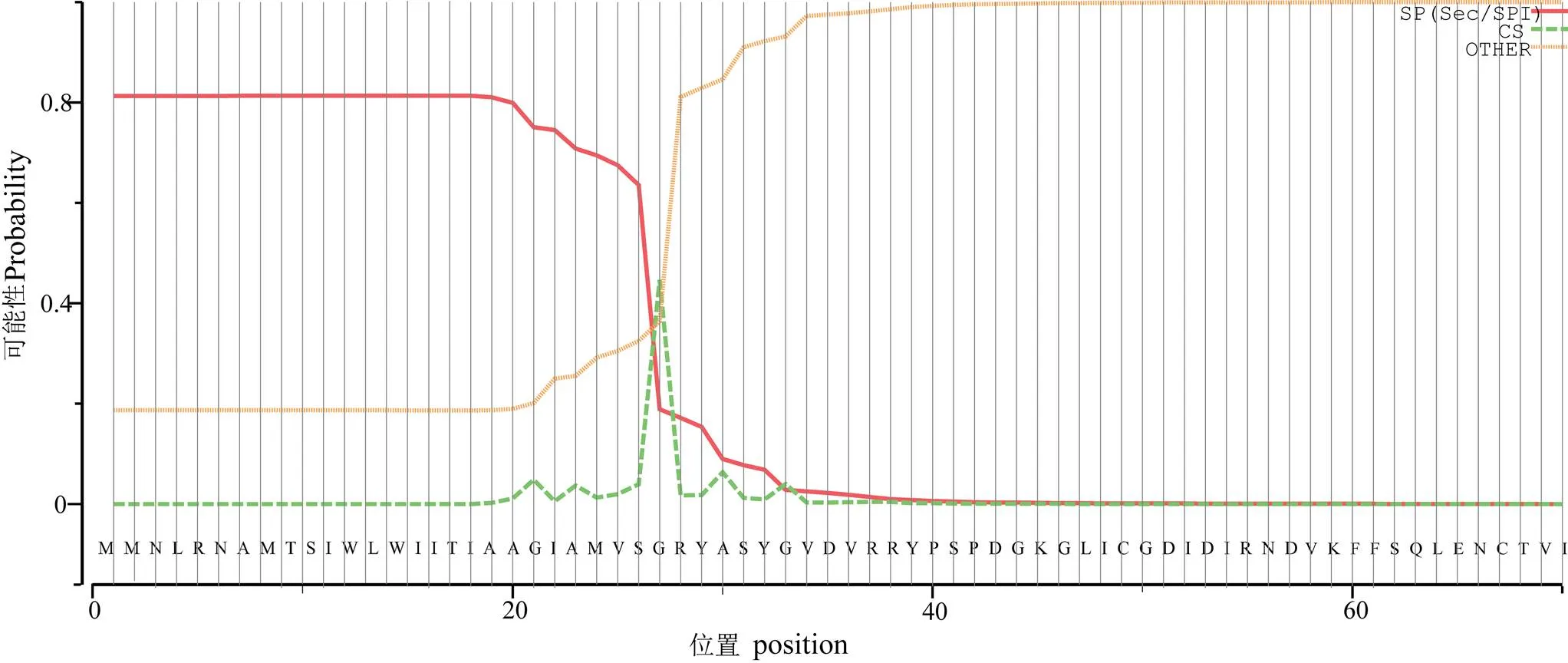
图2 PmIRR信号肽序列
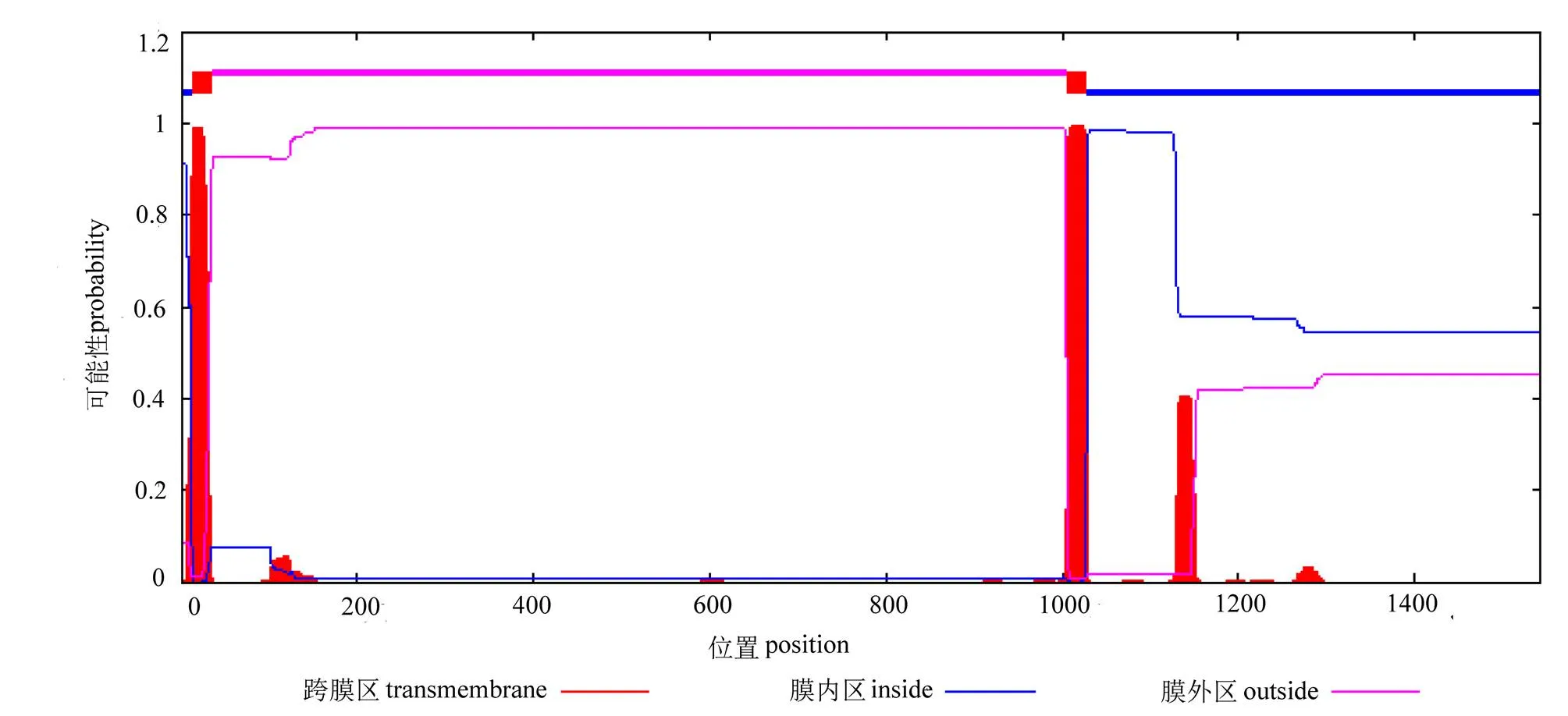
图3 PmIRR跨膜结构域预测
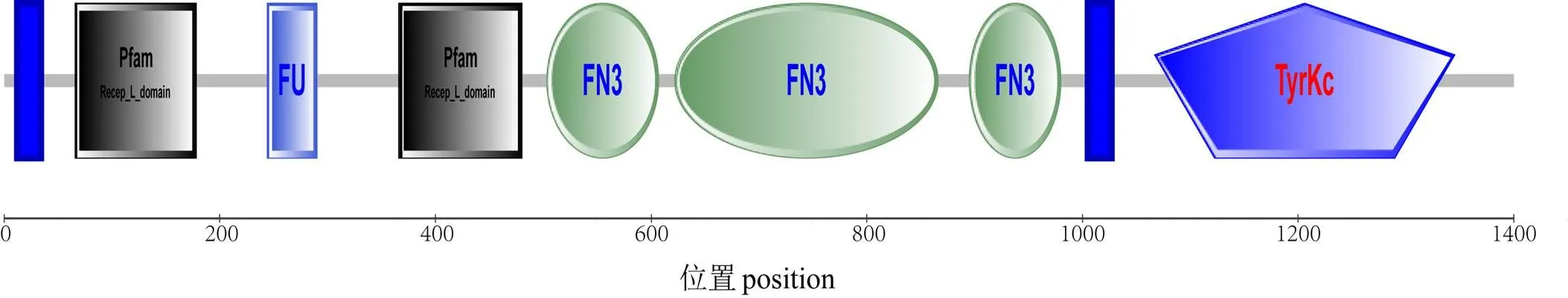
图4 PmIRR蛋白质结构
2.2 PmIRR蛋白质理化性质
预测得PmIRR理论分子质量为75.3 ku;等电点为6.49;负电荷残基202个,正电荷残基192个。氨基酸含量分析结果显示,其脂溶指数(Aliphatic index)为75.89;总平均亲水性(Grand averageofhydropathy, GRAVY)为 – 0.531,属于亲水性蛋白(图5)。经预测,PmIRR的二级结构中,α 螺旋结构占整体的25.36%,延伸链占18.48%,β 转角结构占4.67%,无规则折叠占51.49%(图6)。
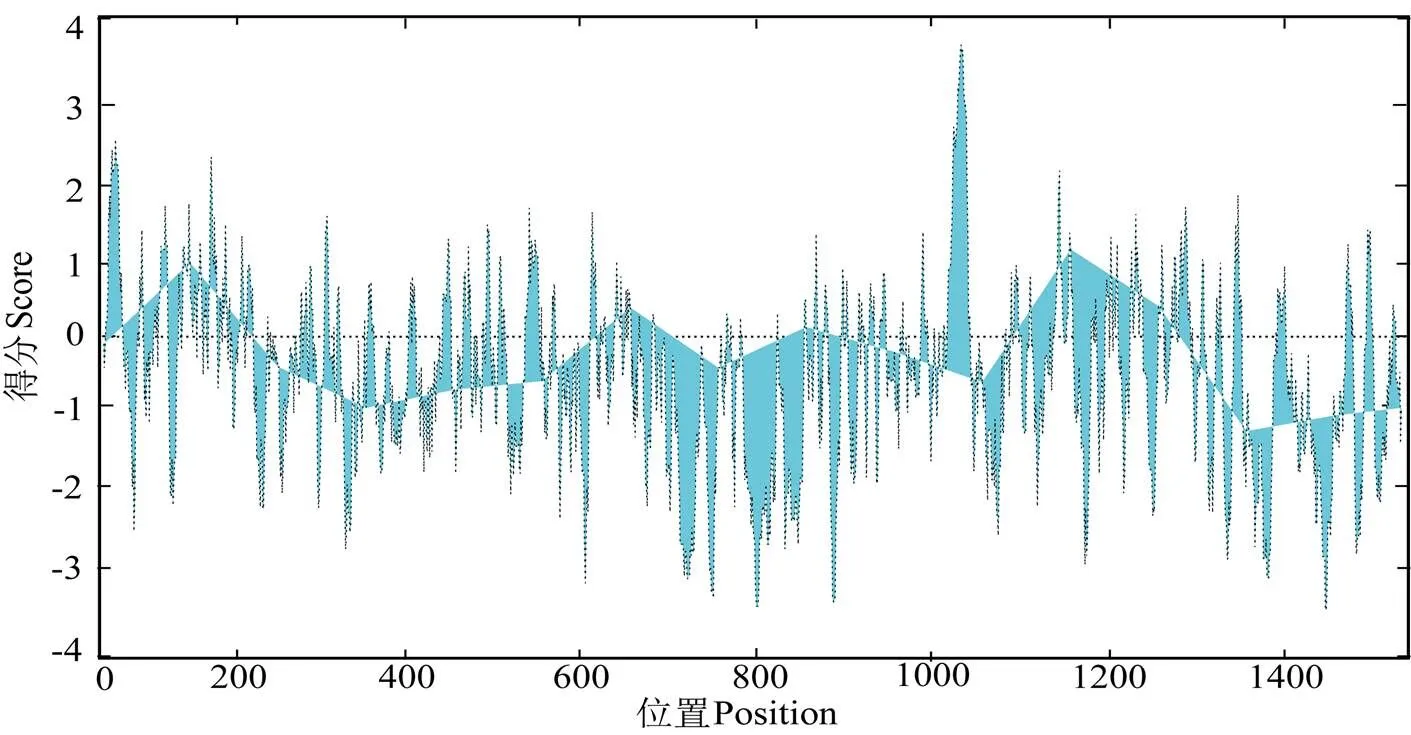
图5 PmIRR蛋白疏水性
图6 PmIRR蛋白二级结构
2.3 同源对比与进化树分析
将美洲牡蛎()、太平洋牡蛎()、马氏珠母贝()和虾夷扇贝()的Tyrkc结构域进行同源性对比(图7),发现大珠母贝PmIRR的Tyrkc结构域与这些物种的Tyrkc结构域相似度较高,说明IRR氨基酸序列有较高保守性。构建PmIRR与其他物种IRR的进化树(图8),发现与双壳贝类聚为一支,跟亲缘性最近,与节肢类动物亲缘关系相对较远。

深蓝色表示一致;粉色表示强相似性;蓝色表示弱相似性
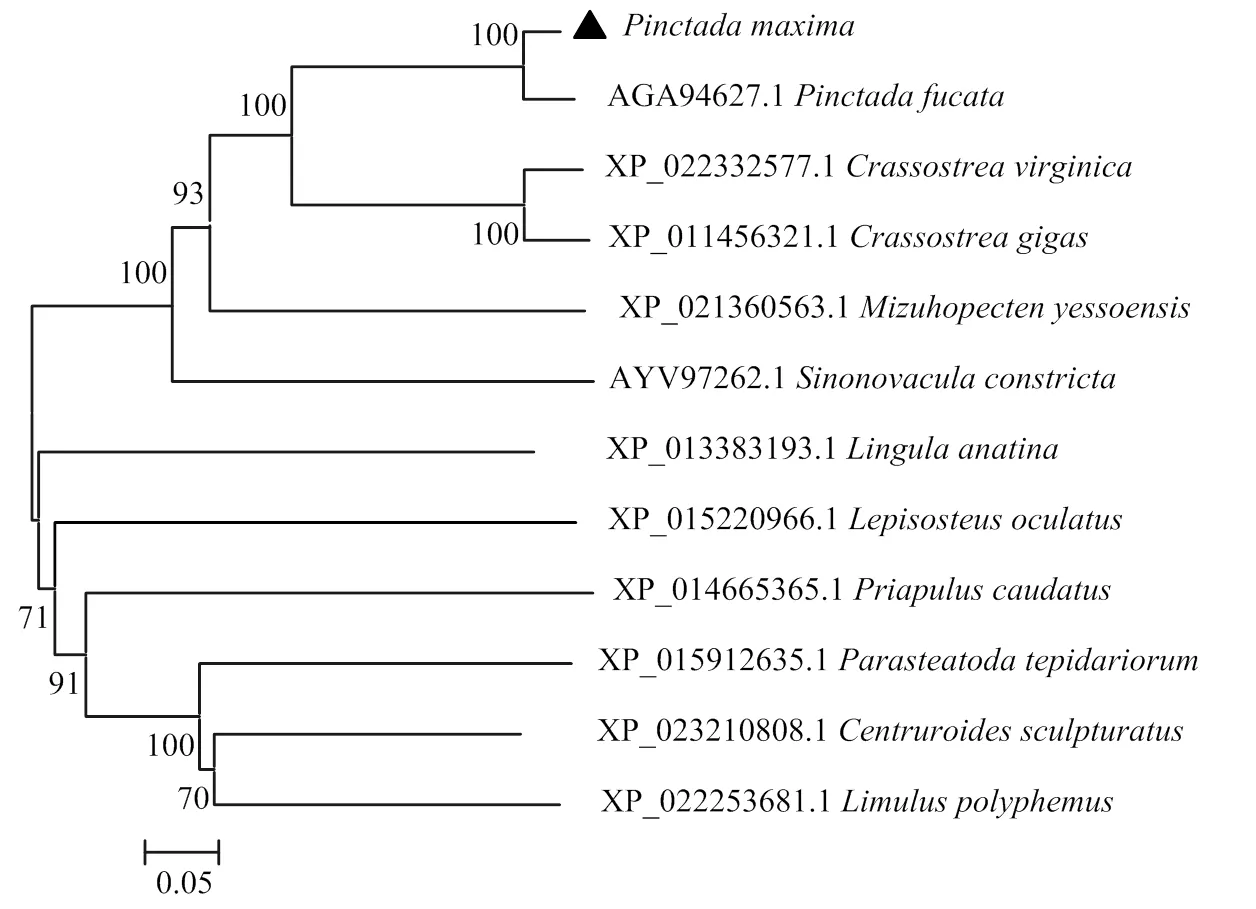
图8 邻接法构建的PmIRR家族系统进化树
2.4 PmIRR基因组织表达模式分析
结果显示,在各个组织中均有表达,其中在鳃、肝胰腺、边缘膜等组织表达量较高,在闭壳肌和足中表达量较低(图9)。
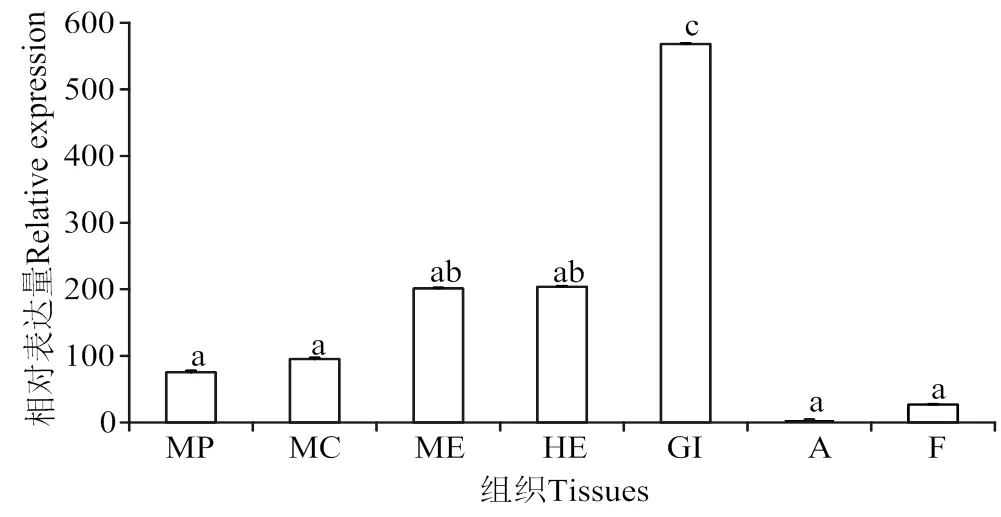
ME,边缘膜;MP,套膜区;MC,中央膜;A,闭壳肌;GI,鳃;HE,肝胰腺;F,足; 无共同字母(a、b、c)表示差异显著(P < 0.05);凡含一个相同字母者表示差异不显著(P > 0.05)
3 讨论
胰岛素相关肽基因及受体在一些软体动物中已有报道,已被证明对生物的生长和发育有重要意义[9,15-17]。为了解胰岛素相关肽受体基因在大珠母贝生长中的作用,本实验利用RACE技术在大珠母贝中克隆得到胰岛素相关肽受体基因的cDNA全长序列。分析发现,该基因氨基酸序列包含1个Tyrkc结构域、3个FN3结构域和1个Fu结构域,多序列比对结果表明其Tyrkc结构域与其他物种的Tyrkc结构域比对结果呈现较高的保守性。Tyrkc结构域为酪氨酸激酶催化结构域在信号传递中发挥重要作用[18-19]。Fu结构域能识别特定氨基酸序列,激活重要的多肽和蛋白的前体[20-21],可能跟IRR的信号传导机制有关。有报道称纤维连接蛋白III型FN3结构域是受体的碱敏感特性的重要区域[22-24]。此外,发现IRR有两个典型的Recep-L-domain结构域,为酪氨酸激酶受体小家族的典型结构域。因此可能发挥胰岛素相关肽受体的功能。
为进一步探究在大珠母贝中的功能,本研究通过荧光定量PCR技术检测其在大珠母贝不同组织中的表达模式,结果显示,在所检测的7个组织中均有表达。其中,在鳃组织中显著高表达,其次是肝胰腺和外套膜边缘区。在这些特定组织中的高表达现象,表明其可能参与机体的能量代谢。在贝类中,鳃组织是呼吸和滤食的主要器官[25]。研究发现胰岛素相关肽()在缢蛏鳃组织中有非常高的表达量,提示胰岛素相关肽可能参与肝糖代谢调控或鳃的再生[26-28]。因此,推测参与大珠母贝鳃组织糖原代谢或细胞再生的调控通路。肝胰腺是贝类重要消化和代谢器官[29],在肝胰腺中高表达可能与其能量代谢的功能相关[5,31]。在外套膜不同区域呈中等表达水平,这与牡蛎[8]的研究结果类似。之前研究已证实外套膜与贝壳的形成有关[32-33]。马氏珠母贝中存在胰岛素相关肽及受体调控通路来调节贝壳的生长和发育[34],表明很可能参与大珠母贝贝壳的生长。郝瑞娟等[35]探究大珠母贝不同步生长代谢机制,确定了双壳类不同步生长中的一些代谢途径和代谢产物。在调控大珠母贝生长发育的作用机制还需进一步研究。
4 结语
本实验从大珠母贝中克隆获得胰岛素肽相关受体()基因cDNA全长序列,发现该基因在大珠母贝鳃、肝胰腺、外套膜边缘区表达量相对较高,可能参与大珠母贝贝壳的生长和机体组织发育等生理功能。本研究为进一步探究在大珠母贝中的生理功能提供基础资料。
[1] SHI Y, GUAN YY, HE M X. Molecular identification of insulin-related peptide receptor and its potential role in regulating development in[J]. Aquaculture, 2013, 408/409:118-127.
[2] BEDINGER D H, ADAMS S H. Metabolic, Anabolic, and Mitogenic Insulin Responses: A Tissue-Specific Perspective for Insulin Receptor Activators[J]. Molecular & Cellular Endocrinology, 2015, 415: 143-156.
[3] MOZHAEV A A, EROKHINA T N, SEROVA O V, et al. Production and immunochemical characterization of monoclonal antibody to IRR ectodomain[J]. Russian Journal of Bioorganic Chemistry, 2017, 43(6): 653-657.
[4] DEVEV I E, MITROFANOVA A V, ZHEVLENEV E S, et al. Structural Determinants of the Insulin Receptor-related Receptor Activation by Alkali[J]. Journal of Biological Chemistry, 2013, 288(47): 33884-33893.
[5] SHI Y, HE M X. PfIRR Interacts with HrIGF-I and Activates the MAP-kinase and PI3-kinase Signaling Pathways to Regulate Glycogen Metabolism in[J]. Scientific Reports, 2016, 6: 22063.
[6] HAMANO K. cDNA structure of an insulin-related peptide in the Pacific oyster and seasonal changes in the gene expression[J]. Journal of Endocrinology, 2015, 187(1): 55-67.
[7] GRICOURT L, BONNC G, BOUJRD D, et al. Insulin-like system and growth regulation in the Pacific oyster: hrIGF-1 effect on protein synthesis of mantle edge cells and expression of an homologous insulin receptor-related receptor[J]. General& Comparative Endocrinology, 2003, 134(1): 44-56.
[8] JOUAUX A, FRANCO A, Heude-Berthelin C, et al. Identification of Ras, Pten and p70S6K homologs in the Pacific oysterand diet control of insulin pathway[J]. Gen Comp Endocrinol, 2012, 176(1): 28-38.
[9] LARDANS V, C0PPINJ F, JEROME V, et al. Characterization of an insulin receptor-related receptor inembryonic cells[J]. Biochimica et Biophysica Acta, 2001, 1510(1/2): 321-329.
[10] DENG Y W, Fu S, Liang F L, Du X D, XieSH. Growth and survival of pearl oysterspat reared under different environmental conditions[J]. Shellfish Res, 2013, 32 (3): 675–679.
[11] WANGZ M, LIANGF L, HUANG R L et al.Identification of the differentially expressed genes ofindividuals with different sizes through transcriptome analysis[J].Regional Studies in Marine Science, 2019, 26: 1-7.
[12] 林伟财, 谢绍河, 杜晓东, 等. 影响大珠母贝育珠效果的主要因素分析[J]. 中国农学通报, 2016, 32(32): 29-33.
[13] HAO R J, ZHENG Z, WANG Q H, et al, et al. Molecular and functional analysis of PmCHST1b in nacre formation of[J]. Comparative Biochemistry and Physiology. Part B, Biochemistry &Molecular Biology, 2018, 225: 13-20.
[14] 王姿曼, 郝瑞娟, 邓岳文, 等. 马氏珠母贝基因的克隆及表达[J]. 广东海洋大学学报, 2019, 39(1): 14-21.
[15] KIMURA K D,TISSENBAUM H A, LIU Y, et al. daf-2, an insulin receptor-like gene that regulates longevity and diapause in[J]. Science, 1997, 277(5328): 942-946.
[16] SONETTID,HEMEN W R A V , ROUBOS E W. Light- and electron-microscopic immunocytochemistry of a molluscan insulin-related peptide in the central nervous system of[J]. Cell and Tissue Research, 1992, 267(3): 473-481.
[17] GOMOTA,GOMOT L, MARCHAND C R, et al. Immunocytochemical localization of insulin-related peptide(s) in the central nervous system of the snail Helix aspersa Müller: Involvement in growth control[J]. Cellular and Molecular Neurobiology, 1992, 12(1): 21-32.
[18] 刘瑞, 白怀, 刘秉文. 胰岛素受体信号传递[J]. 生理科学进展, 2001(3): 63-65.
[19] 王蕊, 黄昆. Structure and Function of Insulin Receptor Family Proteins%胰岛素受体家族的结构与功能[J]. 中国生物化学与分子生物学报, 2009, 25(12): 1102-1109.
[20] 王涛, 赵晶, 杨安钢. 弗林蛋白酶:一种广泛参与前体蛋白切割的内切蛋白酶[J].医学分子生物学杂志,2006(3): 202-205.
[21] 张萍, 庞义, 杨波, 等. 弗林蛋白酶—一种重要的前体蛋白内切蛋白酶[J].生物技术通讯, 2002, 13(2): 63-65.
[22] DEVEV I E, POPOVA N V, PETRENKO A G. Determination of Alkali-Sensing parts of the insulin receptor-related receptor using the bioinformatic approach[J]. Acta Naturae, 2015, 7(2): 80–86.
[23] DEVEV I E, CHACHINA N A, SHAYAHMTOVA D M, et al. Mapping of alkali-sensing sites of the insulin receptor-related receptor. The role of L2 and fibronectin domains[J]. Biochimie, 2015, 111: 1-9.
[24] IGOR D, NATALIA C, EGOR Z, et al. Site-Directed Mutagenesis of the Fibronectin Domains in Insulin Receptor-Related Receptor[J]. International Journal of Molecular Sciences, 2017, 18(11): 2461.
[25] 王芳, 范瑞青. 四种滤食性贝类滤食器官鳃的扫描电镜观察[J]. 青岛海洋大学学报, 1998, 28(2): 240-244.
[26] GóMEZ-MENDIKUTI A, ELIZONDO M , VENIER P, et al. Characterization of mussel gill cells in vivo and in vitro[J]. Cell & Tissue Research, 2005, 321(1): 131-140.
[27] 封利颖. 虾夷扇贝IGFBP5、IGF2BP2基因克隆、表达分析及与生长相关的SNP位点筛查[D]. 青岛:中国海洋大学, 2013.
[28] BERTHELIN C, KELLNER K, MATHIEU M. Histological characterization and glucose incorporation into glycogen of the pacific oystertorage cells[J]. Marine Biotechnology, 2000, 2(2): 136-145.
[29] 马学敏. 双壳贝类消化生理的研究进展[J]. 生命科学仪器, 2010(1): 13-15.
[30] 詹秋羽, 郑普强, 王骥腾, 韩涛. 虾蟹类对糖类的利用和代谢[J]. 动物营养学报, 2020, 3(32): 97-110.
[31] 刘正琮, 上官步敏. 缢蛏性腺和消化腺的生化组成与组化反应[J]. 台湾海峡, 1993(3): 218-224.
[32] ZHENG Z, HAO R J, XIONG X W, et al. Developmental characteristics of pearl oyster: insight into key molecular events related to shell formation, settlement and metamorphosis[J]. Bmc Genomics, 2019, 20(1): 1-12.
[33] ADDADI L, JOESTERr D, NUDELMAN F, WEINER S. Mollusk shell formation: a source of new concepts for understanding biomineralization processes, Chemistry – A European Journal , 2006, 12(4): 980–987.
[34] 张立娟, 何毛贤. 生长激素与胰岛素样生长因子-Ⅰ对马氏珠母贝壳生长形成相关基因表达的影响[J]. 热带海洋学报, 2012, 13(2) :96-101.
[35] HAO R J, WANG Z M, YANG C Y, DENG Y W, ZHENG Z, WANG Q H, DU X D. Metabolomic responses of juvenile pearl oysterto different growth perfor-mances[J]. Aquaculture, 2018, 491: 258–265.
Gene Cloning and Expression Pattern Analysis of Insulin-related Peptide Receptor from
ZHAO Zi-han1, WANG Zi-man1, DENG Yue-wen1,2, YANG Chuang-ye1,2, LI Jun-hui1
(1.//2,524088,)
【】The insulin-related peptide receptor () gene ofwas cloned and its expression patterns in various tissues were analyzed.【】The full-length cDNA sequence ofwas cloned by rapid amplification of cDNA ends (RACE) and the tissue expression pattern of this gene was determined by RT-PCR. 【】The full-length ofwas 6 009 bp. It consist of a 5′UTR (615 bp), a 3′UTR (764 bp) and an open reading frame (4 627 bp) which encoded for 1542 amino acids. d Amino acids analysis of PmIRR showed that it contained a conservative domain Fu, three FNIII domains, a tyrkc domains, a signal peptide and two transmembrane domains.was differentially expressed in different tissues of, and the expression was significantly higher in gills, hepatopancreas and mantle edge.
; insulin-related peptide receptor; gene cloning; expression pattern
Q78; Q959.215+.4
A
1673-9159(2020)05-0034-09
10.3969/j.issn.1673-9159.2020.05.005
2020-04-09
农业产业技术体系专项(CARS-49);广东省教育厅珍珠研究创新团队(2017KCXTD016);广东省农业农村厅现代农业产业技术创新团队专项(2019KJ146)
赵子涵(1996-),女,硕士研究生,研究方向:珍珠培育与加工。E-mail:1922754931@qq.com
邓岳文(1975-),教授,主要从事珍珠贝遗传育种与养殖研究。E-mail:dengyw@gdou.edu.cn
赵子涵,王姿曼,邓岳文,等. 大珠母贝胰岛素相关肽受体基因的克隆与表达[J].广东海洋大学学报,2020,40(5):34-42.
(责任编辑:刘岭)
