印度尼西亚峇淡岛海域鱾新本尼登虫(Neobenedenia girellae)形态学及28S rRNA分子鉴定
2020-07-21乔莹马笑晚邵彦翔陈超
乔莹,马笑晚,邵彦翔,陈超
( 1. 自然资源部第四海洋研究所,广西 北海 536000;2. 中国水产科学研究院黄海水产研究所,山东 青岛 266071)
1 引言
本尼登虫作为世界海水养殖鱼类的一种主要的寄生性单殖吸虫,隶属于吸虫纲(Trematoda)单殖吸虫目(Monogenea)分室科(Capsalidae Bard, 1853)本尼登亚科(Benedeniinae Johnston, 1931),多分布于热带和亚热带海区。本尼登虫的宿主特异性较低,寄生宿主种类广泛,包括各类石斑鱼(Epinephelinae)、大黄鱼(Larimichthys crocea)、金目鲈(Lates calcarifer)、真鲷(Pagrus major)等多种野生及养殖品种。本尼登虫主要以虫体前后吸器及钩状结构吸附寄生于各类海洋鱼类的体表、鳍条、鳃丝等部位,破坏鱼类黏液组织,吸食宿主的表皮细胞及血液,营寄生生活。由于其寄生还可造成鱼体表面创伤,引发继发性的细菌感染,造成宿主死亡。至今关于本尼登虫病感染的报道屡见不鲜[1-4],有研究表明新本尼登虫已成为世界渔业及水产养殖中最主要的单殖类寄生虫[5-6],其宿主自然感染率高达70%,发病鱼死亡率平均在30%~40%,最高甚至可达90%以上,给养殖业者带来巨大的经济损失,严重影响着海水养殖产业的健康发展。
以往寄生虫分类方法是以传统的光学显微镜观察为主,依靠观察描述虫体及其器官的形态、大小及位置,并记录测量相关数据来进行形态鉴定。但传统分类学方法对鉴定者经验要求较高,且压片制片过程对于虫体及器官的位置和大小影响较大,故常存在同种异名的情况,引起分类归属的争议。新本尼登虫的分类争议同样存在,其争议点主要存在于新本尼登属的10个种是否存在同种异名的情况。Whttington和Horton[7]于1996年将已有报道的10种新本尼登虫重新进行分类,仅保留了 Neobenedenia melleni、N. adenea、N.isabellae、 N. longiprostata、 N. muelleri和 N. pacifica 6个种,N. girellae和N. pargueraensis被分为N. melleni的同种异名,但仍有学者对此分类持不同意见[8]。以核糖体基因(rRNA)和转录间隔区(First Internal Transcribed Spacer,ITS)等为分子标记[9-13]在分子分类学研究的应用,很好地解决了传统分类学方法的局限性,同时,鉴于该方法避免了样品处理对虫体形态的影响与人为形态观察的主观性,其已成为寄生虫分类的一种重要方法。
本研究针对印度尼西亚峇淡岛海域网箱养殖的卵形鲳鲹 Trachinotus ovatus (Linnaeus,1758)体表寄生的本尼登类单殖吸虫的形态学特征进行观察和测量,并结合其28S rRNA基因序列的扩增结果,对其进行分类鉴定,以确定危害印度尼西亚峇淡岛海域网箱养殖鱼类的本尼登类单殖吸虫种类,从而为该地区海水网箱养殖鱼类本尼登虫病的防治提供依据,为国家一带一路海水养殖业的发展提供技术支持。
2 材料与方法
2.1 标本采集
2017 年12 月于印度尼西亚峇淡岛海域(0°38′19″N,104°15′23″E)海水养殖网箱采集感染本尼登虫的卵形鲳鲹,寄生部位:体表、鳍条、鳃丝。经淡水浸泡后,收集脱落的寄生本尼登虫虫株(Isolate Batam, IB)。虫株分别使用10%中性甲醛和90%无水乙醇固定待用。
2.2 形态学测量
采集虫体使用盖玻片和载玻片轻轻压片后,使用10%中性甲醛进行固定。使用OLYMPUS光学显微镜对固定虫体形态学特征进行观察和测量。测量的形态学数据包括体长,体宽,前吸器长、宽,后吸器长、宽,咽长、宽,边缘膜,附甲片,前钩长,后钩长,卵巢长、宽,睾丸长、宽,共16组数据,测量单位均以μm计, 测量虫体数目为15~30 个, 并计算算术平均值和标准差。
2.3 28S rRNA扩增及测序
将使用适量90%无水乙醇固定的虫株样品,充分研磨后,12 000 r/min离心5 min,去除上清,使用无菌水重悬,充分洗涤样品,去除无水乙醇。虫体基因组DNA提取采用EasyPure®海洋动物基因组DNA试剂盒(Transgen Biotech,EE151-01),DNA 抽提与纯化步骤参照说明书进行。
28S rRNA扩增引物选用C1: 5′-ACCCGCTGAATTTAAGCAT-3′和 C3: 5′-CTCTTCAGAGTAC TTTTCAAC-3′[14]。扩增体系为:DNA模板 0.5 ng,PCR 预混体系 10 μL、Taq DNA 聚合酶 0.6 μL、上游引物C11μL、下游引物C31μL、加入无菌水至总体积50 μL。反应条件如下:95℃预变性5 min、95℃变性 30 s、55℃ 退火 45 s、72℃ 延伸2min、40个循环,最后72℃延伸7 min。PCR扩增产物经1% (w/v)琼脂糖凝胶电泳分离后,用EasyPure®凝胶回收试剂盒(Transgen Biotech, EG101-01)纯化。纯化产物由上海生工生物工程服务有限公司进行测序。
2.4 进化树构建
测序序列通过NCBI数据库进行比对,选取同源序列,使用MEGA X[15]软件进行多重序列比对,结果使用 ESPript软件(http://espript.ibcp.fr/ESPript/ESPript/)进行可视化展示。使用PhyML 3.0软件[16]进行进化树构建,自动模型选择使用赤池信息量准则(Akaike Information Criterion)模型,进化树改良类型选择SPR(Subtree Prune Regraft)。
3 结果
3.1 形态学描述
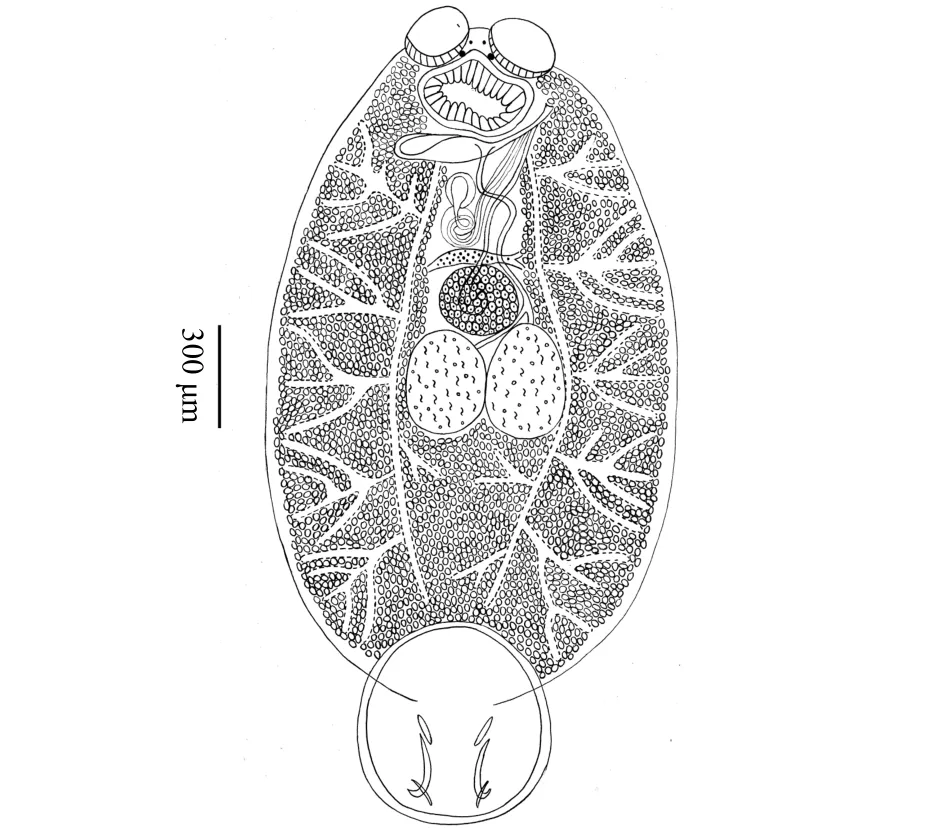
图1 鱾新本尼登虫峇淡岛虫株形态图Fig.1 Morphological structure of Neobenedenia girellae isolate Batam
形态描述:如图1所示,虫体呈现长椭圆形,体前部具有两个前吸器,体后部具有1个后吸器;体长(包括 后 吸 器 ) 为2976.54~4186.16 μm((3 275.33 ±516.12) μm),体宽1739.49~2 189.76 μm((1 922.41 ±189.27) μm);前吸器呈并列的吸盘状,大小为 [ 279.95~472.47 μm((364.37 ± 70.75) μm)]×[ 242.53~382.82 μm((323.61 ± 51.62) μm)];前吸器后具有两对眼点,前小后大,呈八字形排列,位于咽之前。咽呈圆形,边缘具有缺刻,大小为[ 274.80~553.95 μm((390.37 ± 109.24) μm)]×[ 426.57~685.19 μm((520.94±101.41) μm)];肠管于咽之后分叉,向后延伸至虫体末端形成盲端,并向虫体中央及边缘发出众多外侧分支。卵球形睾丸1对,并列排列于虫体中部,边缘具缺刻,大小为[ 330.57~440.95 μm((398.31 ± 45.77) μm)]×[ 280.06~392.71 μm((308.23 ± 47.57) μm)];卵巢位于虫体中部睾丸之前,无阴道结构,大小为 [ 235.43~292.62 μm((262.47 ±21.83) μm)]×[ 235.99~394.73 μm((296.28 ± 67.38) μm)];后吸器呈盘状,无隔膜结构,大小为[ 873.19~1 246.64 μm((1 049.25 ± 144.62) μm) ]×[ 818.44~1 237.99 μm((1 014.10 ± 164.83) μm)];后吸器上具边缘膜、附甲片、前钩1对、后钩1对以及边缘小钩数个。边缘膜大小为 35~54.74 μm((44.82 ± 7.28) μm);附甲片大小为 168.58~227.91 μm((201.12 ± 21.98) μm);前钩末端坚硬锋利,长 244.09~347.4 μm((301.05 ±46.78) μm); 后 钩 长 53.78~ 116.56 μm((90.49 ±30.53) μm)(表 1,图 2)。
3.2 28S rRNA序列分析及系统发育分析
测序获得IB虫株28S rRNA序列长度为394 bp(GenBank No. MN209942)。与 GenBank中同源序列的比对结果显示,IB虫株与其他本尼登虫属比对相似性在85.86%~99.47%之间。其中与鱾新本尼登虫N. girellae isolate N1: MH843708.1,isolate IDW109-08:MH843694.1,isolate IDW04-12:MH843693.1 等相似性最高,达99.47%;与珻氏新本尼登虫N. melleni:EU707805.1相似性为99.21%;与N. melleni isolate Zh-HGS20020517 相似性为98.91%;与Benedenia cf. seriolae FAS-2013 isolate SL93B5的相似性仅为85.86%。IB虫株28S rRNA序列与其他同源序列比对结果如图3所示。
基于28S rRNA序列构建系统进化树显示,IB虫株与其他鱾新本尼登虫和珻氏新本尼登虫聚为一簇,属于新本尼登虫属。卡氏嗜鼻虫(Nasicola klawei,HQ721184.1/ HQ721185.1/ HQ721186.1) 与 Capsala pricei(FJ971982.1)、Capsala laevis(FJ971979.1)聚为一簇,又与本尼登虫属种类聚类为一簇(图4)。
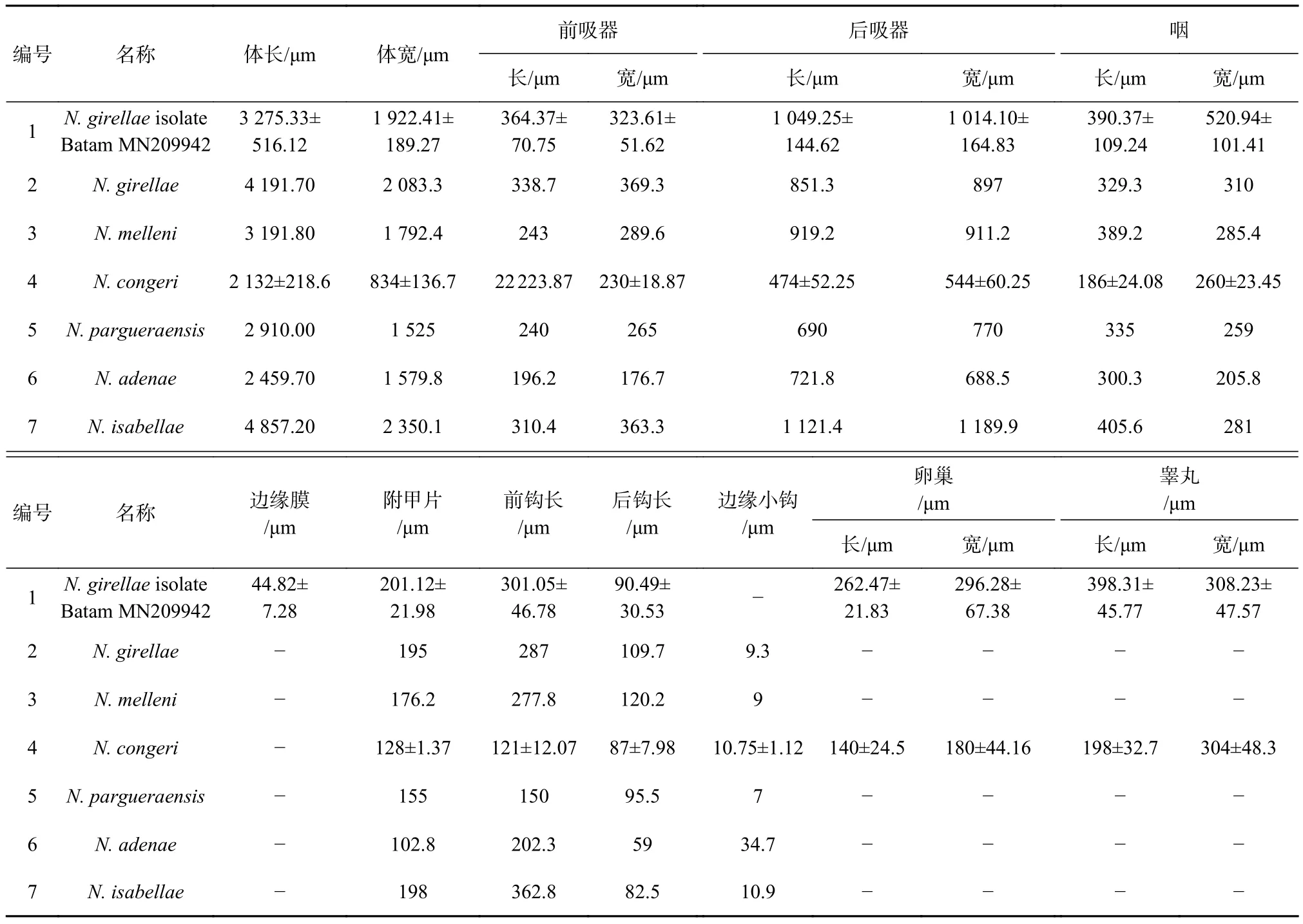
表1 N. girellae isolate Batam与其他新本尼登虫形态比较Table1 Comparison of morphological characters of N. girellae isolate Batam with other similar species
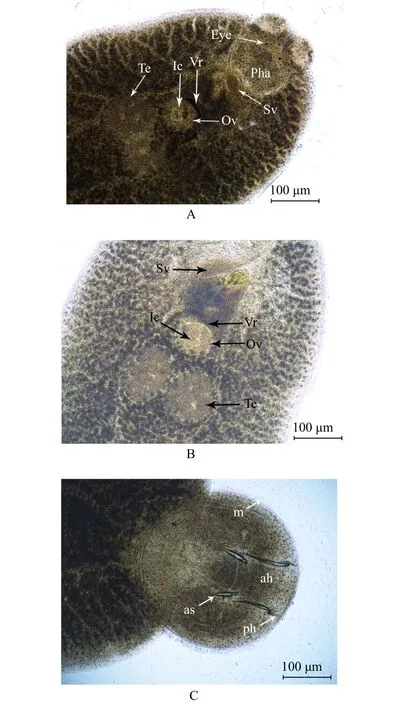
图2 鱾新本尼登虫光镜观察Fig.2 Morphology of Neobenedenia girellae isolate Batam
4 讨论
长期以来,形态学特征一直是系统发育及分类学研究的主要依据,而单殖吸虫类的分类多采用传统分类学方法,依赖肉眼和光学显微镜,通过对寄生虫外部形态、内部结构、寄生部位、流行性规律进行观察,从而进行分类鉴定。1963年Yamaguti[17]将单殖吸虫分室科分为5个亚科,其中本尼登亚科至今已通过研究鉴定到包括Allobenedenia、Neobenedenia、Pseudoentobdella等16属共50余种。本研究中,对印度尼西亚峇淡海域养殖卵形鲳鲹体表分离得到的IB虫株进行观察,结果显示其虫体后吸盘不具分隔,具窄边缘膜和3对形状不一的中央大钩。前列腺囊包裹于阴精囊内,无阴道结构,根据Yamaguti[17]及Whittington和Horton[7]的分类学标准,IB虫株具有新本尼登虫属的特征。
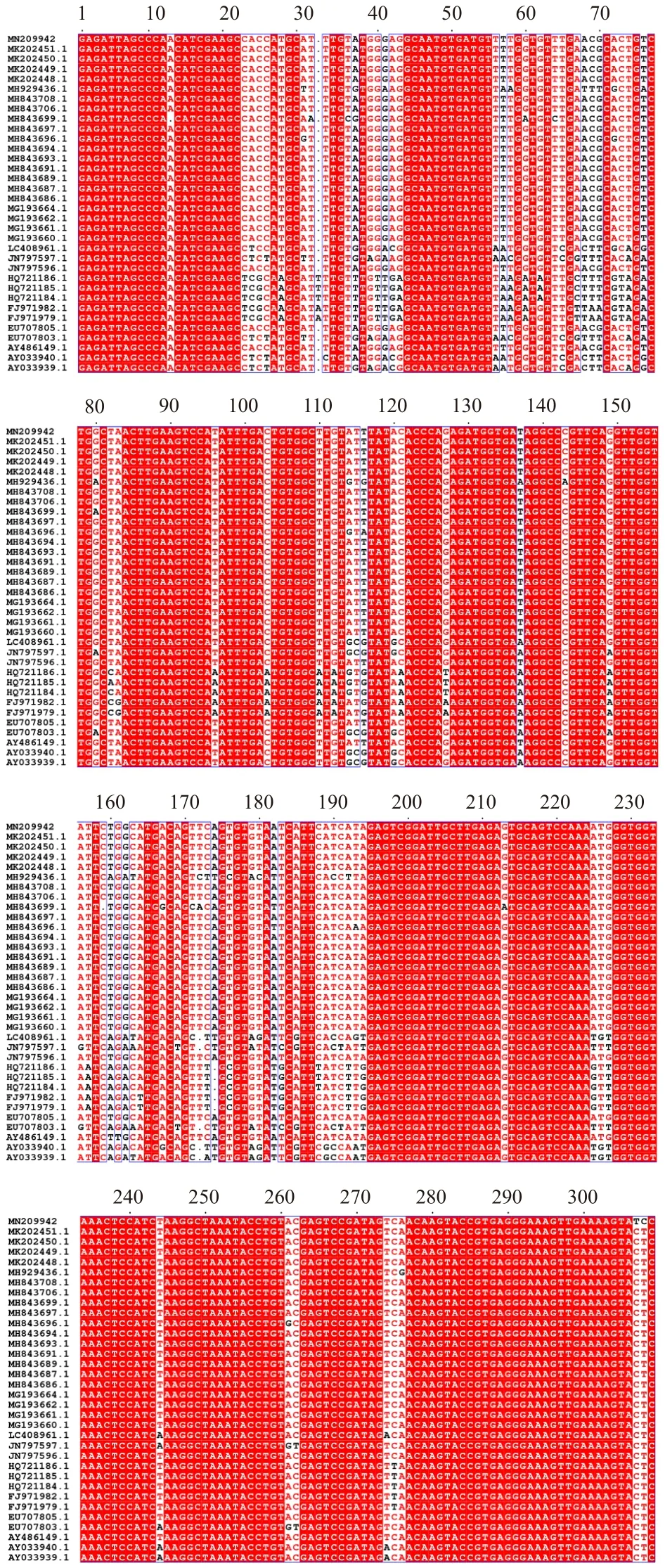
图3 鱾新本尼登虫28S rRNA序列同源比对Fig.3 Homology comparison of 28S rRNA of Neobenedenia girellae isolate Batam
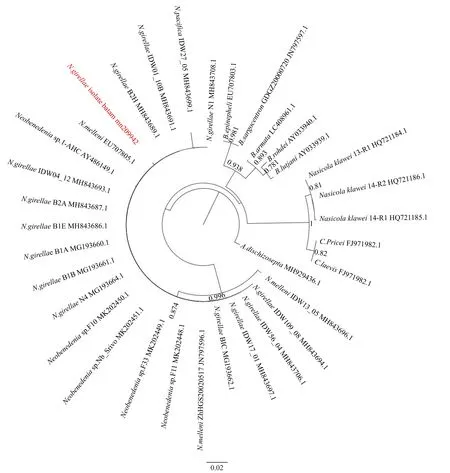
图4 鱾新本尼登虫系统进化树Fig.4 Phylogenetic tree analysis of Neobenedenia girellae isolate Batam
本研究中IB虫株的各项形态特性与杨文川记录的珻氏新本尼登虫形态较为相似,但形态学测量数据却存在一定的差异。已有研究表明寄生虫的宿主种类、寄生虫的年龄、寄生虫样本的处理方法均会对其虫体大小、形态学特征有一定的影响。此外Whittington和Horton[7]的研究同样证实了宿主的不同会导致相同种类虫体外部形态存在一定的差异这一观点。本研究在样品保存及处理过程中发现,使用酒精及福尔马林溶液固定的新本尼登虫,会出现萎缩、褶皱等不同程度的变形。以上通过形态学方法进行分类鉴定所暴露出的问题显示,仅仅依靠形态学的数据对新本尼登虫进行分类的结果将会存在诸多争议。
事实上自1963年Yamaguti[17]建立新本尼登属Neobenedenia开始,世界各地的新本尼登虫报道繁多,但由于该属的分类问题一直存在较大争议,故存在大量同种异名的情况。其中最为典型的争议就是N. girellae与N. melleni是否为同种异名品种。N. mel-leni最早在1927年由MacCallum首次发现并定名为Benedenia melleni。1963年Yamaguti建立新本尼登属后,将B. melleni更名为N. melleni,并作为该属的模式种。N. girellae由Hargis 1955年首次发现并定名为B.girellae,后由Yamaguti更名并移入新本尼登属。Hargis[18],Yamaguti[17],Bondad-Reantaso 等[19],Yoshinaga 等[8]研究观察到 N. melleni相较于N. girellae虫体更为宽短,N. girellae具有明显的滕腺,且两者后吸器钩大小及形状不同,认为珻氏新本尼登虫(N. melleni)和鱾新本尼登虫(N. girellae)分别为两个独立的种。而Whittington等[10]则认为两者仍存在如睾丸边缘缺刻,无阴道等共同的内部形态特征,故认为两者为同种异名的品种。
随着分子生物学技术的发展,采用分子生物学技术进行物种鉴定,遗传演化及物种亲缘关系的研究逐步增多。分子生物学技术与形态学相结合,在解决寄生虫系统分类以及某些疑难、易混淆种类的分类鉴定方面获得了很大成功[9,20-21]。Gunningham等最早采用分子生物学技术对鲑鱼3种三代虫的18s rRNA进行分析研究,显示不同虫种18S rRNA基因的V4区域存在明显差异,并以此设计出特异性探针来进行不同虫种的识别[22]。杨文川等[23]对鱾新本尼登虫生活史中发育各期幼虫的形态进行了详细的观察,描述和测量,发现其形态特征与Theo等描述的珻氏新本尼登虫各期幼虫特征非常相似,认为其属于同种异名。而张纹等[20]对于福建海区的鱾新本尼登虫与珻氏新本尼登虫28S rRNA序列进行比对分析,发现其遗传相似性高达99.7%,甚至远远高于测定的珻氏新本尼登虫种间相似度(99.41%),支持了两者为同种异名品种的论点。
本研究针对印度尼西亚峇淡岛海域分离得到的本尼登类寄生虫28S rRNA进行研究,并与GenBank中本尼登虫及新本尼登虫等亲缘物种的同源基因进行序列比对和系统进化研究。序列比对的结果显示本研究中的IB虫株与其他本尼登虫属的种类相似性在85.86%~99.47%之间,其中与鱾新本尼登虫类相似性最高,达99.47%;与珻氏新本尼登虫类相似性次之,为99.21%。而系统进化树结果显示,IB虫株与其他鱾新本尼登虫和珻新本尼登虫聚为一簇,属于新本尼登虫属。卡氏嗜鼻虫与Capsala pricei、C. laevis单为一簇,后又与本尼登虫属合为一簇,显示IB虫株与鱾新本尼登虫和珻氏新本尼登虫具有非常近的亲缘关系,该结果同样从分子生物学角度支持了Whittington等[10]的研究结果,即鱾新本尼登虫(N. girellae)和珻新本尼登虫(N. melleni)为同一种类的同种异名。
