哈欠传染的发展与演化:一项探索性研究 *
2020-07-20苏金龙苏彦捷
苏金龙 肖 瑶 苏彦捷
(1 南京师范大学心理学院,南京 210024) (2 北京大学心理与认知科学学院,行为与心理健康北京市重点实验室,北京 100871)
1 引言
哈欠是一种不由自主的、阵发性的呼吸活动,常常伴随着张大嘴巴、深吸气、短暂窒息、肌肉收缩达到短暂的高峰、闭上嘴巴同时伴有缓慢的呼气等一般过程,整个过程平均持续6秒左右(Provine, 1986, 2005)。哈欠可以自发产生(自发性哈欠),也可被诱发产生,如直接或间接观看他人打哈欠、阅读与哈欠有关的材料、听打哈欠的声音、甚至仅仅思考打哈欠这件事都能诱发哈欠(Arnott, Singhal, & Goodale, 2009; Norscia &Palagi, 2011)。这种经诱发而产生的哈欠被称为传染性哈欠(contagious yawning),其在人类和灵长类动物中广泛存在(Guggisberg, Mathis, Schnider, &Hess, 2010),并被认为具有重要的社会功能(Palagi, Leone, Mancini, & Ferrari, 2009)。在人类研究中,研究者多以大学生为被试,考察传染性哈欠与自发性哈欠的区别及相关机制(Platek,Mohamed, & Gallup, 2005);在灵长类研究中,研究者则多以黑猩猩为研究对象,尝试揭示传染性哈欠可能具有的演化功能(Anderson, Myowa-Yamakoshi, & Matsuzawa, 2004)。依照哈欠的社会功能假说,传染性哈欠是一种非言语交流形式,在维持和协调群体行为方面起着重要作用(Provine,1986)。该理论也得到了一些研究结果的支持。例如,研究发现,在雄性处于主宰地位的黑猩猩群体中,相比雌性发出的哈欠,个体更容易被雄性的哈欠传染(Massen, Vermunt, & Sterck, 2012);而在雌性主宰的倭黑猩猩群体中,群体成员则更容易被雌性的哈欠传染(Demuru & Palagi,2012)。这提示,群体中处于支配地位个体的哈欠更容易在群体中传染,而这类个体恰恰担负着协调群体行动的角色(Sauer & Sauer, 1967)。
在人类社会中,与社会交流存在密切联系的一项能力是观点采择,即理解他人并站在他人的角度进行思考的能力,它是人类交流得以实现的重要基础(Batson, Early, & Salvarani, 1997)。结合演化的有关观点,如果传染性哈欠是社会交流的表现形式之一,那么观点采择与其很可能存在某种联系,而近年来的研究也提示了这种可能性。例如,尽管自发性哈欠早在人类胎儿中就已被发现,但传染性哈欠直到四岁才出现,而恰恰是在这一年龄阶段附近,儿童才获得识别和理解他人心理状态的能力(Singer, 2006)。此外,在观点采择能力存在缺陷的精神分裂症患者和自闭症儿童中,研究者同样发现传染性哈欠功能缺损的迹象(Haker & Rössler, 2009; Senju et al., 2007)。另一方面,神经科学方面的证据则显示,被试在观看哈欠视频时,其打哈欠的冲动与腹内侧前额叶的激活存在相关(Nahab, Hattori, Saad, & Hallett,2009),腹内侧前额叶正是观点采择能力的重要神经基础(Frith & Frith, 2006)。然而,尚未有研究从行为层面直接考察传染性哈欠与观点采择之间的关系。
此外,从演化的时间轴来看,相比自发性哈欠,传染性哈欠的出现可能相对更晚。相比于自发性哈欠单纯的生理功能(如调节大脑唤醒水平),传染性哈欠被赋予了新的社会交流功能(Gallup, 2011)。自发性哈欠在很多物种中普遍存在,而传染性哈欠,虽然在部分低等哺乳类动物当中也发现了该现象,但其还是更多地存在于人类和一些高级灵长类动物当中(Gallup, 2011;Guggisberg et al., 2010)。另一方面,传染性哈欠和无意识模仿(mimicry)、行为同步化(synchrony)等现象具有高相似性,这种相似性不仅体现为外在特征的类似,也体现在其共享的神经网络(镜像神经元)方面(Haker, Kawohl, Herwig, & Rössler,2013),而上述现象与群际关系维持都存在密切联系(Feldman, 2006; Lakin, Jefferis, Cheng, &Chartrand, 2003)。这一系列的证据提示,从演化的角度来说,传染性哈欠应该产生于自发性哈欠的基础之上,并在功能上与相对更加高级的社会功能产生联系。如果这一假设成立,则可以预测,相比自发性哈欠,传染性哈欠与同样在演化上出现相对较晚的一些高级认知功能的相关应该更高,如抑制控制。因为该能力与无意识模仿样行为(如无意识模仿、行为同步化、情绪感染)在功能上密切相关,并隶属于后者的直系上行调控系统(Logan, Yamaguchi, Schall, & Palmeri, 2015;Spengler, von Cramon, & Brass, 2010)。同时,Sigurdsson等人(2017)也发现,大脑的抑制性与传染性哈欠存在直接相关,而他人在场的影响也提示了抑制机能和传染性哈欠之间的某种联系(Gallup, Church, Miller, Risko, & Kingstone,2016);相应地,与哈欠相关的一些生理因素,如警觉程度和睡眠状况(Giganti & Zilli, 2011),也应该更多地与自发性哈欠相关(与传染性哈欠相比)。如果这些能够被证实,则可以从另一个层面为传染性哈欠的社会功能假说提供某些支持。
综上所述,本研究着眼于在人类群体中初步验证传染性哈欠的社会功能假说。一方面,通过考察观点采择与传染性哈欠的关系来对该假说进行相对直接的验证;另一方面,通过考察抑制控制、同哈欠有关的睡眠状况和警觉程度,分别与自发性哈欠和传染性哈欠相关程度的差异来进行间接佐证。在研究群体的选择上,本研究采用大学生和老年人两个样本。纳入老年人样本是基于已有研究多围绕儿童和大学生展开,老年人群体的数据较少(Massen & Gallup, 2017)。此外,通过不同年龄群体的对比,也有助于深入了解传染性哈欠背后可能存在的深层规律。总的来看,作为一项探索性研究,结合已有实证研究和相关理论,提出以下假设。
假设1:观点采择与传染性哈欠存在正相关关系。假设2:相比自发性哈欠,传染性哈欠与抑制控制之间的相关应该更高。假设3:相比传染性哈欠,近期睡眠状况和警觉程度与自发性哈欠之间的相关应该更高。
2 方法
2.1 被试
大学生样本为通过网络公开招募的北京某高校在读学生57名,年龄M=22.55岁,SD=2.97岁,其中男性20名;老年人样本则采用方便取样的方式,招募了山西某地企业退休职工52名,年龄M=71.76岁,SD=6.09岁,其中男性44名。所有被试的视力或矫正视力正常,无发展障碍、精神疾病或神经系统疾病。
2.2 实验材料
视频材料:哈欠视频和控制视频。其中哈欠视频记录了个体打哈欠的过程,对应的控制视频则由同一个体保持中性表情,不做出任何行为(参见图1)。视频由研究者自行制作。在具体制作过程上,首先收集了49段人物哈欠视频,之后由12 名大学生(6名男生)被试对这些哈欠视频中主角表现出的哈欠进行真实性评分(Likert 5点计分),选出平均评分高于3.5的哈欠视频作为正式实验材料。最终,进入本实验的哈欠视频包含10个(视频中哈欠主体包含婴儿、大学生和老年人,性别平衡),平均时长8.2s。在正式实验中,每段视频呈现2次,不同视频呈现顺序随机(对应的10段控制视频在上述内容参数和呈现参数上进行匹配)。
抑制控制测量。采用Simon任务(参见图2),该任务由Simon和合作者开发,其基本原理为:刺激与反应特征的匹配程度会影响被试的反应,具体表现为二者越匹配,被试的反应速度越快,准确率越高(Simon & Wolf, 1963)。本研究中,屏幕左侧或右侧会随机呈现“左”或“右”字,被试需要对汉字进行反应,“左”字按“v”,“右”字按“n”,其中汉字出现的频次及位置均进行匹配。任务包括16个练习试次和40个正式试次,以在正式试次中一致条件下(“左”字呈现在屏幕左侧或“右”字呈现在屏幕右侧)的反应时减去不一致条件下(“左”字呈现在屏幕右侧或“右”字呈现在屏幕左侧)的反应时作为被试的抑制控制得分。得分越高,抑制控制能力越强。

图1 哈欠组(左)和控制组(右)视频材料示例
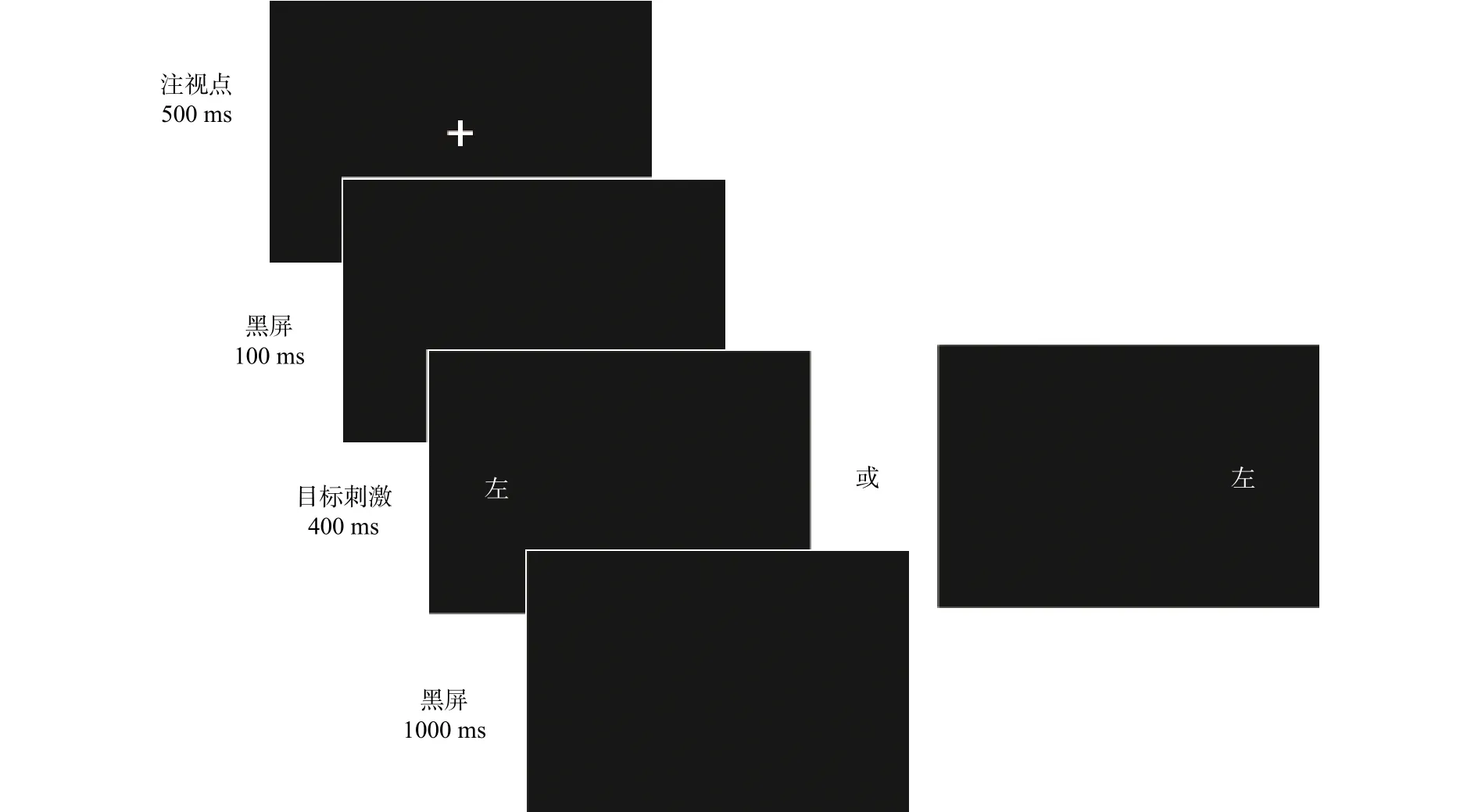
图2 Simon任务流程图
观点采择测量。采用人际关系指针量表(Interpersonal Relationship Index, IRI)中的观点采择维度,共包含5个条目(Likert5点计分),如“在我决定做某件事情之前,我会试着去听听其他人的不同意见”,被试的得分为5道题目得分的总和(张凤凤, 董毅, 汪凯, 詹志禹, 谢伦芳,2010)。在本研究中,问卷的克隆巴赫α系数分别为0.774(大学生被试)和0.828(老年人被试)。
睡眠状况及警觉程度测量。分别采用匹兹堡睡眠质量指数(PSQI)(Buysse, Reynolds III,Monk, Berman, & Kupfer, 1989)和斯坦福嗜睡量表(SSS)(Hoddes, Zarcone, Smythe, Phillips, &Dement, 1973)。前者主要用来评定最近1个月的睡眠质量,其中参与计分的18个条目测量了入睡时间、睡眠时长、效率、睡眠障碍、自评睡眠质量、催眠药物应用和日间功能7个方面,每个方面按照0~3计分,所有方面得分总和即为睡眠质量指数得分,得分越高,被试的睡眠质量越差。后者主要用来测量被试当下的警觉程度,需要被试对自己当前的警觉程度进行自我评估,从“思维非常活跃,意识非常清醒”(计1分)到“无力抵抗困意,很快开始入睡,有梦一般的朦胧意识”(计7分)一共七个层级。得分越高,警觉程度越差。
2.3 实验程序
进入实验室后,被试会被带入一个内置有隐蔽摄像头的房间观看哈欠视频和对应的控制视频,并完成Simon任务和矩阵游戏。指导语为:“你需要认真观看视频,将注意保持在视频中人物的面孔上,想象视频中的人正坐在你对面。”被试独自在房间中按操作说明播放视频,观看哈欠视频和控制视频的先后顺序在被试间平衡。其间,在每组视频呈现后有1.5 min空白视频(一般认为,传染性哈欠可能会有一定延迟,这种延迟一般在1.5 min之内)(Helt, Eigsti, Snyder, & Fein,2010; Provine, 2005),此时被试可以在椅子上休息,但不能离开座位,不能使用手机。隐蔽摄像头始终正对被试面孔,以便记录哈欠反应。
之后,被试需要完成测量观点采择、睡眠状况和警觉程度的问卷,并填写年龄、性别等一般人口统计学信息。最后,主试向被试解释本实验的实验目的,并支付一定数量的报酬。
2.4 数据处理
由一名不知道实验目的与视频内容的大学生志愿者对隐蔽摄像头录制的视频进行编码,记录被试在观看视频过程中的哈欠频次,编码的标准包括张嘴、面部肌肉收缩、深呼吸等打哈欠时的一般性身体反应,只有在三个标准同时满足的情况下,被试的表现才被认为是哈欠。此外,随机抽取其中20%的数据,由另一名不知道实验目的及视频内容的大学生志愿者编码,编码一致性为100%。
针对被试在Simon任务中的反应时数据,删除了不一致条件下平均反应时低于一致条件下平均反应时的被试的数据。所有数据的统计处理都采用SPSS20.0完成。
3 结果
大学生被试。为了从总体上考察在被试群体中是否存在传染性哈欠,本研究把被试在观看视频过程中的表现进行2点计分(是/否打哈欠),发现相比观看控制视频(55%的被试打哈欠),被试在观看哈欠视频(76%的被试打哈欠)过程中打哈欠的可能性显著提升(Mcnemar检验,χ2=7.04,p<0.05)。之后,用被试在观看哈欠视频时打哈欠的次数减去其在观看控制视频时打哈欠的次数,把结果大于0的被试限定为表现出传染性哈欠的个体,发现有64%的个体表现出传染性哈欠,具体传染性哈欠个数的频次分布参见图3。对比表现出传染性哈欠的大学生与没有表现出传染性哈欠的大学生在观点采择、抑制控制、PSQI、SSS上的得分,均无显著差异,|t|s≤1.32,ps≥0.19。

图3 传染性哈欠个数的频次分布
以表现出的传染性哈欠的数量作为哈欠易感程度指标,统计分析显示,哈欠易感程度与年龄(Spearman'sr=-0.13,p=0.50)和性别[Spearman'sr=-0.16,p=0.42(男=1, 女=0)]都不存在显著的相关。观点采择与哈欠易感程度也不存在显著相关,Spearman'sr=-0.08,p=0.70。关于抑制控制、睡眠状况和警觉程度分别与传染性哈欠和自发性哈欠之间的关系,参见表1。可以看出,抑制控制显示出更强的与传染性哈欠相关的趋势(-0.37、-0.08),而两项与哈欠有关的生理因素则都显示出更强的与自发性哈欠相关的趋势(0.25、-0.08;0.21、-0.14),虽然只有抑制控制与传染性哈欠的相关系数在统计上达到了边缘显著,p=0.09。

表1 变量相关系数列表(Spearman's rs)
老年被试。统计结果显示,观看控制视频时3个(5.7%)被试打了哈欠,观看哈欠视频时6个(11.3%)被试打了哈欠,不存在显著差异,χ2=0.57,p=0.45(McNemar检验),提示老年人群体中没有表现出哈欠传染的现象。如果从个体层面看,仍然用被试在观看哈欠视频时打哈欠的频次减去其在观看控制视频时打哈欠的频次,把结果大于0的个体限定为被哈欠传染的个体(n=5),进一步发现被哈欠传染的个体和没有被哈欠传染的个体在观点采择、抑制控制、PSQI、SSS得分上均无显著差异,|Z|s≤1.62,ps>0.10(Mann-Whitney U检验)。
最后,为了更全面地考察传染性哈欠的可能影响因素,对大学生的数据和老年人的数据进行联合分析。以传染性哈欠的数量为因变量,以年龄、性别、PSQI、SSS、观点采择和抑制控制为预测变量建立回归模型。依据数据特点,选择广义线性模型里的泊松回归,以log函数为联接函数,采用最大似然法对数据进行参数估计,结果见表2。可以看出,所有预测变量皆不显著。考虑到最大似然估计对样本量的依赖,也用迭代重加权最小二乘法(iterative reweighted least squares)进行了参数估计,有关结果在显著性上并没有发生变化。
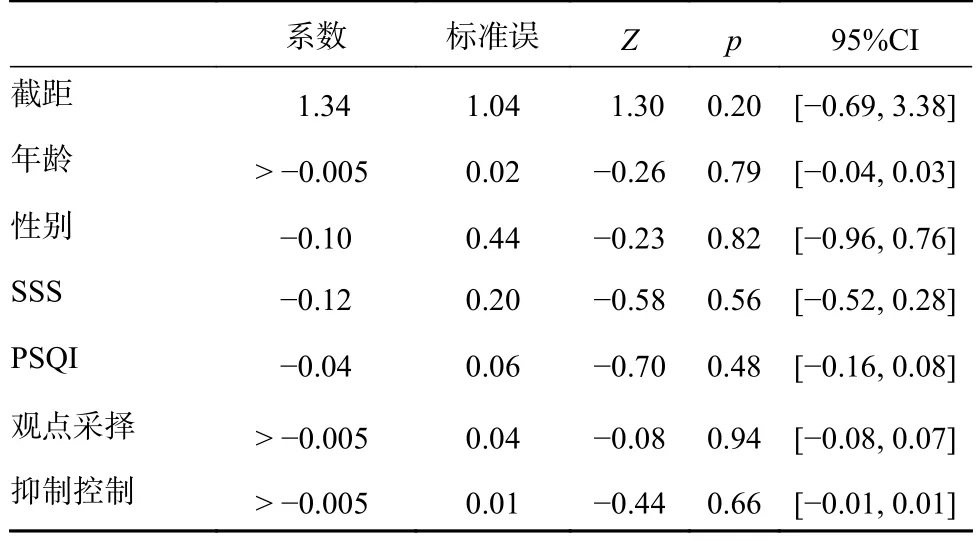
表2 有关变量预测传染性哈欠数量的广义线性模型拟合结果
此外,还对比了大学生和老年人在观点采择、SSS、PSQI上的得分,发现均无显著差异,ts<1;仅在抑制控制能力上,年轻人要明显优于老年人,t=-5.32,p<0.001。
4 讨论
作为一种生理现象,哈欠是疲劳状态下的一般反应,在调节血氧水平和大脑唤醒程度方面起着重要作用(Ramirez, Ryan, Eldakar, & Gallup,2019)。但如同在演化过程中一些表型(phenotype)(如,歌唱)总会被以某种形式重新利用而被赋予新的功能(Pearce, Launay, MacCarron, & Dunbar,2017),哈欠也可能具有类似的演化历程。
首先,大学生群体中存在着传染性哈欠,这与已有研究结果相一致(Platek et al., 2005)。但在老年群体中,很少个体表现出传染性哈欠。已有研究显示,传染性哈欠在个体4岁左右开始出现(Helt et al., 2010),而本研究结果提示,步入老年期后,传染性哈欠可能会逐渐消失。为什么会这样呢?本研究结果部分对比了大学生和老年人在观点采择、抑制控制、SSS、PSQI上的得分,发现只有在抑制控制能力上两个群体存在显著差异,结合大学生群体中发现的抑制控制与哈欠易感程度的正相关边缘显著,提示抑制控制能力与传染性哈欠可能存在着共变关系。此外,抑制控制能力在4岁时显著提升(徐芬, 王卫星, 高山,Sabbagh, 2003),也提示了这种共变关系的可能性。而其原因有两种可能性:第一,抑制控制和传染性哈欠两个变量中,某个变量为另一变量的基础;第二,两个变量共同以第三个变量为基础。基于相关研究,本研究认为后一种可能性更大,即二者属于在人类演化过程中服务于某同一功能而演化出的心理能力。
其次,在大学生群体中发现,相比自发性哈欠,抑制控制呈现出与传染性哈欠相关更强的趋势;而相比传染性哈欠,睡眠状况和警觉程度则呈现出与自发性哈欠相关更强的趋势。某种程度上支持了假设2和假设3。这也提示传染性哈欠和自发性哈欠在演化上可能具有一定的时序性,之前提及的哈欠在物种中普遍存在,而哈欠的传染仅在某些物种中存在,以及“哈欠在新生儿中已经出现、但传染性哈欠却需要到4岁左右才会出现”也佐证了这种可能性(Gallup, 2011; Helt et al.,2010)。然而,需要注意的是,这些证据还很粗浅,也不够直接。未来的研究可借助神经科学的方法,结合不同脑区演化早晚的已有证据,探索不同哈欠类型的脑机制以进一步明确二者的区别,将对理解该问题意义深远。
此外,研究结果并没有支持假设1。本研究并未发现传染性哈欠与观点采择存在相关。目前并没有研究专门探讨两者的关系。进一步检索文献发现,有3篇文献在探讨传染性哈欠与其他变量的关系时附带报告了这两者的相关。但遗憾的是,上述3篇文献均未发现二者存在显著相关(Bartholomew &Cirulli, 2014; Gottfried, Lacinová, & Širůček, 2015;Haker & Rössler, 2009)。研究者考察了被试在共情商数问卷(EQ问卷)(Baron-Cohen & Wheelwright,2004)中的得分以及一阶错误信念任务中的得分与传染性哈欠的关系,发现均存在显著正相关(Arnott et al., 2009; Platek, Critton, Myers, & Gallup,2003),而这两个测量与观点采择能力存在着密切联系,尤其被试在EQ问卷上的得分,其反映的共情能力与观点采择存在很大的重合。鉴于此,本研究对观点采择能力与传染性哈欠的关系持相对开放的态度。此外,Arnott等人的研究测量的是听觉通道的哈欠传染,即听到打哈欠的声音时被试是否会表现出更多的哈欠。而本研究是通过视觉通道呈现哈欠表情,这也提示不同的传染通道可能会影响相关效应。在今后的研究中,考察不同通道呈现哈欠刺激对传染性哈欠的影响将对进一步研究传染性哈欠有一定的助益。同时,本研究中观点采择的测量是采用自我报告的方式,一方面,基于自我报告方法本身的局限,其数据在反应对应测量构念上可能会存在一定偏差;另一方面,Likert5点计分方式由于选项本身数量受限,导致被试间存在的真实变异程度可能被低估,这就需要更加灵敏的测量(如反应时)。这意味着,对观点采择测量的进一步精准化也将对相关问题的解答提供必要支撑。
那么,传染性哈欠是否具有社会功能呢?虽然本研究无法直接给出答案,但基于研究结果,至少可以推测,相比自发性哈欠,传染性哈欠更可能与一些高级的心理机能相联系。传染性哈欠与认知控制能力的关联提示,其与演化上出现相对更晚的额叶存在联系,而与更低级的生理机能(如睡眠状况和警觉程度)在关联程度上的弱化,也提示了其不像自发性哈欠那样更多地服务于生理调节。关于额叶的演化,存在着不同的视角,Dunbar(1992)认为其与人类群体规模存在密切联系,规模越大越需要复杂的群体协作模式,所以演化出了新皮层;脑演化的文化视角则认为,额叶尤其是前额叶的演化与人类的复杂文化存在密切联系(Muthukrishna & Henrich, 2016),而文化的基础是社会学习(Tennie, Call, &Tomasello, 2009);智力假说则认为额叶的演化与人类越来越高的智力直接相关,相比其他物种,人类更为发达的智力体现在心理理论、模仿和语言等方面(Roth & Dicke, 2005),而上述三个成分无一不和人类的社会性密切关联。可以看出,虽然不同的观点在解释额叶的演化上存在差异,但存在一个共识,即额叶的演化与人类的社会机能存在联系。这也间接提示了传染性哈欠可能具有一定的社会功能。另一方面,和一些更加高级的心理成分如心理理论(ToM)比起来,传染性哈欠可能具有的社会功能显然要弱很多,这一点也可以从本研究获得的其与观点采择之间的阴性结果中得以窥见。实际上,结合引言部分提及的镜像神经元及外在表现结构特征,传染性哈欠在社会功能上可以更多地与情绪感染类比,而具体功能及其机制的揭示则有赖于未来更多的实证研究。
研究表明在大学生群体中表现出哈欠传染的现象,并且哈欠传染程度具有年龄差异。抑制控制与哈欠传染具有共变关系,为传染性哈欠的社会功能假说提供了初步的实证支持。而关于传染性哈欠与观点采择等变量的关系,则更多地为未来研究提供了某些思考。
5 结论
传染性哈欠承载着潜在的社会功能,且该现象具有年龄差异。这一现象在大学生群体中表现明显,但步入老年期之后则逐渐消失,抑制控制能力可能在这种年龄变化中起到了某种作用。
