不同林龄油松根际土壤微生物群落多样性及其影响因子
2020-07-08冀玉良
丁 丽, 冀玉良, 李 懿
(1.包头铁道职业技术学院 建筑工程系, 内蒙古 包头014060; 2.中国科学院 成都生物研究所, 成都 610041)
作为地球化学循环过程中的重要参与者,土壤微生物参与土壤中有机质分解、元素循环和土壤矿化等化学活动,能够促进土壤物质循环、稳定与保持土壤的生产力和健康[1-2]。土壤生态系统如果想使功能、完整性和可持续性得到维持,必须要保持土壤微生物的多样性。多年的研究发现,微生物的多样性是增强微生物功能和生态系统稳定性的重要因素[3-5]。作为直接受植物根系和分泌物影响的土壤区域,根际土壤为土壤微生物与植物相互作用提供了场所[4],使微生物与根系之间保持了适应性协同进化和互惠的关系[6-8]。在植物根系活动的影响下,根际土壤的养分含量及其他土壤理化性质发生了变化,进一步使根际微生物群落的组成发生改变,非根际与根际土微生物群落组成和多样性也出现了很大的差异。植物生长需要根际微生物的参与和作用,土壤营养物质能够被促植物生长细菌(PGPR)的代谢活动进行分解、转化,成为植物可吸收利用的形态后被吸收,有效地促进了植物的生长,而一些细菌分泌的激素能够提高植物的适应性[9-10]。植物的种类、健康状况、生长阶段等因素会影响根际土壤微生物的群落结构。另外,如土壤营养状况、pH值、温度和水分等土壤理化性质会对微生物群落产生较大影响[11]。
植被生长除了需要一定的水分和养分之外,还需要合适的、有酶活性及微生物活动的土壤环境,能够促进土壤活性及肥力。很多学者从土壤及微生物的关系角度,探究植被生长发育过程,发现微生物活动及其肥力的保持等规律,明确植被、土壤及微生物之间的关系[12-13],为实践提供理论指导。但是,目前的研究中,研究岷江流域的根际土壤较少。因为该地区的土质较差,贫瘠没有肥力,影响了植被的生长,造成水土流失较为严重,为保持该区域的水土[14-15],一般通过人工育林的方式进行巩固,就对树种的要求较高,否则人工林的生长发育会受到影响。油松(Pinustabulaefrmis)是一种松科常绿乔木,大多分布在我国的亚热带区域,具有良好的生长特性和环境适应性,作为人工育林树种入选,表现优异,能够有效地调节森林生态平衡,促进退耕还林工作的发展。油松的生长发育期较长,中间会出现林地肥力下降等问题[16-17]。这时,根际是油松与土壤进行能量交换与吸收的转换器,生物活性较强,发挥了巨大的调节作用,通过分泌不同的有机质,平衡土壤中的营养元素,改善土壤结构,根际土壤特性变化比较明显[18-19]。在此基础上,本研究采用Illumina-MiSeq高通量测序技术,对不同林龄油松非根际与根际土壤中真菌18SrDNA基因V4区片段和细菌的16SrDNA基因V3—V4区片段做测序,发现和挖掘不同林龄油松非根际与根际土壤微生物群落多样性及其与土壤环境因子之间的关系,为做好以油松为代表的人工育林工作提供参考和借鉴。
1 材料与方法
1.1 研究区概况
本试验选定区域地处岷山南麓、长江流域最大的支流——岷江流域,具有非常复杂的地形地貌,拥有的干流超过735 km,并因地势高低形成3 km左右的落差,上中游分界处为乐山、都江堰,上游区域的峡谷很多,地形复杂多变,并未形成大量成片的耕地,居住的人口非常少;中下游区域地势比较平缓,大多是丘陵平原,利于人类居住,分布有大量的生活区和农业种植区。受经纬度影响,该区域为亚寒热带气候,上游区域气温低于中下游气温,据多年气象资料记载,该区域内夏季比较炎热,最高温接近39℃,冬季气温较低,最低温为-4℃左右,出现在1月份,所以年均气温为9℃。该区域的无霜期接近半年,全年日照时长为2 100 h,雨季分明,土壤为紫色土。
1.2 试验方法
2018年8月选取岷江上游的不同林龄的油松林(幼龄、中龄、成熟龄、过熟龄),每种类型油松林设置3个长、宽均为50 m的样地,定期在每个样地标记3株油松采集根际和非根际土壤样品,其中根际土采用抖落法,即抖落大块土壤,以根围5 mm内的土壤作为根际土,根围外5 mm土为非根际土壤,总共采集土壤样品24份(根际土12份,非根际土12份);其中每一份根际土和非根际土分为3小份,一份用液氮保存,带回实验室后-80℃保存测定微生物群落结构;另外一部分车载冰箱4℃保存测定土壤微生物量碳氮和酶活性;第三部分常温下风干处理用以测定土壤养分。
1.3 土壤养分和酶活性
采用滴定法、平板梯度稀释法和比色法分别测定土壤脱氢酶、过氧化氢酶及脲酶,对有机碳、pH值、有效磷和有效氮等土壤养分[20],借助于《土壤农化分析》进行测定。
1.4 土壤总DNA提取
利用Fast DNA spin kit for soil 试剂盒(MP BIO,Inc.,Irvine,CA,USA)进行土壤DNA的提取,然后采用1%的琼脂糖凝胶电泳,NanoDrop ND-1000微光分光光度计测定浓度。
1.5 土壤微生物基因组DNA的提取
往2 ml无菌离心管中滴入1 ml根际土壤溶液,于4℃,10 000 g离心30 s,弃上清,作为根际土壤微生物基因组DNA提取之用;往2 ml无菌离心管中滴入大约0.2 g非根际土壤(湿重),用无菌药匙量取,作为提取基因组DNA之用。提取基因组DNA时使用OMEGA试剂盒E.Z.N.ATM Mag-Bind Soil DNA Kit试剂盒(OMEGA),检测DNA的长度和完整性时使用1%琼脂糖凝胶,检测DNA的浓度和纯度时使用NanoDrop。
1.6 DNA扩增及测序
对细菌16S rDNA V3—V4区进行扩增时,使用两轮法。第一轮扩增,上游引物341F:CCCTACACGACG-ACGATCTTACAGATAGGGGGCGCAG,下游引物805R:GACTGGAGTTCCTTGGCACCCGAGAATTCCAG-ATACGGGTATCTAATCC,利用融合了barcode序列的通用引物进行;真菌18S rDNA扩增,第一轮扩增,上下游引物序列为NS1:CCTACACGACGATCCTATGCTTAACGTG,Fungi:GAAGTCCCTTATACGGGGATTTACCG-TAAACGT,利用融合了barcode 序列的通用引物进行。反应体系30 μl,包含各1 μl引物(10 μmol/L),15 μl 2×Taq master Mix(Thermo),模板DNA 20 ng。扩增条件为:94℃ 30 s,94℃ 3 min,65℃ 30 s,45℃ 20 s,扩增5个循环;94℃ 20 s,72℃ 30 s,55℃ 20 s,扩增20个循环,72℃延伸5 min。第二轮扩增,以第一轮的PCR产物为模板,使用Illumina 桥式PCR兼容引物。反应体系同上。扩增条件:95℃ 15 s,95℃ 30 s,72℃ 30 s,55℃ 15 s,扩增5个循环,72℃延伸5 min。PCR产物经磁珠法回收并定量。等量混合每个重复个体的PCR产物,使每份混合样品中都包含全部重复个体,将混合样品送至生工(上海)生物工程有限公司,使用Illumina-MiSeq平台进行高通量测序。
1.7 数据分析
去除引物接头序列和各低质量碱基(pH值red Quality Score=20)后,对测序原始数据进行拼接,去除长度短于200 bp的序列,非特异性扩增序列及嵌合体,就会得到每个样本的有效序列数据划定阈值为97%,针对18S和16S序列进行操作分类单元(operational taxonomic unit,OTU)的划分。进行主坐标分析(PCoA)时,以样本OTUs组成之间的weighted unifrac距离矩阵为依据,寻找微生物群落之间的相似性;对土壤养分和微生物群落之间进行冗余分析(Reundancy analysis,RDA)时,采用Canoco 4.5软件。
用SPSS 18.0和Excel 2007进行方差检验和数据统计,为检验各指标之间的相关性,可以通过单因素方差分析(One-way ANOVA),平均值±标准误差表示(Mean±SE)和Pearson相关系数法实现。
2 结果与分析
2.1 不同林龄油松根际与非根际土壤养分
从表1中看出,不同林龄油松样地,土壤中根际土壤pH值低于非根际土壤,说明由于根在土壤中长期生长,过程中不断向根际分泌出有机化合物,油松林的土壤pH值处于弱酸性,属较强酸性土壤,随着林龄的增大,pH值也逐渐增加。土壤中根际和非根际有机碳含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有机碳含量显著高于非根际(p<0.05),过熟林根际有机碳含量与非根际差异并不显著(p>0.05);根际土壤有机碳含量在幼林、中林、成熟林和过熟林分别高于非根际15.98%,17.63%,10.81%,15.31%。土壤中根际和非根际全氮含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际全氮含量显著高于非根际(p<0.05),过熟林根际全氮含量与非根际差异并不显著(p>0.05);根际全氮含量在幼林、中林、成熟林和过熟林分别高于非根际13.95%,12.39%,11.57%,3.33%。土壤中根际和非根际有效磷含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有效磷含量显著高于非根际(p<0.05),过熟林根际有效磷含量与非根际差异并不显著(p>0.05);根际有效磷含量在幼林、中林、成熟林和过熟林分别高于非根际16.98%,10.90%,24.28%,1.81%。土壤中根际和非根际有效钾含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有效钾含量显著高于非根际(p<0.05),过熟林根际有效钾含量与非根际差异并不显著(p>0.05);根际有效钾含量在幼林、中林、成熟林和过熟林分别高于非根际20.23%,12.59%,10.14%,6.37%。

表1 不同林龄油松根际与非根际土壤养分
注:*表示根际与非根际差异显著(p<0.05),下表同。
2.2 不同林龄油松根际与非根际土壤酶活性
由表2可知,不同林龄油松表现出各种酶类的活性,其根际土壤高于非根际土壤。土壤中根际和非根际蔗糖酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际蔗糖酶活性显著高于非根际(p<0.05),过熟林根际蔗糖酶活性与非根际差异并不显著(p>0.05);根际蔗糖酶活性在幼林、中林、成熟林和过熟林分别高于非根际28.00%,33.73%,27.66%,12.48%。土壤中根际和非根际碱性磷酸酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际碱性磷酸酶活性显著高于非根际(p<0.05),过熟林根际碱性磷酸酶活性与非根际差异并不显著(p>0.05);根际碱性磷酸酶活性在幼林、中林、成熟林和过熟林分别高于非根际23.53%,12.62%,39.44%,4.69%。土壤中根际和非根际脲酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际脲酶活性显著高于非根际(p<0.05),过熟林根际脲酶活性与非根际差异并不显著(p>0.05);根际脲酶活性在幼林、中林、成熟林和过熟林分别高于非根际20.97%,41.83%,16.43%,8.47%。土壤中根际和非根际过氧化氢酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际过氧化氢酶活性显著高于非根际(p<0.05),过熟林根际过氧化氢酶活性与非根际差异并不显著(p>0.05);根际过氧化氢酶活性在幼林、中林、成熟林和过熟林分别高于非根际27.57%,14.73%,9.60%,0.31%。

表2 不同林龄油松根际与非根际土壤酶活性 mg/g
2.3 不同林龄油松土壤微生物群落多样性
对于土壤细菌多样性,由表3可知,油松根际和非根际土壤细菌覆盖度、丰富度指数、Shannon-Wiener指数、均匀度指数、ACE、Chao 1和Simpson指数随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,过熟林有所降低。差异性检验表明,油松根际土壤细菌丰富度指数、Shannon-Wiener指数、ACE、Chao 1指数均显著高于非根际(p<0.05);而根际土壤细菌覆盖度、均匀度和Simpson指数与非根际差异并不显著(p>0.05)。
对于土壤真菌多样性,由表4可知,油松根际和非根际土壤真菌覆盖度、丰富度指数、Shannon-Wiener指数、均匀度指数、ACE、Chao 1和Simpson指数随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,过熟林有所降低。差异性检验表明,油松根际土壤真菌丰富度指数、Shannon-Wiener 指数、ACE、Chao 1指数均显著高于非根际(p<0.05);而根际土壤细菌覆盖度、均匀度和Simpson指数与非根际差异并不显著(p>0.05)。

表3 油松根际与非根际土壤细菌多样性
注:同列不同小写字母表示不同林龄差异显著(p<0.05),下表同。

表4 油松根际与非根际土壤真菌多样性
2.4 不同林龄油松土壤微生物群落组成
由图1可知,不同林龄油松根际土壤细菌群落中,优势类群主要是变形菌门(36.29%)、酸杆菌门(28.92%)和芽单胞菌门(7.63%),还包括浮霉菌门(5.19%)、放线菌门(4.36%)、拟杆菌门(0.57%)、后壁菌门(4.19%)、绿弯菌门(5.23%)、疣微菌门(4.07%)。非根际土壤细菌群落中,优势类群主要是变形菌门(46.23%)、酸杆菌门(21.47%)和芽单胞菌门(11.53%)。其中根际土壤细菌酸杆菌门相对丰度高于非根际,变形菌门相对丰度低于非根际。
油松根际土壤真菌群落中,优势类群主要是子囊菌门(56.39%)和担子菌门(24.13%),还包括结合菌门(5.02%)、壶菌门(0.24%)、新丽鞭毛菌门(6.57%)、球囊菌门(2.01%)、芽枝菌门(2.19%)。非根际土壤真菌群落中,优势类群主要是子囊菌门(48.23%)和担子菌门(29.56%),其中根际土壤细菌酸杆菌门相对丰度高于非根际,变形菌门相对丰度低于非根际。
主坐标分析(PCoA)分析表明:油松土壤根际和非根际细菌群落具有很好的相似性,并且根际和非根际细菌群落产生明显的分离效应;油松土壤根际和非根际真菌群落具有很好的相似性,并且根际和非根际真菌群落产生明显的分离效应(图2)。
2.5 土壤养分、酶活性与微生物群落结构的相关性
相关性分析表明(表5):油松土壤细菌Coverage,ACE与土壤养分均没有显著的相关性(p>0.05);土壤pH值与土壤细菌多样性呈负相关,土壤有效磷、有效钾和蔗糖酶活性均没有显著的相关性(p>0.05)。土壤有机碳与细菌Richness,Shannon呈极显著的正相关(p<0.01),与细菌Evenness,Chao 1和Simpson呈显著的正相关(p<0.05);土壤全氮与细菌Richness,Shannon呈极显著的正相关(p<0.01),与细菌Chao 1和Simpson呈显著的正相关(p<0.05);碱性磷酸酶活性与Shannon,Chao 1和Simpson呈极显著的正相关(p<0.01),与Evenness呈显著的正相关(p<0.05);脲酶活性与Shannon,Evenness和Chao 1呈极显著的正相关(p<0.01),与Simpson呈显著的正相关(p<0.05);过氧化氢酶活性与Shannon,Evenness,Chao 1 Simpson呈极显著的正相关(p<0.01)。
油松土壤真菌Coverage,ACE与土壤养分均没有显著的相关性(p>0.05);土壤pH值与土壤真菌多样性呈负相关,土壤有效磷、有效钾和蔗糖酶活性均没有显著的相关性(p>0.05)。土壤有机碳与真菌Shannon,Evenness,Chao 1呈显著的正相关(p<0.05);土壤全氮与Shannon呈极显著的正相关(p<0.01),与Richness,Chao 1和Simpson呈显著的正相关(p<0.05);碱性磷酸酶活性与Shannon,Evenness,Chao 1和Simpson呈显著的正相关(p<0.05);脲酶活性与Shannon,Evenness,Chao 1和Simpson呈显著的正相关(p<0.05);过氧化氢酶活性与Shannon,Evenness,Chao 1和Simpson呈显著的正相关(p<0.05)。

图1 油松根际和非根际土壤微生物群落组成
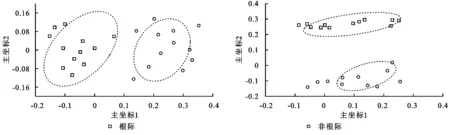
图2 基于Weighted unifrac 距离的土壤微生物群落的主坐标分析(PCoA)
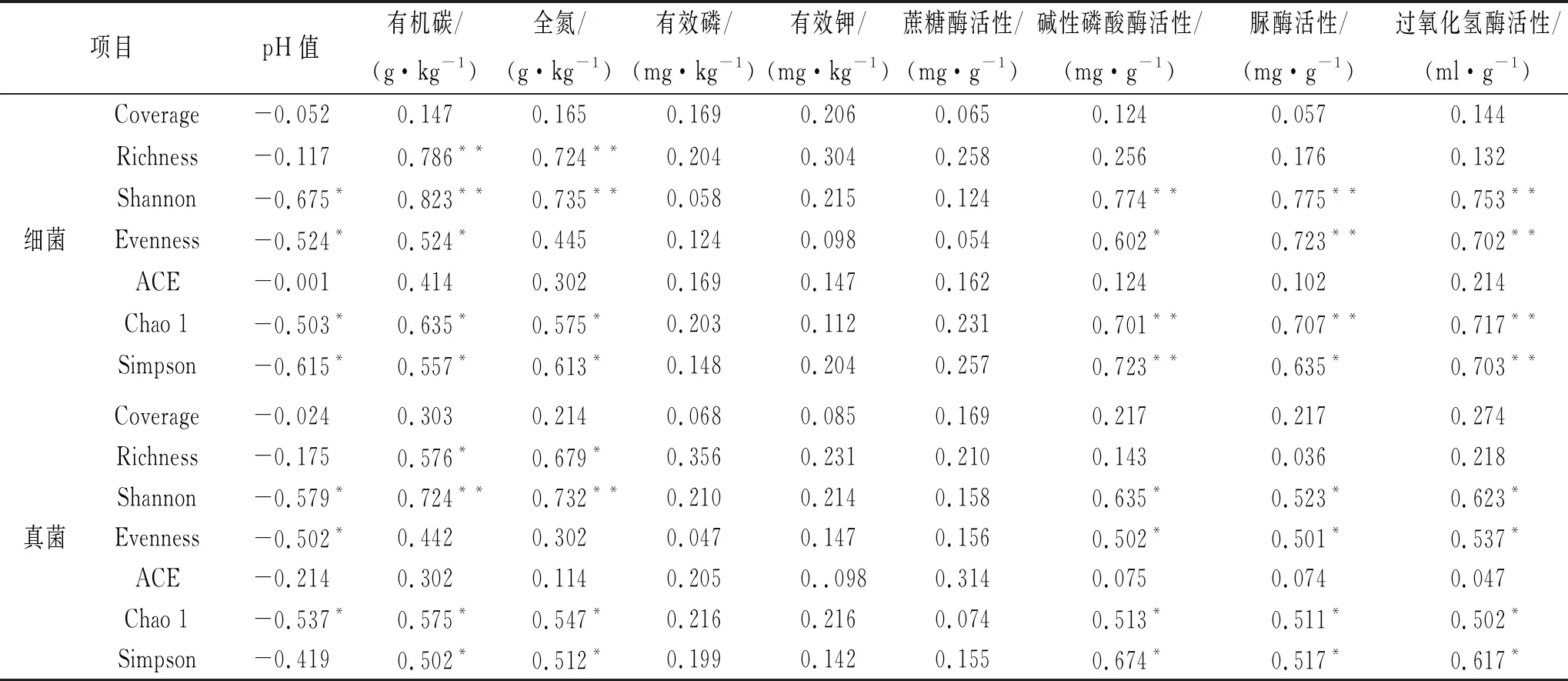
表5 土壤养分、酶活性与土壤微生物群落多样性相关性
注:*,**分别表示在0.05,0.01水平上差异显著。
2.6 土壤微生物群落的RDA分析
蒙特卡罗置换检验结果显示,第一典范轴p<0.05(F=36.59),所有典范轴的p值为0.913,表明该排序模型的解释变量(即土壤环境因子)与响应变量(即土壤细菌和真菌群落结构)的变化关系较为显著。油松根际土壤细菌和真菌群落与土壤环境因子的冗余分析结果见表6。对于土壤细菌群落,RDA前两个排序轴的特征值分别为0.652 4,0.211 7,分别解释了65.24%,21.17%的细菌多样性变化。本文所选的9个土壤环境因子共解释了86%的总特征值,对油松根际土壤细菌群落有显著影响。其中对土壤细菌多样性影响较大的有SOC和STN,而pH值对土壤细菌多样性影响为负。对于土壤真菌群落,RDA前两个排序轴的特征值分别为0.606 8,0.235 6,分别解释了60.68%,23.56%的真菌多样性变化。本文所选的9个土壤环境因子共解释了86%的总特征值,对油松根际土壤真菌多样性有显著影响。其中对土壤真菌多样性影响较大的有SOC和STN,而pH值对土壤真菌多样性影响为负(图3)。

表6 冗余分析结果
3 讨论与结论
水分及养分是大多数植被所赖以生存的生长发育物质,能够保证其获得能量和转换,并影响最终的存活。在这个阶段中,微生物的活动至关重要,承担了陆地有机质分解者的角色,凋落物等腐殖质通过微生物作用得到分解或降解,转化成植被可以吸收的能量和养分,从而促进养分的矿化及分解活动。微生物是土壤生态的重要组成部分,其数量及新陈代谢的变化能够直接作用于土壤,使其结构发生变化,并在一定程度上体现了土壤的肥力及活性。众所周知,林木生长阶段存在不同现象,同样的,微生物多样性也会变化[21-22]。一般而言,油松非根际(p<0.05)土壤pH值明显高于根际,非根际(p<0.05)土壤的全氮、有机碳、有效磷和有效氮均低于根际,非根际和根际土壤中的全氮、有机碳、有效磷和有效氮含量的变化区间一直,即伴随着油松林的增长,呈现先增加后降低的趋势,生长至成熟林时,处于最高值。非根际土壤中微生物多样性和酶活性(碱性磷酸酶活性、脲酶活性、蔗糖酶活性、过氧化氢酶活性)明显低于根际,两者的土壤酶活性变化曲线一致,随着油松林龄的增长,呈现先增加后降低状况,并在成熟林阶段处于最大值,过熟林时开始降低,这反映了根际聚集效应,表明土壤中的有效钾、有效磷含量较高。但是对于幼龄油松来说,其根系并未发育完善,根产物较少,所以主要通过腐殖质分解获得有机质;对于中龄林和成熟林来说,可以通过根产物收获大量的有机质。随着油松林龄的增加,土壤酶活性会呈现“低—高—低”的状态,先降后升,于中龄时达到最高点后再开始下降。研究显示,根际酶活性远远高于非根际酶活性[23-24],油松根系分泌的酶进入土壤会形成极强的酶活性。在微生物作用下,根际的作用范围缩小,微生物分布数量及养分在外界环境变化的情况下,会改变内部结构,直接影响酶的分泌,导致不同土壤区域的酶活性出现较大差异。对于根际或非根际土壤来说,在林龄不断增加的前提下,过氧化氢酶、脱氢酶、脲酶的活性的变化特点为先降后升,并在一定情景下呈现下降趋势,即“低—高—低”的状态。这说明,随着树龄的变化,油松对土壤的作用会发生变化和差异,中龄的油松根系活力更强,能够通过产生更多的分泌物,加速有机质降解,提高微生物活动效率,提高能量转化速度。
土壤和植被具有密不可分的关系,前者为后者生存提供载体和场所,后者能反过来影响和制约前者,土壤中的养分能够促进植被的生长发育,微生物则通过降解凋落物等腐殖质,将其转化为能够得到利用吸收的养分[16-18],能够帮助土壤保持有效的肥力。另外,在这个阶段中,酶所起的作用也不容忽视,能够促进有机质分解,加速土壤的生化反应,提高土壤活性。经研究可知,油松土壤非根际和根际真菌群落和细菌的相似性很高,这两种细菌群落会产生较显著的分离效应。据分析相关资料可知,油松土壤的细菌和真菌多样性与有效钾、有效磷和蔗糖酶活性不存在明显的相关性(p>0.05),油松土壤真菌Coverage,ACE和细菌与土壤养分不存在明显的相关性(p>0.05),微生物的生长特点,决定了过高的土壤pH值对微生物生长和新陈代谢不利,养分及酶活性则对微生物分布多样性具有有利影响,这导致了微生物分布在不同的土壤中会出现较大的差别。在不同的发育阶段,油松对土壤的养分需求会发生变化,也反映在微生物的变化和根际土壤变化方面,凸显了根际的作用。整体来看,根际土壤中分布的微生物数量明显多于非根际土壤。但是油松的生长发育会受到很多因素的影响,根际及非根际土壤还会受到季节更迭的影响,所以这些也需要学界在今后加强具有针对性的研究[25-27]。
土壤微生物群落多样性和组成,受到土壤理化性质和不同生境之间的差异影响,在不同的生境中,根际微生物群落存在很大差别。另外,油松土壤的非根际和根际真菌群落及细菌具有较强的一致性,非根际和根际细菌群落存在显著的分离效应。经分析发现:油松土壤细菌和真菌多样性与土壤pH值存在负比例关系(p>0.05),油松土壤真菌Coverage,ACE和细菌与土壤养分不存在显著的相关性(p>0.05);土壤细菌和真菌多样性与土壤电导率和全磷均不存在显著的相关性(p>0.05)。冗余分析显示:真菌82%和细菌86%的总特征值由9个土壤环境因子分别进行解释,可知油松土壤细菌和真菌多样性收到了土壤环境因子的影响。其中SOC和STN对土壤细菌和真菌多样性影响较大,而土壤细菌和真菌多样性受到pH值的影响为负。
