晚全新世淇澳岛红树林有孔虫记录与古环境意义
2020-07-02陈慧娴王建华董玮琛黄康有张恺陈子豪
陈慧娴,王建华,董玮琛,黄康有,张恺,陈子豪
1.中山大学地球科学与工程学院,广州 510275
2.南方海洋科学与工程广东省实验室,珠海 519000
在隐蔽的低能海岸潮间带区域,植被类型单调,环境受控于潮汐作用[1]。随着高程的增大,盐沼被海水淹没与出露的时间随之减小,在一些对应于潮位特征值的高程处,盐沼受海水淹没和出露的时间有一个突变,从而造成盐沼各种环境特征(植被、沉积物、盐度)均有明显的垂直分带现象[1-2]。这种垂直分带现象使得潮间带沉积物成为高精度古海平面的理想指示物[3]。20世纪80年代末期,加拿大学者发现潮间带盐沼有孔虫分布与平均海平面高程紧密相关,不同的有孔虫组合占据不同植物带,对应不同的高程区间[4]。在加拿大 Nova Scotia(Chezzetcook)盐沼,经典的分带中对应最大高潮位(HHW)的有孔虫组合代表属种为Trochammina macrescens,Tiphotrocha comprimata。这一对应关系十分稳健,在钻孔沉积物中,如果把最高高潮位(HHW)处的有孔虫组合作为精确海平面指示物,可以大大提高有孔虫重建海平面的精度[4]。并且盐沼有孔虫群落以胶结壳为主,分异度远小于河口区域,种群组成比较简单,但有孔虫数量丰富,利于统计,这些特质使得盐沼有孔虫组合对古海平面的指示更加便捷可靠[5-6]。
在亚热带及热带的红树林潮间有孔虫与高程之间的关系也同样适用,也即存在垂直分带性。研究显示在Flordia地区以红树林有孔虫为基础的古海平面重建记录与盐沼有孔虫可以达到同样的精度[7]。在澳大利亚红树林地区胶结壳组合占据了平均高潮线以上的区域,而潮间带下部则以钙质壳为主,指示的高程范围分辨率可达0.8 m[8-9];但是,也有学者认为在季节性降水差异较大的地区,红树林有孔虫组合的变化主要由气候因素控制,而非单纯的海平面变化[10]。总体来看,在世界范围内低纬度红树林地区的有孔虫记录数量仍然较少。
国内针对现代潮间带地区有孔虫及其环境意义的研究见李建芬[11]、王绍鸿[12]和尤坤元[13]等的报道。研究显示在渤海湾潮间带有孔虫可划分为5个亚带与潮汐线对应[11];在福建深沪湾,对比现代潮间带表层样品和钻孔样品中的有孔虫,发现中全新世以来海平面到达钻孔位置以后经历了高潮位—中潮位—低潮位的潮间带环境变化[12];在苏北建川的开敞型盐沼,表层有孔虫受高程控制,分3个带,指示的海平面精度最高可达±5 cm[13]。然而在这些成果中,典型的盐沼胶结质壳有孔虫少见,仅在建川剖面的上部盐沼发现了胖砂轮虫Trochammina inflata(Montagu)、瘦瘪砂轮虫Jadammina macrescens(Brady)、缝裂海伦虫Helenina anderseni(Warren)组合[13],而依据钙质有孔虫划分的带占据高程范围大且与潮下带的属种混合在钻孔中往往难以辨认。这样的有孔虫分布可能是由中国沿海的区域性特征导致,华北、华东沿海地区过高的Ph值使得胶结壳在盐沼中不易保存[14]。再者近现代人类活动强烈,致使原始盐沼生态遭受破坏,也可导致具有指示意义的胶结壳体的盐沼有孔虫少见[15-16]。
本文选取了华南地区保存较好、范围较大的淇澳岛红树林湿地保护区作为取样地点,初步探究了红树林区域内的钻孔QA中有孔虫的属种分布以及壳体埋藏保存状态,并讨论了其对晚全新世古环境尤其是高精度海平面变化的指示意义,评估了淇澳岛红树林有孔虫作为高精度古环境指标的潜力。
1 研究区概况
本次研究的钻孔位于广东省珠海市淇澳岛西北部的珠海淇澳—担杆岛省级红树林自然保护区(图1),保护区临近珠江水系横门河口,为物种丰富的红树林湿地生态系统。淇澳岛面积24 km2,北与虎门相对,东与香港、深圳隔海相望,有淇澳大桥与唐家镇相连。东北与西南部多山,中间平地为居民区,环岛形成多个海湾,其中以石井湾和大围湾面积最大[17]。岛上属南亚热海洋性气候,年平均气温22.4 ℃,最低气温出现在 1 月(平均温度 15.3 ℃),历年极端最低温 2.5 ℃,年平均降水量 1 964.4 mm[18]。淇澳岛夏季以东南风为主,冬季以东北风为主。海水盐度年平均18.2‰,土壤属于滨海盐渍草甸沼泽土,其表土(0~13 cm)全盐量为 20.8‰[19]。
淇澳岛邻近伶仃洋西滩,潮汐属不正规半日潮,其特点是相邻的两个高潮或低潮的潮高不等,涨落潮历时不等,平均潮差随洪季、枯季及大、小潮而不同,一般为0.96~1.77 m。参考附近的横门潮位站资料,平均高潮位为1.98 m,平均低潮位为−1.08 m[20]。外海传入的潮流能通量大部分自南向北在珠江口内汇聚,淇澳岛—内伶仃岛—蛇口半岛区域以北的内伶仃洋四周区域的泥沙呈现顺时针方向水平环绕的环流运移[21],而淇澳岛周围潮流呈逆时针旋转[22],沉积物悬浮浓度在岛屿东南部达到最高[21]。淇澳岛周边床底泥沙较细,主要为粉砂和黏土,砂含量在10%以下[23],主要来源于横门水道来沙以及涨潮时金星门小环流的输沙,海岸滩涂广阔,历史上曾生长大片红树林[24]。
珠江口红树林一般发育在平均海平面或者稍上至大潮平均高潮线之间的潮间带区域[25]。根据野外调查结果,淇澳岛的红树林种类有海桑(Sonneratia caseolaris)、 无瓣海桑(Sonneratia apetala)、银叶树(Heritiera littoralis)、 海 漆(Excoecaria agallocha)、卤蕨(Acrostichum aureum)、木榄 (Bruguiera gymnorhiza)、桐 花 树 (Aegiceras corniculatum)、秋茄(Kandelia candel)等,此外,半红树林呈散生状态,主要种类有海芒果(Cerbera manghasL.)、水黄皮(Pongamia pinnata)等。滩涂群落除上述的红树林群落外,由入侵种形成的互花米草群落(Spartina alternifloraCom.)占地最广,已大面积出现[19,26]。
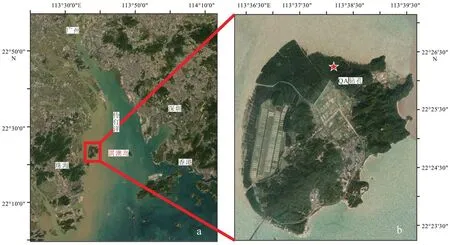
图1 珠江口伶仃洋(a)以及淇澳岛钻孔位置(卫星图)(b)示意图Fig.1 Map of Lingding Bay of Pearl River estuary (a) and location of core QA from Qi’ao Island in satellite image (b)
2 样品与方法
QA钻孔位于珠江三角洲(22°24′37.3″N、113°16′58″E),高程 1.5 m,由 GPS 读取,并且通过附近高程2.0 m的标志点校正,确定钻孔位置高程。取样地点位于平均高潮线以上、平均高潮线以下的红树林泥滩潮间带上部环境,周围覆盖的植被主要为人工种植的无瓣海桑。使用俄罗斯钻手动钻孔方法进行取样,取样率接近100%,取样过程无压实。钻孔取得岩芯长300 cm,岩芯照片见图2,取样间隔为5 cm,共60个样品,每个样品包含5 cm厚度的沉积物。
从钻孔中3个层位取得14C测年材料,样品送至波兰的Poznań放射性同位素实验室进行AMS14C测年,测年结果利用 CALIB v.7.10[27]进行校正,其中碳屑样品采用Intal13曲线,贝壳样品采用Marine13曲线进行校正,并且贝壳样品同时进行海洋碳库效应修正,采用参数(∆R)为-128±40[28-29]。
有孔虫样品取样间隔为10 cm,每个样品包含的沉积物厚度为5 cm,共取得30个样品。将沉积物置于70 ℃的烘箱中烘干48 h,取出后称量并记录干重,接着在烧杯中加水浸泡,待样品分散之后用63 μm孔径筛网过筛保留大于63 μm粒径的筛上部分,置入烧杯中进烘箱烘干。在Olympas SZ61体视显微镜下对烘干后的样品中的有孔虫壳体进行观察与统计,记录样品中的有孔虫种属、个数。并计算样品的有孔虫多样性和丰度。
沉积物粒径分析取样间距为5 cm,每个样品包含的沉积物厚度为5 cm,共取得60个样品,实验分析仪器为马尔文激光粒度仪Mastersizer 2000。取2 g左右干样加入10 mL浓度为10%的HCl溶液和10 mL浓度为10%的H2O2溶液搅拌,充分反应后,洗至中性,加入 10 mL 浓度为 0.1 mol/L 的 Na6O18P6溶液并充分搅拌分散,静置24 h,将样品上机进行粒度测试。
灰度首先采用白色卡校正钻孔沉积物的颜色,然后采用Image J软件,由钻孔沉积物照片连续获得灰度数值。灰度值为0表示颜色为全黑,灰度值为255表示颜色为全白,即灰度值越低,则样品颜色越深,反映有机质含量越高;反之,则反映有机质含量越低[30]。

图2 钻孔岩性照片及深度标尺Fig.2 Pictures of core sediment with scale of depth
3 结果
3.1 钻孔岩性和年代
钻孔总长300 cm,根据室内岩性观察结合粒度数据,钻孔深度300~220 cm的沉积物为均匀的灰色黏土质粉砂(图3),有丰富的贝壳碎片及腹足类壳体出现。沉积物中黏土、粉砂、砂含量平均值分别为21%、75%、4%,中值粒径变化范围为6.0~6.8Φ,平均值为6.5Φ;220~180 cm为灰色的黏土质粉砂夹丰富的植物碎屑,沉积物组份中黏土、粉砂、砂含量平均值分别为19%、72%、9%,中值粒径变化范围为5.6~6.7Φ,平均值为6.1Φ,较下部沉积层粒度稍粗;180 cm以上为含大量植物碎屑的黄灰色砂质粉砂,砂组分含量显著升高,上升至21%,黏土、粉砂含量平均值分别为15%、64%,中值粒径变化范围为4.5~6.9Φ,平均为5.5Φ,较下部沉积层粒度明显变粗,其中在深度130~90 cm和60~40 cm处夹有两层细颗粒的灰色黏土质粉砂沉积。沉积物颜色显示出清晰的潮汐层理,且该段沉积物含有多层深灰—黑色的富泥炭黏土层,其中深度200~190 cm、145~135 cm、70~60 cm 为黑色富泥炭黏土层。沉积物的灰度值与岩性层理变化有较好的对应,深度300~180 cm的灰色粉砂质黏土灰度值变化范围为82~103,平均值为94;深度180 cm到顶部的灰度变化范围较大,为56~145,平均值为102,其中黑色富泥炭黏土层位对应着灰度谷值。
测年结果详见表1。钻孔底部深度为295 cm处用有机质泥获得的校正年龄为 3 887±165 cal.aBP,在孔深为195和140 cm处分别由有机质泥和贝壳获得的校正年龄分别为 2 996±80 cal.aBP、1 024±64 cal.aBP。其余年龄由 Bacon 所建立的年代-深度模型内插和外延获得(图4)。模型显示钻孔沉积物约为 4 200 cal.aBP 以来的沉积。
3.2 钻孔QA有孔虫记录
经镜下鉴定后统计结果显示QA钻孔连续分布有孔虫壳体,共发现有孔虫16属29种,主要属种见图5。该钻孔有孔虫丰度变化范围为7~900个/20g,差异显著,分异度变化范围为2—12种。根据有孔虫百分含量聚类结果,钻孔QA的有孔虫类群被显著分为两组(图6),与钻孔岩性变化对应,其中Ⅰ组为钙质壳有孔虫组合,对应灰色黏土质粉砂(中值粒径均值6.5Φ),Ⅱ组主要包含胶结壳有孔虫,对应含植物碎屑的灰黄色砂质粉砂(中值粒径均值5.5Φ)。各有孔虫分组属种组成如下:
Ⅰ组:埋深300 ~ 180 cm (4 200~ 2 400 cal.aBP),该组有孔虫优势属为Ammonia,平均占整个群落的80.9% (变化范围70.3%~98.5%),包括的种有Ammonia becсarii,Ammonia tepida,Ammonia pauciloculata,Ammoniaconvexidorsa,Ammonia aberdoveyensis,此外还含有一定数量的Proelphidium granosum(0~14.8% ,均 值6.6% )和Elphidiella kiangsuensis(0~19.0%,均 值 5.6%),Cribrononion vitreum(0~23.1%,均值 2.2%),以及少量的Elphidium simplex,Elphidium hispidulum,Florilus limbatostriatus,Rectoelphidiella lepida,Helenina anderseni,含量均小于 5%。其中 300~220 cm (4 200~3 300 cal.aBP)有孔虫丰度值稳定,为全钻孔最高,变化范围为408~900 个/20 g,平均值为 604 个/20 g;分异度也较高,变化范围为8~12,平均值为10.0。但是220~180 cm(3 300~2 400 cal.aBP)层位中,分异度显著降低,平均值为6.3。该组样品间有孔虫群落组成相似度高,聚类组间距离<2。

图3 钻孔QA岩性、沉积物组分、灰度值、中值粒径随深度的变化曲线(灰色阴影表示3个深色泥炭层的位置)Fig.3 Vertical variation of lithology,sediment fraction,gray scale,median grain size with depth from core QA( The three gray shadow layers indicate peat horizons)

表1 钻孔QA沉积物AMS14C测年结果Table 1 AMS14C radiocarbon dates from sediment of core QA

图4 钻孔 QA 年代-深度模型(Bacon)[31]Fig.4 Age-depth model established with Bacon for core QA[31]
Ⅱ组:埋深180~0 cm,总体来说该段的优势种为胶结壳Arenoparella mexicana,占群落总体比例高达74.7%。其他胶结壳属种Miliammina fusca,Trochammina inflata和Haplophragmoidesspp.也是群落重要构成部分,但是该组样品中以Ammonia becсarii为主的组合在某些层位中也间杂分布,样品间差异显著,可细分为5个带。
(1)带Ⅱa,埋深 180~130 cm (2 400~1 022 cal.aBP),除了样品165 cm以外,有孔虫带以Arenoparellamexicana为绝对优势种(32.3%~71.4%,平均值50.6%),胶结壳Miliammina fusca(13.2%~29.0%,均值 17.5%),Trochammiasp.(0~14.3%,均值 6.8%),Haplophragmoides manilaensis(0~6.6%,均值 3.3%),Haplophragmoides welbrite(0~9.6%,均值 2.4%)以及Jadammina macrescens(0~13.3%,均值 6.9%)。样品165 cm中有孔虫组成与组Ⅰ相似,以Ammoniabecсarii(81.7%)为优势种,混有少量Arenoparella mexicana(7.5% ) ,Miliammina fusca(2.2% )和Proelphidium granosum(7.4%)。丰度和分异度为全钻孔最低,平均值分别为25.6个/20 g和5.75。
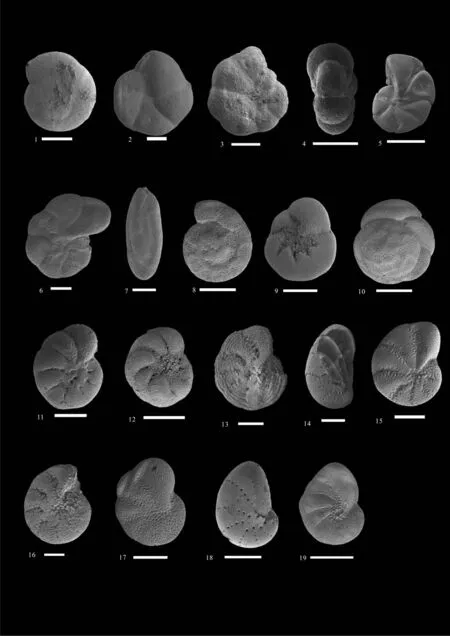
图5 有孔虫扫描电镜照片 (比例尺=100 μm)Fig.5 Scanning Electron Microscope pictures of foraminifera (scalebar=100 μm)1,2.Arenoparrella mexicana (Kornfeld,1931); 3,4.Haplophragmoides manilaensis (Andersen,1952); 5.Trochammina sp.;6.Jadammina macrescens(Brady,1870); 7.Miliammina fusca (Brady,1870); 8,9.Ammonia beccarii (Linné,1758); 10.Ammonia aberdoveyensis (Haynes,1973);11.Cribrononion subincertum (Asano,1951); 12.Proelphidium granosum (d’Orbigny,1846); 13.Elphidium hispidulum (Cushman,1936);14.Rectoelphidiella lepida (He,Hu & Wang,1965); 15.Elphidium sp.(Williamson,1858) ;16.Elphidium margaritaceum (Cushman,1930);17.Elphidiella kiangsuensis (He,Hu & Wang,1965); 18.Cribrononion vitreum (Wang,1980); 19.Florilus limbatostriatus (Cushman,1931)
(2)带Ⅱb,埋深 130~90 cm (1 022~729 cal.aBP),该有孔虫带以Ammonia becсarii(38.0%~81.0%,均值64.7%)为优势种,次要种有Arenoparellamexicana(0.9%~20.8%,均值 7.2%),Rectoelphidiella lepida(0~26.0%,均 值7.1%),Miliammina fusca(0~18.3%,均值5.3%)以及少量的Proelphidium granosum(4.6%),Elphidiella kiangsuensis(2.1%)和Elphidium hispidulum(2.2%)。该段丰度平均值为209.0 个/20 g,分异度为 8.5。
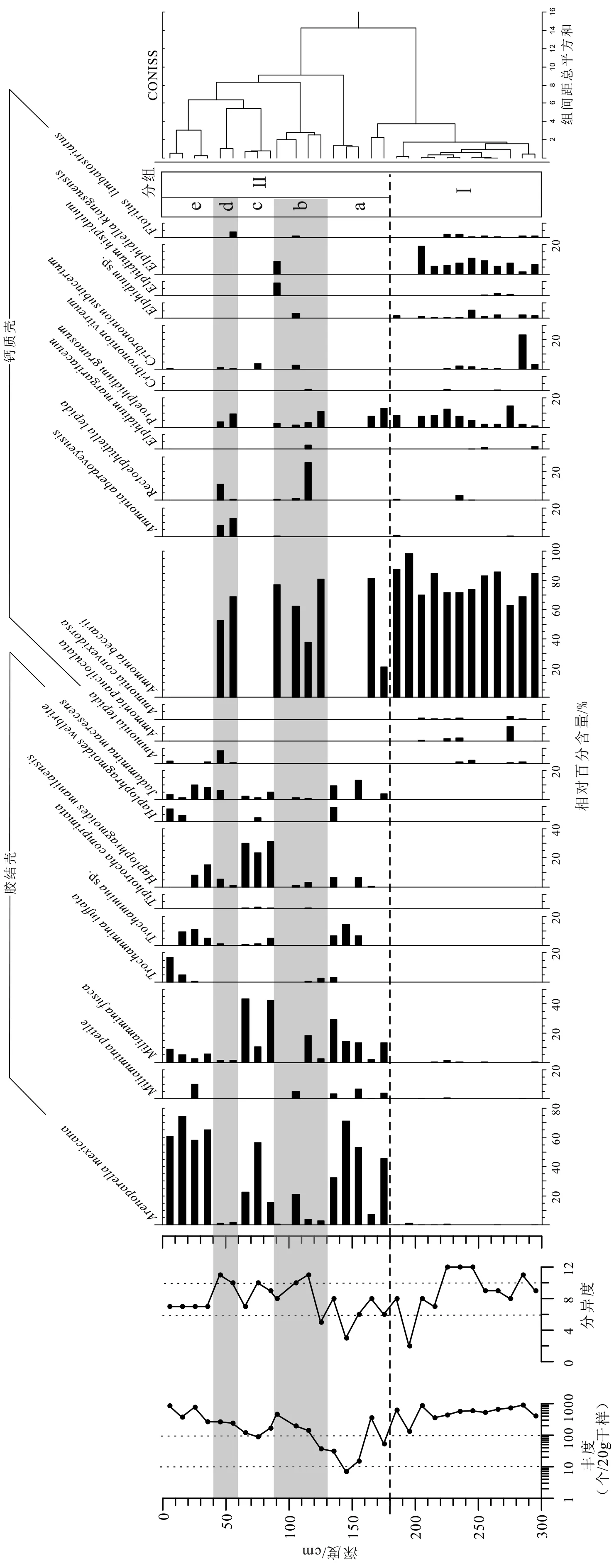
图6 钻孔QA有孔虫丰度,分异度变化,各属种所占百分比及有序聚类结果(所用软件为Tilia 2.0[32])Fig.6 Variation of foraminiferal abundance,diversityand clusters based on stratigraphy cluster (Produced by Tilia 2.0[32])
(3)带Ⅱc,埋深 90~60 cm (729~508 cal.aBP),有孔虫带与Ⅱa类似,以Arenoparella mexicana(15.6%~56.5%,平均值31.5%)和Miliammina fusca(10.6%~43.3%,平均值32.0%)为优势种。次要种为Haplophragmoides manilaensis(23.5%~31.3%,平均值 28.3%),还有Trochamminasp.,Haplophragmoides welbrite以 及Jadammina macrescens,Cribrononion vitreum等种属含量不超过5%。该段丰度平均值为125.3 个/20 g,分异度为 8.6。
(4)带Ⅱd,埋深 60~40 cm (508~58 cal.aBP),该有孔虫带仅包含两个样品,Ammonia becсarii为优势种,含量约为75.4%,次要种有Rectoelphidiella lepida(6.0%),Proelphidium granosum(6.4%),以及少量的Haplophragmoides manilaensis(3.3%)以及Jadammina macrescens(3.0%)。该段丰度平均值为 253.0个/20 g,分异度为10.5。
(5)带Ⅱe,埋深 40~0 cm (358~0 cal.aBP),有孔虫带以Arenoparella mexicana为绝对优势种(58.2%~ 74.7%,平均值 64.7%),次要种有Miliammina fusca(2.3%~8.8%,平均值5.6%),Trochammina inflata(0~16.8%,5.5%),Trochammiasp.(0~11.0%,平 均 值 6.5%),Haplophragmoides manilaensis(0~15.1%,平均值5.8%),以及Jadammina macrescens(1.1%~9.8%,平均值5.6%)。含量少于5%的属种有Miliammina petile(2.5% ) 和Haplophragmoides welbrite(3.1%)。该段丰度平均值为 556.3 个/20 g,分异度为7。
4 讨论
4.1 有孔虫属种分布与指相意义
钻孔有孔虫分析显示,有孔虫组合显著分为两大类,其中带Ⅰ以钙质壳为主,丰度和分异度较高。其中以Ammonia becсarii(78%)为优势种,次要种 有Proelphidium granosum(6.6%)和Elphidiella kiangsuensis(5.6%),以及少量的Elphidium simplex,Elphidiumhispidulum,Floriluslimbatostriatus,Rectoelphidiella lepida,Cribrononion subincerium,Cribrononion vitreum。有孔虫组合与现代伶仃洋以Ammonia becсarii和Proelphidium granosum为特征种的河口浅滩相类似,这一组合分布在半咸水、水深小于10 m的浅滩区域[33-35]。结合该段岩性为均匀的黏土质粉砂沉积,包含少量贝壳碎屑,由此认为Ⅰ段沉积物为河口潮下带沉积。有孔虫带Ⅱ的优势种为胶结壳,丰度和分异度较低,并且丰度有随埋藏深度增加而减小的趋势。其中以Arenoparella mexicana为主,占群落总体比例高达74.7%,同时其他胶结壳属种Miliammina fusca(17.5%),Trochammina inflata(6.8%)和Haplophragmoidesspp.(6.6%)也是群落重要构成部分。以Haplophragmoides Shenzhenensis(62.0%)为主,Ammoniacf.beccarii(17%)和Ammonia tepida(12%)为次要种属的有孔虫组合曾见于深圳湾红树林泥滩[36]。尽管优势种不同,但是这样以胶结壳为主的有孔虫组合与淇澳岛类似,并且可与现代盐沼、红树林地区的有孔虫记录进行对比。
在澳大利亚的红树林上部,Haplophragmoidesspp.作为次要种与Trochammina inflata组成了高高程带(高高潮线以上)的代表性组合[37]。在经典的有孔虫分带中高盐沼组合为Jadammina macrescens和Trochammina inflata[4],并且这一组合也可见于大量的潮间带盐沼有孔虫文献中[38-40]。Arenoparella mexicana种主要占据了高潮线附近的中高盐沼区域,分别与Miliammina fusca,Trochammina inflata种属组合在北美的新泽西州,北卡罗莱纳州的Albemarle-Pamlico河口盐沼地区出现[6,41]。以Miliammina fusca为优势种的有孔虫组合,及少量的Ammobaculitesspp.,是北美典型的低盐沼代表种[42-44],占据平均海平面以上高潮线以下的盐沼环境。结合钻孔QA顶部样品中的有孔虫组合,有孔虫分带Ⅱe组Arenoparellamexican,Haplophragmoidesspp.和Trochammina inflata组合位于淇澳岛现代红树林的高潮线以上区域,而分带Ⅱa和Ⅱc含有更高含量的Miliammina fusca则可能位于高潮线以下、平均海平面以上的潮间带区域。
在现代盐沼及红树林有孔虫研究中,以Ammonia属为主的钙质壳占据平均海平面附近的泥质光滩,而胶结壳占据潮间带中上部的垂直分带现象并不少见。除了上文提到的北美地区,澳大利亚大堡礁地区的红树林分为低、中、高3个带[9],最低的带优势种为Ammonia tepida,Elphidium discoidale multiloculum,中间带为Miliammina fusca,最高的为Trochammina inflata;印度的Sundarbans红树林中低盐沼沉积中以钙质壳Quinqueloculina,Ammonia属为主,而高盐沼以胶结壳Jadammina macrescens,Trochammina inflata,Miliammina fusca为主,由此可见有孔虫的垂直分带性在红树林中也普遍存在[45]。这样典型的有孔虫属种分带组合使得珠海淇澳岛的红树林有孔虫具有高精度重建相对古海平面的潜力。
4.2 有孔虫组合与古环境变化
红树林湿地的水动力条件和生物化学环境与河口沉积环境差异显著,湿地内部植被繁多,使得湿地内部水动力较弱,多形成粉砂质—泥质的湿地海岸,但研究区离淇澳岛东南部丘陵地形较近,受径流影响,向陆一侧可能搬运粗粒的花岗岩风化产物,使得红树林沉积相含砂量较潮下带丰富[46-47]。潮汐周期性的浸没与暴露,使红树林内的孔隙水的地球化学性质异常,且丰富的植物碎屑经过埋藏作用,在水体中分解形成腐殖酸,造成水体低pH值,从而不利于钙质壳有孔虫的生存与壳体保存[14,48],因此在不同的潮汐区间形成截然不同的有孔虫分带(Ⅰ和Ⅱ)。
QA 钻孔记录了全新世约 4 200 cal.aBP 以来的沉积(图7),至 3 300 cal.aBP,研究区沉积均质的灰色黏土质粉砂,稳定的钙质壳有孔虫组合反映了潮下带环境。3 300~2 400 cal.aBP(220~180 cm)对应着潮下带转变为红树林沉积相的过渡带,沉积物中植物碎屑增加,有孔虫组合仍以Ammonia becсarii为主,有孔虫的分异度急剧降低,且出现溶蚀现象,平均值由10种降低至6种,丰度也有明显波动,说明沉积环境海相性减弱,水体盐度降低,同时沉积物粒径也较下部潮下带沉积更粗,含砂量更高,说明水体变浅,受径流影响增强,说明沉积物淤积至海平面以上,区域内红树植被开始发育。有孔虫组合由Ⅰ段的钙质壳变为Ⅱ段胶结壳为主,反映了在约 2 400 cal.aBP(钻孔深度为 180 cm)前,研究区由河口潮下带逐渐演变为红树林的潮间带中上部,沉积物粒度平均值由6.4Φ增大至5.6Φ佐证了沉积环境的变化。这一沉积环境的转变与文献中淇澳岛北偏西侧的钻孔的记录一致,该钻孔指示在约4 200~2 500 aBP 期间 C/N 显示有机质来源以海源为主,2 500 aBP之后则变为以陆源为主,且有机碳、氮含量显著增大[18],印证了红树林的发展。若不考虑沉积压实作用,则指示平均海平面在此时曾位于-180 cm的位置,结合钻孔顶部高程为1.5 m,即与现今海平面相比低约30 cm。
前人在珠江三角洲及伶仃洋钻孔中的由海平面标志层位所重建的海平面曲线显示,晚全新世以来(约 3000 cal.aBP)海平面接近现今高度,保持稳定[49-50]。此时三角洲盆地充填完毕,沉积物向海淤积,前积作用加强,导致岸线向海迁移[51-53],造成了大片平缓的滩涂,当光滩淤积至平均海平面以上,则为红树林的发育提供了充足的空间[25]。番禺万顷沙钻孔GZ-2孢粉分析表明红树孢粉在此阶段达到全新世最盛[54],而在整个珠江三角洲盆地草本花粉和莎草科种类花粉的增多成为晚全新世的主要特征,说明了随着相对海平面下降,珠三角盆地广泛发育草地和湿地沼泽[52,55],本研究的结果与此背景相符。在约360 cal.aBP,有孔虫组合由以Arenoparella mexicana和Miliammina fusca为优势种的潮间带中下部(Ⅱa和Ⅱc)变为以Arenoparella mexicana为绝对优势种,含Haplophragmoides manilaensis和Jadammina macrescens的潮间带上部组合(Ⅱe),说明了红树林的促淤作用使得泥砂不断淤积,导致沉积物垂向加积,致使红树林向海迁移,而原先的潮间带中下部演变成了潮间带上部。
深度 180~130 cm 之间,有孔虫丰度由 495个/20 g降低至25.6个/20 g。除了由沉积环境变化引起的有孔虫种群差异以外,即水体稳定的半咸水河口相比地化条件异常的红树林相有孔虫丰度更高以外,壳体的保存与埋藏作用也不可忽视,因为相对于钻孔130 cm以浅的红树林沉积,该段的有孔虫丰度异常偏低,且发现钙质有孔虫遭受溶蚀,仅剩有机质膜保存的现象。从沉积物岩性来看,深灰色的黏土质粉砂,可见潮汐层理,与下伏地层的岩性连续一致,说明这一现象与气候无关。可能是在红树林发展的初期,红树植物根系及死亡的植物残体埋藏腐殖化之后形成的酸性环境使得下伏泥滩中的钙质有孔虫溶解,因此引起了位于带Ⅰ和带Ⅱ过渡地层中的有孔虫壳体的低保存状态。这一现象在澳大利亚的Cocoa Creek红树林中也有发现[56]。因此,也佐证了研究区内红树林在这一时期发育。
4.3 红树林沉积记录对短尺度事件的响应
近 2 400 cal.aBP 以来海平面趋势平稳,研究区为中高红树林潮滩,但是在胶结壳有孔虫组合中仍可见钙质有孔虫组合(图6,7),指示了潮下带河口相或者潮间带光滩的沉积物来源。在细分的有孔虫带Ⅱb (埋深 130~90 cm)和带Ⅱd(埋深60~40 cm)仍以Ammonia becсarii(> 60%)为优势种,含量超过5%的次要种有Rectoelphidiella lepida(7.1%),Proelphidium granosum(4.6%),Arenoparella mexicana(7.2%),Miliammina fusca(均值 5.3%),还含有少量的Elphidiella kiangsuensis(2.1% )和Elphidium hispidulum(2.2%)。这两段沉积对应的年代为1 022~729 cal.aBP 和 508~358 cal.aBP,沉积物粒径较细,砂含量显著降低,有机物的堆积明显减少。从本钻孔的粒度数据看,潮下带的河口相沉积较潮间带的沉积物粒度更细。推测可能形成这两个沉积带的原因有:一是短暂的海平面上升,使得沉积物性质更偏向潮间带下部的特征;二是短时气候事件导致的潮下带沉积物快速堆积。
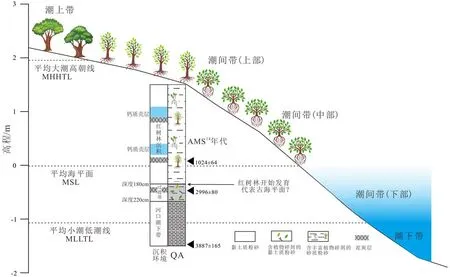
图7 淇澳岛红树林潮滩环境对应潮汐高程及古环境演变示意图(据QA孔)Fig.7 Sketch map of tidal datum and mangrove tidal flat from Qi’ao Island,and its paleoenvironmental evolutions revealed by core QA
尽管前人根据海平面标志沉积层重建的海平面曲线显示晚全新世海平面大体相对稳定[50],但是3 000 cal.aBP 以来海平面标志点数量偏少,次一级的海平面波动难以被记录到。不少区域性的研究显示在此期间海平面有过小幅度的波动,雷州半岛的海滩沉积序列指示在距今1 200年,海平面比现今至少高128 cm[57],结合珠江口的潮差来看,足以使得部分潮间带地区淹没成为潮下带。而据历史文献资料记载,晚全新世 2 000、1 300 aBP 左右出现过两次较长时间的高海平面时期,当时海平面高出现今约100 cm,造成广州城从番禺迁至顺德[58]。不同时期的海滩岩分布和高程大致可以反映当时的海平面高度,而华南沿海的海滩岩分布也揭示了晚全新世可能存在次一级的海平面波动,2 000 aBP的时候曾经出现峰值[59]。
在泥炭环境中,灰度值与有机质含量、腐殖化具有较强的相关性,可以作为气候变化的替代指标[60]。而泥炭腐殖化程度受植被数量、温度、湿度等控制,腐殖化程度高说明气候温暖、湿润,湿地植被盖度高,残体堆积多,细菌分解作用强[30,61]。在QA孔的红树林沉积物中灰度值向钻孔顶部逐渐变大,说明泥炭腐殖化程度变低,也即晚全新世2 500 a以来气候逐渐向凉干发展,与珠江三角洲孢粉记录所反映的趋势一致[54,62]。而全钻孔灰度值最低,也即颜色最深的层位,深度为 200~190 、145~135 、70~60 cm,说明了沉积物有机质含量高,腐殖化程度高,反映了晚全新世气候较为暖湿的几个时期,对应年代分别为约 3 000~2 700、1 200~1 100、600~500 cal.aBP,这些温暖潮湿的气候阶段与有孔虫和粒度反映的海相性较强的阶段有较好的一致性(图7),基本出现在高海平面之前的200年内。
当然,这两个钙质壳层可能是极端气候条件下的快速沉积,如台风导致的风暴潮沉积。红树林植被有利于风暴扰动后的悬沙沉积,台风过境之后,光滩的细颗粒沉积物发生侵蚀被搬运至植被覆盖的区域沉积[63],形成了细粒沉积层包含有潮下带有孔虫。但风暴潮沉积一般会形成下部侵蚀界面、粒度递变层理,这些在本剖面中并未出现。此外,风暴潮沉积所形成的沉积层厚度与台风的大小等级相关,Ⅱb带的厚度达到50 cm,因此一次风暴潮是否能够沉积如此厚的沉积层尚值得怀疑。
5 结论
(1)淇澳岛红树林QA钻孔中鉴定出两大类有孔虫组合,一是以钙质壳为主的、丰度和分异度较高,优势种为Ammonia becсarii(78%)为优势种,次要种有Proelphidium granosum(6.6%)和Elphidiella kiangsuensis(5.6%),另一个是以胶结壳为主的、丰度和分异度较低,优势种为Arenoparella mexicana,占群落总体比例高达74.7%,同时含其他胶结壳属种Miliammina fusca(17.5%),Trochammina inflata(6.8%)和Haplophragmoidesspp.(6.6%)。
(2)QA钻孔的沉积物可划分出两个沉积环境变化阶段:4 200~2 400 cal.aBP 为以钙质壳有孔虫为主的河口相潮下带后过渡为潮间带下部的光滩沉积,沉积物粒径较细;2 400 cal.aBP 至今是平均海平面(MSL)至平均高潮线(MHW)之间的潮间带红树林沉积环境,沉积物粒径变粗,砂含量增加。沉积环境总体上的演变与三角洲快速前积、滩涂不断发育有关。
(3)QA 孔自 2 400 cal.aBP 以来的潮间带红树林沉积相中夹有两层钙质有孔虫的细粒沉积物,深度为 130~90 和 60~40 cm,对应年代为 1 022~729和 508~358 cal.aBP,沉积物性质和有孔虫组合的变化推测可能与晚全新世短时间海平面波动上升有关。
(4)钻孔灰度值反映了红树林沉积物的腐殖化程度,腐殖化程度最高的3个层位为200~190 、145~135、70~60 cm,对应晚全新世气候较为暖湿的几个时期,年代分别为约 3 000~2 700、1 200~1 100、600~500 cal.aBP, 这些温暖潮湿的气候阶段与有孔虫、粒度反映的海相性较强的阶段比较接近。