CD47相关信号通路及其与相关疾病关系的研究进展
2020-06-27葛闯胡加亮徐寒梅
葛闯, 胡加亮, 徐寒梅
(中国药科大学 江苏省合成多肽药物发现与评价工程研究中心,江苏 南京 211198)
整合素相关蛋白CD47最初发现于无Rh抗原性溶血性贫血患者体内,其血红细胞缺乏CD47[1]。前期研究发现,CD47 与整合素 αvβ3,αⅡbβ3和 α2β1相互作用,调节整合素功能和调节细胞对含有精氨酸-甘氨酸-天冬氨酸的胞外基质蛋白的应答,因而被称作整合素相关蛋白[2]。CD47在多种肿瘤中表达升高,如卵巢癌、白血病、乳腺癌、结肠癌、膀胱癌、胶质母细胞瘤、肝细胞癌和前列腺肿瘤等[3-5]。研究表明,抗CD47抗体能够刺激巨噬细胞清除肿瘤细胞[5],因而CD47成为近年来研究的热点。
CD47通过与胞外、胞内以及其他的跨膜蛋白的横向相互作用发挥不同的功能,介导了不同的信号转导通路,这些信号通路在肿瘤、血管类疾病、自体免疫性疾病等多种疾病中发挥着不同作用。
1 CD47结构、表达与相互作用
1.1 结构
CD47属于免疫球蛋白超家族,有2种异构体,一种含有N糖基化修饰,另一种在其64位和79位丝氨酸含有糖胺聚糖修饰[7]。CD47结构包含3个部分,N末端胞外免疫球蛋白多变(IgV)结构域、高度疏水的5次跨膜片段和1个短的多变C末端胞内段[6],如图1所示。
CD47通过胞外段IgV结构域与其配体凝血栓蛋白1(thrombospondin 1,TSP1)和信号调节蛋白α(signal regulatory protein alpha,SIRPα)相互作用,IgV结构域被N糖基化和O-连接型糖胺聚糖修饰,糖胺聚糖修饰是CD47与TSP相互作用所必需的[7]。IgV结构域上的氨基酸残基Cys33与跨膜区最后一个胞外环上的氨基酸残基Cys263之间存在长二硫键,这一二硫键对于CD47配体的结合、信号转导以及定位到脂筏都是必要的[8]。胞内段存在4种可变剪接体形式,根据基因结构、多肽序列不同,分为Ⅰ型、Ⅱ型、Ⅲ型和Ⅳ型,氨基酸数量从3 ~ 36个不等。Ⅱ型是目前最主要的、表达最多的异构体,其次是Ⅳ型,Ⅳ型主要在脑部和外周神经系统表达,而I型只在角质细胞中显著表达,Ⅲ型主要与记忆巩固相关[9-10]。
1.2 表达
CD47在几乎所有的细胞表面都有表达,包括红细胞和血小板,其表达水平因机体的免疫状态或病理状态不同而改变。CD47在多种肿瘤中表达较高,并且通过阻断CD47能够增强肿瘤的吞噬作用[3-5]。Xu等[11]发现抗CD47治疗可增加巨噬细胞的吞噬功能,降低肿瘤重量,并抑制骨肉瘤异种移植模型中的自发转移。Zhang等[5]发现阻断CD47可增强巨噬细胞对肿瘤细胞的吞噬作用,减轻肿瘤负荷,提高胶质母细胞瘤患者的存活率。同样地,Yoshida等[12]也发现通过阻断CD47能增强巨噬细胞对胃癌细胞的吞噬作用。以上均表明CD47可能是肿瘤细胞的一个重要生存信号分子,与肿瘤发生发展密切相关[3]。在免疫应答过程中,CD47在CD4效应T细胞表面表达升高,防止记忆性T细胞被巨噬细胞清除,延长生存周期[13]。同样,造血干细胞(hematopoietic stem cells,HSCs)表面CD47高表达也使其免受巨噬细胞吞噬,而当CD47在CD34+CD38-HSCs表面表达选择性下调时,就有可能发生噬血细胞综合征(hemophagocytic lymphohistiocytosis,HLH)[14]。
1.3 相互作用
CD47通过胞外IgV结构域主要与3种胞外配体相互作用,分别为CD47-TSP1、CD47-SIRPα及CD47-CD47。由于CD47胞内结构域较小,只有有限的几种胞内信号蛋白与CD47相互作用,主要是促凋亡调节蛋白(BCL2/adenovirus E1B 19 kDa protein-interacting protein 3,BNIP3)以及连接整合素相关蛋白和细胞骨架蛋白(protein-linking integrinassociated protein and cytoskeleton,PLIC)。而CD47与其他跨膜蛋白的横向相互作用可能在CD47的信号转导通路中发挥着重要作用,如整合素αvβ3、血管内皮细胞生长因子受体2(vascular endothelial growth factor receptor 2,VEGFR2)、Rh血型抗原肽复合物、CD47、自杀相关因子(factor associated suicide,Fas)、CD14 等。
1.3.1 胞外相互作用 首先被发现的CD47内源性配体就是TSP1[15]。TSP1属于TSP家族成员之一,这一家族除了TSP1之外,还包括TSP2 ~ TSP5。TSP1是一种瞬时表达的细胞基质蛋白,在不同环境中,通过与细胞表面受体或其他的胞外基质成分相结合调节细胞功能[16]。TSP1结构很大,包含多种结构域,使其能与多种细胞表面受体和胞外基质相互作用,包括整合素(主要是β1和β3)、CD47、CD36、硫酸乙酰肝素蛋白多糖、低密度脂蛋白相关蛋白1和极低密度脂蛋白受体等[16]。TSP1通过其碳末端结合结构域(C-terminal binding domain)上的VVM序列与CD47相结合,进而影响多种细胞功能,如细胞迁移和黏附、细胞增殖和凋亡、调节血管生成和炎症反应等[17]。
第2个被发现的内源性配体是SIRPα[18]。SIRPα属于免疫球蛋白超家族,是一种跨膜蛋白,表达在骨髓细胞表面,如单核细胞、巨噬细胞、粒细胞和CD4阳性树突状细胞,以及在中枢神经系统神经元细胞中以不同水平表达。SIRPα在吞噬细胞表面表达水平似乎非常稳定,不会受到炎症水平的影响,而CD47的表达却随不同的免疫状态或疾病的改变而改变[19]。SIRPα胞外N末端包含3个Ig样结构域,1个单一的跨膜结构域,胞浆C末端含有4个酪氨酸残基,带有2个免疫受体酪氨酸抑制基序(immunoreceptor tyrosine-based inhibitory motif,ITIM),ITIM是酪氨酸磷酸酶SHP(src-homology domain 2-containing protein tyrosine phosphatase)-1和SHP-2的结合位点,ITIM介导SHP-1和SHP-2的募集和激活,使得磷酸化蛋白底物去磷酸化,从而影响胞内信号通路转导,如抑制生长因子受体和其他的一些促酪氨酸激酶活性受体的信号转导[19]。SIRPα通过其N末端IgV结构域与CD47的IgV结构域结合,SIRPα与CD47之间的相互作用在控制细胞吞噬中发挥作用,能够产生一种“不要吃我”的信号[20]。
除了与TSP1和SIRPα结合,也有研究表明,细胞间发生黏附需要CD47的存在而并不需要其配体,表明2个细胞上CD47的IgV结构域也存在着同型结合[21]。不过这种CD47与CD47相互作用能否产生信号转导还缺乏足够的证据。
1.3.2 膜表面相互作用 CD47首先被发现与整合素αvβ3存在相互作用[2],后续又发现其与多种整合素亚型相互作用,如 αⅡbβ3、α2β1、αLβ2、和 α4β1[22-23]。CD47通过其IgV结构域与整合素αvβ3发生相互作用,激活整合素,从而引发后续的胞内信号转导,而并不依赖其跨膜结构域[24]。CD47与这些整合素发生结合,相互作用,能够改变整合素信号靶点,如黏着斑激酶和桩蛋白[25],并且CD47-整合素复合体还与三聚体G蛋白相关,参与调控cAMP信号转导[26]。
CD47能够与血管内皮细胞生长因子(vascular endothelial growth factor,VEGF)以及 VEGFR2发生相互作用。而TSP1能够通过CD47抑制VEGF下游信号级联通路[27]。此外,免疫共沉淀和荧光共振能量转移表明,CD47能够与VEGFR2直接发生相互作用,不过这种相互作用会因TSP1和CD47结合以及VEGF与VEGFR2结合而被破坏,从而抑制VEGFR2信号转导过程[28]。
在红细胞中,CD47与Rh抗原复合体相互作用,形成复合体,这一复合体通过锚定蛋白与细胞骨架上的血影蛋白连接,有助于维持红细胞形态和氨的运输[29],不过CD47在其中的功能尚不清楚。
在T细胞中,Fas被发现与CD47发生横向相互作用。只有在表达CD47的细胞中,通过激活Fas才能够引起细胞凋亡。CD47与Fas的相互作用对于Fas下游的信号通路包括caspase-7和聚腺苷酸二磷酸核糖转移酶(poly ADP-ribose polymerase,PARP)裂解都是必要的[30]。
1.3.3 胞内相互作用 将CD47跨膜结构域和C末端尾巴作为酵母双杂交系统诱饵,从人类淋巴细胞cDNA文库中筛选,发现BNIP3是其胞浆结合配体[31]。反向抑制BNIP3能够抑制CD47介导的细胞凋亡,而用TSP1激活CD47能够诱导BNIP3转移到线粒体,导致细胞死亡,而SIRPα与CD47结合并不能引起BNIP3 转移[32]。
利用CD47胞浆尾巴的整合素相关蛋白-2(integrin associated protein-2,IAP2) 和 IAP4剪切异构体作为酵母双杂交系统诱饵鉴别出了另外2种泛素相关蛋白作为CD47胞浆结合配体,分别是连接整合素相关蛋白和细胞骨架蛋白PLIC1和PLIC2,后续研究发现PLIC1结合G蛋白βγ亚基(Gβγ),从而使CD47与三聚体G蛋白相结合,调节其下游信号通路[33]。
2 CD47相关信号通路
在胞外与CD47相互作用的配体分别是TSP1、SIRPα和CD47自身。其中,目前对CD47自身的同型相互作用参与的信号转导知之甚少。因此,目前CD47主要参与的信号通路可以按胞外与之相互作用的配体大致分为2个方面,即TSP1和SIRPα分别与CD47相互作用参与调控的信号通路,如图2所示。
2.1 CD47/TSP-1相关通路
2.1.1 调控G蛋白通路 CD47与整合素形成的复合体可通过其胞内配体PLIC1与三聚体G蛋白相互作用,形成CD47-整合素-G蛋白复合体[33]。CD47的5个跨膜结构域与其配体整合素的2个TM片段形成一个临时的7次跨膜复合体(7TMS),7TMS能够激活G蛋白。不同的整合素配体以及具有不同胞浆尾巴的CD47异构体都能够使7TMS复合体具有激活特异性三聚体G蛋白的功能[34]。G蛋白可以介导CD47多种细胞效应,如CD47相关的血小板激活需要G蛋白激活Syk激酶,从而诱导Lyn和FAK磷酸化[34]。CD47-整合素-G蛋白复合体的另一成分是胆固醇。去除胆固醇不会影响整合素亲和性,但会降低CD47刺激细胞黏附到玻连蛋白的能力。因此,虽然细胞基本的扩散机制与胆固醇无关,但是胆固醇似乎对保持功能性的CD47-整合素-G蛋白复合体起着必要的作用[35]。
2.1.2 调控NO/cGMP信号通路 在血管细胞中,内皮一氧化氮合酶(endothelial NO synthase,eNOs)是主要的一氧化氮(NO)合酶异构体,它能将L-精氨酸转变成L-瓜氨酸并释放NO。eNOs活性受到机械刺激和循环因子如VEGF和乙酰胆碱的调控。VEGF通过内皮细胞上的VEGFR2激活PI3K,后者激活蛋白激酶B(protein kinase B,PKB,又称为Akt),Akt磷酸化eNOs上的第1177位丝氨酸,使其激活[36]。同时活化的VEGFR2募集Src并诱导Ca2+信号通路,进一步激活eNOs[36]。产生的NO结合靶细胞中可溶性鸟苷酸环化酶(soluble guanylate cyclase,sGC)的血红素结构,增加其酶催化活性,导致胞内cGMP积聚,cGMP激活cGMP依赖性激酶 1(cGMP-dependent protein kinase,cGK1),后者磷酸化肌球蛋白轻链激酶并抑制其活性,使得肌球蛋白轻链脱磷酸化而引起平滑肌松弛[36]。TSP1能够与CD47结合,破坏CD47与VEGFR2复合体,抑制VEGFR2激活[28],进而影响下游信号通路。
2.1.3 调控细胞生存 TSP1与CD47结合能够诱导BNIP3转移到线粒体,BNIP3跨膜结构域插入到线粒体膜,引起线粒体通透性转换孔打开,释放细胞色素C,导致细胞死亡[32]。缺乏CD47有助于细胞通过激活保护性的自噬通路促进其生存[37]。自噬是一种维持内环境稳定的分解代谢机制,是胞质溶胶和细胞器被隔离到双层膜的小泡中,由此运送到溶酶体/空泡中降解,并因此形成的大分子进行再循环的一个过程。轻度的自噬可以防止细胞被不利因素损害,有助于其生存。缺乏CD47的细胞受到辐射时,beclin-1、ATG5、 ATG7表达升高,p62表达降低,LC3表达升高,自噬小体形成增多,激活保护性自噬。其中Beclin-1也能够与Bcl-2/Bcl-xL结合构成复合体,从而自噬的激活过程将会受到抑制,因此,Beclin-1与Bcl-2相互作用可直接控制自噬与凋亡[37]。
2.2 CD47/SIRPα相关通路
2.2.1 调控吞噬作用 CD47作为一种自体的识别分子,或者是作为一种“不要吃我”的信号分子,能够与巨噬细胞表面的SIRPα相互作用,防止肿瘤细胞和红细胞被清除[5,38]。这种防止巨噬细胞吞噬的机制涉及到SIRPα的抑制性信号通路,其中部分是通过SHP-1来抑制巨噬细胞吞噬作用[19]。
此外,有研究发现,CD47-SIRPα相互作用在红细胞清除方面的功能可能要更为复杂。人体红细胞的衰老似乎与CD47的构象改变有关,可能是依赖于氧化修饰,其能够将分子从“不要吃我”转变为“吃我”的结构[38]。因此,CD47不仅是一种“不要吃我”的信号,也可能是作为一种调节红细胞清除的开关。因此,CD47或者SIRPα表达异常或相互作用受到干扰都会改变巨噬细胞的吞噬作用,破坏红细胞的体内稳态。
2.2.2 调节树突状细胞和T细胞体内稳态 有研究发现,SIRPα变异小鼠的脾脏CD4+DC细胞数量减少,半衰期降低[39],脾脏中CD4+T细胞数量减少,T细胞聚集区域变小[40]。表明CD47与SIRPα相互作用对于调节树突状(DC)细胞和T细胞体内稳态具有重要的作用。此外有研究发现,DC细胞和T细胞应答在多种水平上受到CD47与SIRPα相互作用的影响,CD47与SIRPα相互作用受到干扰会使得DC细胞功能受损,尤其是在诱导辅助性T细胞Th1、Th2、Th17和自然杀伤性T细胞(natural killer T cell,NKT)应答方面[41-42]。
3 CD47相关信号通路与疾病的关系
3.1 癌症
CD47在多种肿瘤细胞表面高表达,其作为一种“不要吃我”的信号分子,与巨噬细胞表面SIRPα相互作用,从而使肿瘤细胞避免被吞噬清除,增强肿瘤生存能力[3-5]。因此,靶向CD47-SIRPα相互作用可能成为肿瘤治疗的一种新策略。
目前抗肿瘤治疗主要靶向CD47-SIRPα轴,多种治疗药物已处于临床研究阶段,包括常规抗体,融合蛋白和双特异性分子,分别阻断CD47或SIRPα(见表1)。
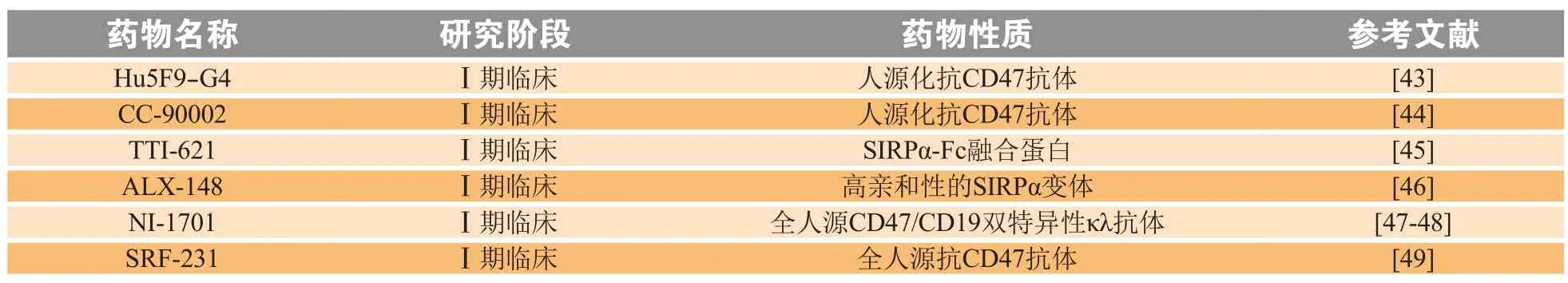
表1 靶向CD47-SIRPα的抗肿瘤药物Table 1 CD47-SIRPα targeting therapeutics for tumor
3.2 心血管疾病
心血管疾病主要的临床表现包括卒中、黄斑变性、外周血管病、高血压、心脏和其他关键器官衰竭、心脏病、伤口愈合不良等。其病因主要是急性或亚急性血流量下降并伴随组织低灌注,而血管细胞内NO信号通路受损是导致心血管疾病的关键因素[50]。在衰老过程中,内源性NO合成减少而TSP1表达升高。如前所述,TSP1和CD47相互作用能够抑制NO/cGMP信号通路,因此抑制TSP1-CD47相互作用能够改善NO信号通路,减轻心血管疾病的发生[28]。
Kojima等[51]采用CD47阻断抗体MIAP410,发现体外可诱导病变的、死亡的与即将死亡的平滑肌细胞和巨噬细胞被清除,并在几种不同的动脉粥样硬化小鼠中,利用抗CD47抗体阻断CD47,从而显著地阻止动脉斑块堆积,使病变血管组织的清除恢复正常。
3.3 自身免疫性疾病
有研究表明,SIRPα和CD47与Th1细胞或Th17细胞相关的自身免疫性疾病的形成有关,如实验性变态反应性脑脊髓炎(experimental autoimmune encephalomyelitis,EAE)、细菌或胶原诱导的关节炎、接触性超敏反应、结肠炎、克罗恩病等[52-53]。如前所述,CD47-SIRPα相互作用对DC细胞的功能具有调节作用,尤其是迁移和抗原呈递,其对于激活自身反应性Th1细胞和Th17细胞是必须的。在关节炎、结肠炎和克罗恩病中,干扰CD47-SIRPα相互作用能够抑制病情,然而在EAE中却会加重自身免疫和相关疾病[53]。研究表明,SIRPα缺失小鼠或者给予抗SIRPα抗体(P84)能够显著减轻Ⅱ型胶原型关节炎小鼠的炎症水平、骨侵蚀和关节炎症状[42,53]。
3.4 噬血细胞综合征
HLH典型特征之一就是噬血细胞作用增强,主要是由于细胞毒性T淋巴细胞(cytotoxic lymphocyte,CTL)和NK细胞清除病原功能受损,导致巨噬细胞过度增殖,噬血细胞增多。CD47在HLH的CD34+CD38-HSCs中表达下调,巨噬细胞对其吞噬作用增强,从而发生噬血细胞综合征[54]。
4 结语与展望
CD47与其2个配体TSP1和SIRPα相互作用,参与多种信号通路,参与机体多种生理功能及病理过程的发生发展,因此靶向CD47为疾病的治疗提供了一种新的方法。目前,靶向CD47的治疗主要还是集中在抗肿瘤领域,治疗策略主要有阻断抗体、重组融合蛋白、反义RNA、小分子抑制剂,其中阻断抗体和重组融合蛋白研究较多,主要是分别靶向CD47及其2个配体SIRPα和TSP1[55]。目前国际上处于I期临床的靶向CD47的抗肿瘤药物包括Hu5F9-G4、CC-90002、TTI-621、ALX148等, 其中TTI-621是可溶性重组SIRPα-Fc融合蛋白,其穿透能力和组织分布性较好,与红细胞的亲和力也更低,安全性更高[19]。国内走在前列的是宜明昂科生物的IMM01,目前已获得临床试验受理。小分子抑制剂,如多肽,体内半衰期较短,且容易酶解,难以发挥治疗作用,但其可用于设计有效稳定的模拟肽,如TSP-1的模拟肽,可用于识别其受体CD36[32]。反义RNA能够阻断CD47 mRNA翻译,从而抑制CD47蛋白表达,也有望成为靶向CD47的有效手段[56]。总之,CD47不仅是目前肿瘤领域研究的热点,在其他多种疾病中也扮演着重要作用,开发靶向CD47药物有望为疾病的治疗提供新的策略。
