韭菜无融合生殖种子形成机制研究进展
2020-06-23张华敏陈建华尹守恒马爱锄肖婉露焦子源
张华敏,陈建华,尹守恒,马爱锄,张 明,肖婉露,焦子源
(平顶山市农业科学院/河南省韭菜工程技术研究中心,河南 平顶山 467001)
无融合生殖(Apomixis)是一种通过种子进行无性繁殖的过程,其种子后代在遗传上是母本的克隆,是植物固定和利用杂种优势的理想途径,具有巨大的潜在利用价值[1]。目前已在52个科的400多种被子植物中发现了无融合生殖现象,其中75%属于禾本科、菊科和蔷薇科,但在主要农作物中未发现无融合生殖现象[2-3]。在发现的具有无融合生殖现象的植物中,大多数是多倍体[4]。葱属(Allium)是百合科(Liliaceae)葱族的重要类群,具有重要的经济价值,常作为蔬菜、调味品、药用和观赏植物。葱属在全世界约有850个种,在我国有140个种、13个变种和1个亚种[5],大多数种是二倍体和同源多倍体共存的[6-10]。由于前人对葱属植物种子生殖方式的研究很少,目前只在韭(A.tuberosumRottler ex Spr.)[11]、野韭(A.ramosumsyn.odorum)[12]和齿丝山韭(A.nutans)[13]3个种中发现了无融合生殖现象。由于葱属植物多倍体材料的广泛存在,在通过种子进行繁殖的多倍体材料中应该还有具无融合生殖能力的材料。
韭菜是百合科葱属中以鲜嫩叶及柔嫩假茎或肉质根为食的多年生宿根草本植物,是我国重要的特色蔬菜作物之一,在居民蔬菜消费中占有重要地位。在植物分类学上,我国栽培和食用的韭菜主要有韭、野韭和宽叶韭(A.hookeriThwaiter.Enum.)3个种[14]。韭和野韭属于葱属根茎组,染色体基数是8,大多数材料是同源四倍体[6],有少量二倍体、三倍体材料存在[15-16]。韭和野韭的亲缘关系很近,有学者认为野韭是韭的祖先[6,17],也有研究表明,韭和野韭是独立进化的2个种,在相当长一段时期没有基因交流[16]。宽叶韭属于葱属粗根组,有22条染色体,其中14条可以配成7对,8条不能配对,因此被认为是一个种间杂种,是双基数的同源异源三倍体[18]。宽叶韭花粉母细胞在减数分裂时染色体配对发生异常,导致不育,只能进行分株繁殖[19],少数染色体加倍为44条的材料育性得到一定的恢复,可结少量的种子[18]。本研究中的韭菜是指具有无融合生殖能力的同源四倍体韭和野韭。
生产中栽培的韭菜大多数是同源四倍体,不仅能进行分株繁殖,还可进行种子繁殖,种子结实正常而且发芽率很高,由于分株繁殖的繁殖系数低,商品生产的韭菜通常通过种子繁殖。韭菜是多倍体,再加上其无融合生殖特性,导致韭菜性状遗传极其复杂,不易分析。国内学者对韭菜无融合生殖特性的认识也存在误解[20-21]。韭菜育种没有按照其兼性无融合生殖特性设计育种路线,造成育种效率低、周期长,给韭菜品种改良带来困难。鉴于此,综述了韭菜无融合生殖种子的形成机制,有利于提高对韭菜无融合生殖特性的认识,为今后韭菜遗传和育种研究的开展提供新思路。
1 韭菜无融合生殖特性的发现
韭菜是同源四倍体作物,但在韭菜的遗传育种研究中,未发现同源四倍体植物的遗传特征[22]。同源四倍体植物有4个等位基因,在减数分裂时由于大小孢子母细胞同源染色体联会和分离紊乱导致部分雌雄配体生长发育异常,造成育性降低,结实率下降[23-24],但是韭菜结实率正常。在发育正常的雌雄配子体中,4个等位基因存在3种不同的分离方式,分别为按染色体随机分离、按染色单体随机分离和基因位点与着丝点间交换为50%时的染色单体分离[25]。有性生殖条件下,无论是自交还是回交,同源四倍体材料的纯合速率较二倍体慢得多,所需要的分离群体也要大得多,因此,同源四倍体育种难度很大,但是韭菜种子后代的表型大多与母本一致,性状变异少[22,26]。
韭菜无融合生殖现象的发现始于学者们对韭菜的胚胎学观察,HÅKANSSON等[12-13]观察到了韭菜胚的孤雌生殖现象,并观察到了野韭胚囊母细胞减数分裂时的核内复制现象,减数分裂后形成染色体未减数的雌配子。随后,有多位学者在韭菜的胚囊母细胞减数分裂中观察到了相同的现象[11,27]。KOJIMA等[28]在进行韭菜未授粉子房培养时获得了大量的四倍体植株,同样表明胚囊中形成的细胞染色体未减数。这些研究都表明韭菜具有无融合生殖能力。
2 韭菜的胚囊发育与花粉形成
2.1 韭菜胚囊母细胞减数分裂与胚囊发育
2.1.1 韭菜胚囊母细胞的减数分裂 韭菜胚囊母细胞减数分裂后大多数形成未减数胚囊,极少数形成减数胚囊。韭菜的未减数胚囊起源于大孢子母细胞,属于二倍体孢子生殖[27]。但是,韭菜未减数胚囊的形成与其他二倍体孢子生殖的植物不同。韭菜未减数胚囊的形成属于核内复制型减数分裂,加倍后的染色体在减数分裂的中期Ⅰ形成与体细胞数目相同的同源二价体,后期Ⅰ进行均等分离,最后发育成未减数胚囊。而在其他二倍体孢子生殖的植物中未减数胚囊类型有2种,一种是蝶须型(Antennaria type),大孢子母细胞直接进行有丝分裂形成胚囊,另一种是蒲公英型(Taraxacum type),大孢子母细胞在进入减数分裂的前期Ⅰ后,不发生同源染色体的联会,形成含有整套体细胞染色体的“重组核”,再进行有丝分裂,产生成熟胚囊[29]。
韭菜未减数胚囊的形成是由于大孢子母细胞中的染色体在减数分裂前的某个时期进行了核内复制加倍,然后再进行减数分裂,最终形成未减数胚囊。在韭菜大孢子母细胞减数分裂的中期Ⅰ,可明显观察到有32个二价体,后期Ⅰ进行32∶32的分离,最终形成未减数胚囊[11,27]。KOJIMA等[28]通过韭菜未授粉子房培养获得的韭菜植株中有98.5%是染色体数未减半的四倍体植株,也表明韭菜大孢子母细胞减数分裂形成的胚囊绝大多数都是未减数胚囊。HÅKANSSON等[12]将这种类型的减数分裂称为核内复制型减数分裂,将减数分裂过程中形成的二价体称为同源二价体(Autobivalent)。
韭菜减数胚囊的发育过程与未减数胚囊相似,不同的是减数分裂前染色体未加倍。在减数胚囊减数分裂的中期Ⅰ,可观察到同源染色体联会形成多价体、二价体和单价体,其中四价体最多[11,27],由于染色体联会的异常,在后期Ⅰ可能出现染色体不均等分离,形成非整倍体的雌配子。KOJIMA等[28]在韭菜未授粉子房培养植株中获得了染色体数为17的非整倍体植株。
2.1.2 韭菜胚囊发育 韭菜雌蕊由3个心皮构成三室子房,每室着生2个倒生胚珠。在胚珠发育的初期,珠心顶端表皮下的一个细胞体积增大,分化为孢原细胞并直接发育为大孢子母细胞[20]。大孢子母细胞经减数分裂,最终形成7胞8核的成熟胚囊。
韭菜的胚囊发育属于双孢子型中的葱型,与大葱的胚囊发育过程相似[30]。韭菜大孢子母细胞减数第1次分裂后形成垂直于珠心表皮排列的二分体,其中珠孔端的很快退化解体,合点端的继续进行减数第2次分裂,形成二核细胞并发育成2核胚囊,2个核分别移向胚囊的两端,并各进行一次有丝分裂形成4核胚囊,这4个核再进行一次有丝分裂,形成8核胚囊,胚囊的两端各有4个细胞核。在珠孔端的4个核中,靠近珠孔的1对与胚囊纵轴垂直的孪生核形成2个助细胞,而与胚囊纵轴平行的1对孪生核分别形成卵细胞与珠孔极核。在合点端的4个核中,1对孪生核形成2个反足细胞,另1对孪生核分别形成第3个反足细胞与合点极核,2个极核移向胚囊中央形成中央细胞[20,31]。韭菜反足细胞的发育比较特殊,3个反足细胞在形态上类似卵器,其中与合点极核同一来源的反足细胞的形态类似卵细胞,而与胚囊纵轴垂直的1对孪生核所形成的2个反足细胞在形态上类似助细胞[20,27]。
2.2 韭菜小孢子母细胞减数分裂与花粉粒形成
大多数同源四倍体植物的小孢子母细胞减数分裂异常,从而导致花粉的育性降低[32]。韭菜小孢子母细胞的减数分裂与其他同源四倍体相同,也存在分裂异常现象。在韭菜小孢子母细胞减数分裂的中期Ⅰ,染色体联会除形成四价体外,还观察到三价体、二价体和单价体。由于染色体的联会异常,在后期Ⅰ,染色体不均等分裂,可观察到染色体桥、染色体断片和染色体滞后现象,发育成熟的花粉粒中通常有10~19条染色体[11,27,33-34]。另外,韭菜小孢子母细胞中也存在染色体核内复制加倍现象,与大孢子母细胞相同,在减数分裂的中期Ⅰ形成32个二价体,在后期Ⅰ,染色体均等分裂,最后发育成染色体数为32的未减数花粉粒[27,34-35]。韭菜的未减数花粉比未减数胚囊少,在四倍体材料Tender-Pole中有3.0%~6.0%[27],在二倍体材料KaD2中约有9.0%[36]。
韭菜的单花通常有6枚雄蕊,每个花药有4个药室。当韭菜花蕾直径约1.0 mm时,花药开始分化。在横切面上,花药为一团近似棱形的分生组织细胞,在4个角隅的表皮下,分别有4行较大的细胞,为孢原细胞。孢原细胞进行平周分裂,外侧形成初生壁细胞,内侧形成初生造孢细胞。中层由1层扁平细胞构成,细胞和核均切向引长;绒毡层细胞大,近方形,有一个巨大的球形细胞核。当花蕾直径约1.8 mm时,次生造孢细胞发育成小孢子母细胞并进入减数分裂,至花蕾直径约2.0 mm时,小孢子四分体形成。小孢子母细胞减数分裂的胞质分裂为连续型。药壁中层细胞在小孢子母细胞减数分裂前期Ⅰ时开始解体,到四分体时期全部消失。韭菜花药的绒毡层为分泌型绒毡层,在小孢子四分体时期体积达到最大,其内有较大的液泡,在小孢子早期,绒毡层开始退化,到二胞花粉早期,绒毡层完全消失。韭菜的成熟花粉粒为二细胞型。刚从四分体中释放的小孢子是单核花粉,单核花粉进一步发育,进行一次不等分裂,形成二胞花粉,其中生殖细胞长梭形,在核的周围可明显观察到1层染色较淡的细胞质鞘,而营养细胞核近球形。最后花粉发育成近似等腰三角形,有3个萌发孔[31,37]。除正常花粉粒外,还有少量的异型花粉,约有87%的花粉粒可被醋酸洋红或I/KI溶液染色[31,33]。
3 韭菜胚胎发育和种子形成
3.1 韭菜胚胎发育过程
韭菜的卵细胞、反足细胞和珠被细胞都可以分裂形成原胚,但在成熟种子中一般只有一个胚,只有极少量的多胚种子形成[20,38-39]。韭菜的反足胚一般在开花后8 d,随着胚乳的发育而发生退化,由于胚胎观察数量有限,研究者未直接观察到发育成熟的反足胚,但是韭菜多胚种子的存在表明除卵胚外,反足胚和珠被细胞发育形成的不定胚也有发育为成熟胚的可能。
韭菜的胚胎发育属于柳叶菜型。一般在开花后2 d,大多数卵胚开始发育,首先珠孔端的卵细胞进行一次横分裂,形成朝合点端的顶细胞和朝珠孔端的基细胞。接着基细胞横分裂,顶细胞纵分裂,形成“T”型四胞原胚。在基细胞横分裂的2个细胞中,珠孔端的细胞再次横分裂构成2个细胞胚柄,合点端的细胞则纵分裂产生2个细胞参与胚体的构成,以后不再分裂。而顶细胞纵分裂的2个细胞再次纵分裂,形成四细胞层,然后再向各方向分裂构成胚体的主要部分,最后发育成成熟卵胚。卵状反足细胞早期胚胎发育与卵胚相似,但是大多数反足原胚只能分裂数次就逐渐退化[20,38]。
3.2 授粉在韭菜种子形成过程中的作用
韭菜无融合生殖种子的形成需要未减数胚囊发育、卵细胞的孤雌生殖和极核的假受精(Pseudogamy)共同作用才能完成。因此,韭菜种子的形成需要授粉,但是胚发育不需要受精,由卵细胞孤雌生殖形成,胚乳的发育则依赖于极核假受精。韭菜同一朵花中的雌雄蕊发育不同步,雄蕊先熟,在开花当天花药即可开裂散粉,而雌蕊成熟一般在开花后第3天,此时柱头呈球形把手状,能使花粉萌发。在未授粉条件下,胚乳不能正常发育,导致卵胚和反足胚退化,授粉后,胚乳正常发育,卵胚才能继续发育,但是反足胚仍然退化[38]。一般到开花后第4天,柱头授粉后约16 h,花粉管才到达胚珠,释放精细胞进入胚囊,此时,大多数卵细胞已启动孤雌生殖并发育到四胞原胚阶段,因此,在授粉条件下,卵胚仍然是卵细胞孤雌生殖发育形成的[38]。
3.3 韭菜多胚与多胚苗现象
在被子植物的1个胚珠中包含2个或2个以上胚的现象被称为多胚现象。一粒多胚种子同时萌发出2株或2株以上的幼苗,这种一籽多苗被称为多胚苗[40]。胡适宜[41]最先报道了韭菜中存在多胚苗现象,有许多研究结果表明被子植物中的多胚现象与无融合生殖现象有一定的正相关性,因此有学者认为韭菜的多胚苗现象也与无融合生殖现象有关,并进行了进一步研究。王巨媛等[39]研究了我国4个生态区38个韭菜品种的多胚苗现象,发现韭菜多胚苗的自然发生频率为0~4.333%,且大多数是一籽双苗,极少数是一籽三苗,并将一籽双苗的多胚苗分为簇生双苗、中生双苗和对生双苗3类。韭菜的多胚苗性状可遗传,多胚苗发生频率高的材料后代多胚苗发生频率也高,并且化学诱导可提高韭菜多胚苗发生频率[21,42],但是未见韭菜多胚苗胚胎发生机制和多胚苗后代倍性鉴定的后续研究。KOJIMA等[27,38]对韭菜兼性无融合生殖特性的细胞学和胚胎学机制进行了阐述,表明韭菜绝大多数的种子都是无融合生殖形成的。因此,韭菜的多胚苗现象可能与无融合生殖不相关。推测韭菜多胚苗产生的原因有3个:一是韭菜的卵细胞一般情况下都发育成单胚,但也观察到极少数双胚或三胚现象存在;二是韭菜大多数的反足胚虽然都发生退化,但是也不排除极少数反足胚发育成熟的可能;三是韭菜极少数的珠被细胞也能发育成胚胎形成珠被胚[38]。
4 韭菜无融合生殖性状的遗传
4.1 韭菜二倍体无融合生殖和有性生殖材料的鉴定
具有无融合生殖特性的植物几乎都是多倍体,天然无融合生殖植物中二倍体材料极少。多倍体植物的基因组高度杂合且遗传背景复杂,因此无融合生殖性状的遗传分析相当困难。韭菜属于兼性二倍体孢子型无融合生殖,几乎所有的韭菜资源都是同源四倍体且无融合生殖率很高(>90%)[27,43],不能直接用于无融合生殖性状的遗传分析,获得二倍体无融合生殖和有性生殖材料是进行韭菜无融合生殖性状遗传分析的前提。韭菜中有少量的减数胚囊形成,这些减数胚囊中的卵细胞孤雌生殖能够形成染色体数减半的双单倍体(Dihaploid)后代,为获得二倍体无融合生殖和有性生殖材料提供了可能。KOJIMA等[28]在韭菜未授粉子房培养苗中鉴定出了1株双单倍体植株,并在1 016株韭菜的自交和杂交后代中鉴定出5株双单倍体植株[43],从这些韭菜双单倍体材料中鉴定出了1个无融合生殖率为96%的材料KaD2,其二倍体孢子率和卵细胞孤雌生殖率分别是96%和98%[36],随后又从蒙古的地方韭菜品种中鉴定出了3个有性生殖的二倍体韭菜材料94Mo13、94Mo49和94Mo50[44]。
4.2 控制韭菜无融合生殖性状的2个位点
研究表明,韭菜未减数胚囊的发育和卵细胞的孤雌生殖是2个相互独立的事件[27,38,45-46],受2个位点控制,与极核的假受精共同作用形成无融合生殖种子。因此,KOJIMA等[36]用二倍体孢子率、孤雌生殖率和无融合生殖率3个指标分析韭菜的无融合生殖性状(表1)。天谷正行等[47]找到了2个与韭菜孤雌生殖基因连锁的SCAR标记PLM1和PLM3,它们与孤雌生殖基因的遗传距离分别是3.4、1.1 cM。YAMASHITA等[46]利用韭菜二倍体有性生殖材料94Mo13、94Mo49和94Mo50和无融合生殖材料KaD2构建的F1群体和BC1群体,对韭菜的无融合生殖性状进行遗传分析得出3个结果。一是无融合生殖性状不能通过单倍体的花粉遗传给后代。在F1群体包含的110株二倍体和773株三倍体植株中,所有的二倍体植株都是有性生殖的,所有的三倍体植株都是无融合生殖的。二是与二倍体花粉相比,单倍体花粉的活力很差。KOJIMA等[36]观察到KaD2的花粉母细胞发生核内复制形成未减数花粉的比例只有9%,但是F1群体中三倍体植株与二倍体植株的比例为7∶1。作者认为引起单倍体花粉活性差的原因一方面是无融合基因与一些致死因子相关,另一方面是杂合态的韭菜在长期的无融合生殖条件下积累了很多有害的隐性突变。三是韭菜二倍体孢子基因和孤雌生殖基因由在遗传上相互独立的2个显性单基因D和P控制。在52株二倍体孢子生殖的BC1群体中,有28株是孤雌生殖的,其余24株是有性生殖的。同样,在49株有性生殖的BC1群体中,有24株形成二倍体孢子,其余25株的减数分裂正常。
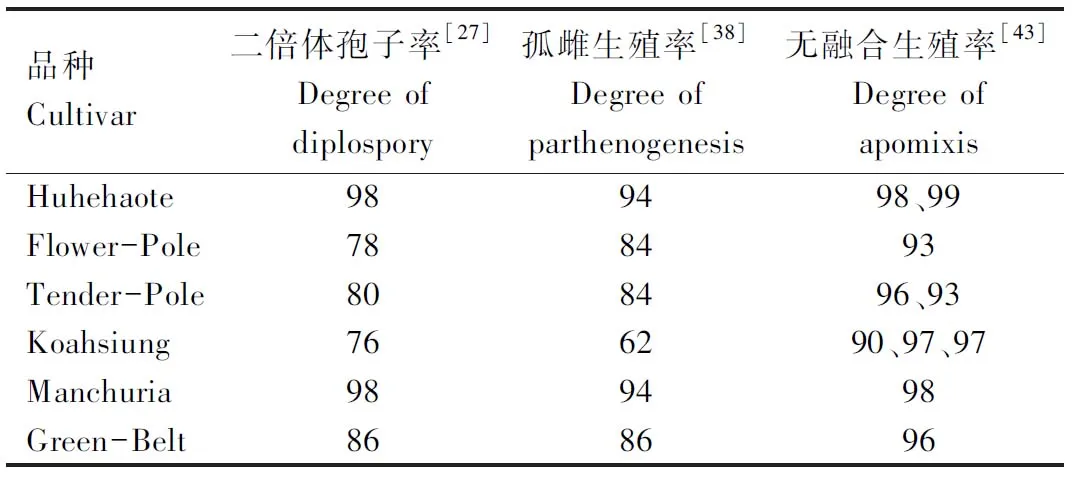
表1 6个韭菜品种的二倍体孢子率、孤雌生殖率和无融合生殖率Tab.1 Degrees of diplospory,parthenogenesis and apomixis in six Chinese chive cultivars %
5 展望
综上所述,韭菜的无融合生殖属于兼性二倍体孢子生殖类型。韭菜大孢子母细胞中的染色体在减数分裂前的某个时期进行了核内复制加倍,然后再进行减数分裂形成染色体未减数的胚囊,胚囊中的卵细胞具有孤雌生殖能力,可不经受精直接发育成胚,但是极核不能自主发育,需要受精才能发育成胚乳,最后形成无融合生殖种子。大多数韭菜材料的无融合生殖率在90%以上,也就是说韭菜仍然保留了少量的有性生殖能力,这些有性生殖后代是父母本雌雄配子结合后发育形成的。对植物育种来说,韭菜的兼性无融合生殖特性是一种理想的生殖类型,符合育种者对无融合生殖特性在育种中应用的要求,超过90%的无融合生殖率保证了后代在遗传上的相对一致性,少量的有性生殖后代又为种质资源创新和新品种的选育提供了可能。
韭菜是目前唯一报道的天然具有无融合生殖能力的栽培作物。但是,与山柳菊、蒲公英等其他无融合生殖植物相比,韭菜的无融合生殖特性研究很少,仅有日本和印度的少数学者对其进行了初步研究。我国也有少数学者关注到韭菜的无融合生殖特性,但是存在误解,认为韭菜的反足胚是无融合生殖胚,而卵胚仍然是有性生殖胚[20-21]。由于国内学者,特别是韭菜育种者对韭菜无融合生殖特性认识不足,仍然按照二倍体作物的育种路线进行育种,因此韭菜遗传育种研究进展缓慢[22]。深入研究韭菜的兼性无融合生殖特性不仅对韭菜的遗传育种研究有重要意义,也可为其他植物无融合生殖特性的研究和利用提供参考。今后的韭菜无融合生殖特性的研究应从以下几个方面开展:一是研究韭菜有性生殖后代的快速鉴定技术,加快韭菜的育种进程;二是研究现有韭菜种质资源的无融合生殖率,从中筛选无融合生殖率低的材料用于韭菜种质资源创新;三是加强韭菜无融合生殖机制的研究,为韭菜无融合生殖特性在育种中的应用提供理论基础;四是根据韭菜的兼性无融合生殖特性,设计适宜于韭菜的育种路线。
植物无融合生殖所形成的种子包含母本的全部遗传信息,它所产生的后代是母本的复制,能够保持母本所有的优良性状,这一特性对于杂交种的种子生产具有重大的意义,能在保持杂种优势的条件下大幅降低种子生产成本。因此,植物无融合生殖特性的研究受到学者们的极大关注。目前生产中应用的主要农作物中未发现有无融合生殖现象,在主要农作物中创造无融合生殖材料并应用于杂交种的种子生产是很多学者的梦想。研究者曾试图将调控无融合生殖的染色体片段导入近缘的栽培系中,但未获得成功[48]。最近,有研究者采用多重CRISPR-Cas9基因编辑技术,定点突变了水稻中的3个减数分裂相关基因PAIR1、REC8、OSD1和1个诱导孤雌生殖的基因MTL,成功获得了具有无融合生殖现象的Fix (Fixation of hybrids)水稻材料[49]。不论是从天然无融合生殖植物中导入,还是人工突变有性生殖相关基因,创造新的无融合生殖材料,都是未来利用植物的无融合生殖特性进行“一系法”杂交种生产的关键环节。
