猪圆环病毒2b型表位嵌合病毒样颗粒在E.coli中的制备及鉴定
2020-06-19刘晴坤任春晓刘运超张改平
冯 华,刘晴坤,2,任春晓,2,刘运超,张改平,2
(1.河南省农业科学院 动物免疫学重点实验室/河南省动物免疫学重点实验室/农业部动物免疫学重点实验室,河南 郑州 450002;2.河南农业大学 生命科学学院,河南 郑州450002)
猪圆环病毒2型(Porcine circovirus type 2,PCV2)属于圆环病毒科(Circovirus)、圆环病毒属(Circoviridae),是一类无囊膜的、直径在17~22 nm的单链环状DNA病毒[1-2],该病毒是引起一系列重要猪圆环病毒相关疾病(Porcine circovirus disease,PCVD)的主要病原[3-5]。其中,PCV2系统性疾病(PCV2-systemic disease,PCV2-SD)是主要由PCV2b病毒引起的一种系统性疾病,主要表现为生长缓慢、呼吸困难、淋巴结肿大、腹泻等。该病主要影响7~16周龄的仔猪,能够造成4%~30%的发病率和4%~20%死亡率,从而使猪场大量减产,导致巨大的经济损失[6]。
PCV2基因组由大小约为1.7 kb的双义DNA组成,包含2个主要的开放阅读框(Open reading frame,ORF):ORF1(904 bp)和ORF2(702 bp)[7],分别编码病毒复制相关蛋白Rep(Rep′)和病毒结构蛋白Cap。其中,Cap蛋白是PCV2病毒的主要功能性蛋白,主要行使细胞受体识别、核酸细胞核运送等功能[8]。同时,Cap蛋白也是PCV2主要的免疫原蛋白,包含重要的病毒中和表位,在体外能够自组装成病毒样颗粒(VLP),常作为血清学检测和疫苗研发的靶标蛋白[9]。大量研究表明,在PCV2 Cap蛋白C端插入外源表位能够同时诱导针对Cap和外源表位的免疫反应[10-11]。因此,在Cap蛋白C端插入其自身的中和表位将有助于进一步提高现有疫苗的保护效率。目前,Cap 蛋白上共鉴定出至少6个表位区域[12-14],中和表位226LKDPPLNP233被证实具有较好的中和免疫原性,能够诱导体液免疫反应,并参与Cap蛋白构象表位的形成,在亚单位疫苗的研发中具有很好的前景[15-16]。
目前,市场上针对PCV2的疫苗仍然以灭活疫苗为主。相比灭活疫苗,Cap亚单位疫苗免疫原性单一,可以避免潜在的由于灭活不完全造成的免疫动物携带疫苗毒的可能,在未来PCV2清除中将会具有更强的优势。为进一步提高现有亚单位疫苗免疫效率,以目前流行的PCV2b为研究对象,将其自身中和表位226—233位氨基酸序列连接到Cap蛋白C末端,成功构建连接自身中和表位的嵌合Cap蛋白,并对其免疫原性和结构特点进行初步鉴定,以期为研发更为安全、有效、廉价的PCV2疫苗提供新的思路。
1 材料和方法
1.1 载体、菌种及主要试剂
重组载体PE-Sumo载体由本实验室保存;大肠杆菌克隆感受态细胞JM109和表达用感受态细胞BL21(DE3)、DNA Marker DL2000、Primer STAR Max DNA 聚合酶、T4 DNA连接酶等均购自Takara公司;限制性内切酶BsaⅠ和XbaⅠ购自NEB 公司;核酸胶回收试剂盒、质粒提取试剂盒等均购自OMEGA公司;异丙基硫代半乳糖苷(IPTG)购自INALCO 公司;Ni-NTA亲和层析柱购自QIAGEN 公司;蛋白质Marker、Sumo蛋白酶购自Smobio公司;聚偏二氟乙烯(PVDF)膜和ECL 显色试剂盒购自索莱宝公司;抗组氨酸标签单克隆抗体、HRP标记羊抗鼠二抗购自Abcam公司;鼠源PCV2 Cap单克隆抗体由本实验室制备保存。
1.2 重组表达质粒的制备和重组表达菌体的构建
利用已经针对大肠杆菌表达系统进行密码子优化的PCV2b(GenBank:AAY969004.1)Cap序列为模板,分别以Capc-F:ATGGTCTCTAGGTATGACGTACCCTCGT和Capc-R:AGTCTAGATTACGGGTTCAGCGGCGGGTCTTTCAGGGGGTTCAAAGGTGGG-T(下划线代表中和表位位置)为上、下游引物进行扩增,将PCV2 Cap中和表位基因构建在Cap基因C端(Capc);目的片段经过限制性内切酶BsaⅠ和XbaⅠ酶切后,利用T4连接酶将其连接在PE-Sumo载体,并转化到大肠杆菌克隆感受态细胞JM109,挑选阳性质粒进行PCR鉴定,并将其送至上海生物工程有限公司进行测序鉴定,将构建成功的阳性重组质粒命名为PE-Sumo-Capc。
1.3 重组蛋白表达条件的优化和可溶性分析
将PE-Sumo-Capc转化到大肠杆菌BL21(DE3)感受态细胞中,并涂布于含有Amp+的LB固体培养基上37 ℃培养过夜;挑取单克隆接种于含0.1% Amp+的LB液体培养基中培养过夜后,以1∶100比例将其接种至新鲜的Amp+LB液体培养基中,37 ℃培养至OD600为0.4~0.6 时,加入终浓度为1 mmol/L的IPTG,分别在18、22、26、30 ℃条件下诱导表达10~12 h,优化表达时间。之后,按照上述方法在最适表达温度下,分别表达8、10、12、14、16 h,探索最适表达时间。将收获的菌体分别重悬于缓冲液(50 mmol/L Tris,250 mmol/L NaCl,pH值为8.0)中,超声破碎菌体分离上清、沉淀,并通过SDS-PAGE凝胶电泳对蛋白质的最适表达条件及其可溶性进行鉴定。
1.4 重组蛋白的纯化及免疫反应性鉴定
收集在优化条件下表达的菌体,重悬于裂解液中,按照上述方法超声破碎,收集上清。以缓冲液(50 mmol/L Tris,250 mmol/L NaCl,pH值为8.0)平衡Ni-NTA亲和层析柱后,将上清置于层析柱中使目的蛋白与柱子充分结合;再以含有终浓度为50 mmol/L咪唑的缓冲液充分清洗层析柱;最后,以含有终浓度为200 mmol/L咪唑的缓冲液对Sumo-Capc蛋白进行洗脱;之后通过SDS-PAGE和Western blot(抗组氨酸标签单克隆抗体为一抗)对目的蛋白进行鉴定。洗脱蛋白经无咪唑缓冲液透析去除咪唑后,按照Sumo蛋白酶说明书对纯化蛋白进行酶切以去除Sumo标签;之后,利用SDS-PAGE对酶切后的目的蛋白进行鉴定,并以鼠源PCV2 Cap单克隆抗体为一抗和HRP标记羊抗鼠为二抗对目的蛋白的免疫反应性进行Western blot鉴定。
1.5 PCV2 VLP动态光散射鉴定及电镜观察
对纯化后的Capc蛋白进行物理特性鉴定,利用动态光散射对纯化后的Capc蛋白的水化半径进行分析。另外,取微量纯化后的Capc蛋白,将其缓慢滴于铜网上,室温静置1 min后用滤纸从液滴边缘将多余液体吸取;同样取微量pH值为7.0的磷钨酸染液,滴于膜上染色约1 min,滤纸移去多余染液,干燥后透射电镜下观察。
2 结果与分析
2.1 重组表达载体PE-Sumo-Capc构建及鉴定
将重组Capc片段插入PE-Sumo载体中BsaⅠ和XbaⅠ酶切位点之间,并转化JM109感受态细胞,经过培养,利用引物Capc-F和Capc-R对挑取的克隆进行PCR鉴定。2%的琼脂糖凝胶电泳结果显示,在约747 bp处出现明显条带,符合预期大小(图1)。将鉴定为阳性的克隆送上海生工生物工程有限公司进行测序鉴定,结果正确。
2.2 重组Sumo-Capc蛋白表达条件的优化和可溶性分析
将重组表达质粒PE-Sumo-Capc转化到大肠杆菌BL21(DE3)感受态细胞后,对重组蛋白Sumo-Capc的最适表达温度和时间进行优化。收获的菌体经超声破碎分离上清及沉淀后,SDS-PAGE鉴定结果显示,在约48 ku处有明显的条带出现,与预期的重组蛋白Sumo-Capc大小相符,证明目的蛋白表达量较高;经过优化,目的蛋白在26 ℃时能够以可溶形式表达于上清中,并且表达量相比其他温度较高,大肠杆菌本体杂蛋白表达量较低;时间优化结果显示,在26 ℃下表达12 h后,目的蛋白表达量不再增加(图2、3)。因此,重组Sumo-Capc蛋白最适表达条件确定为26 ℃诱导12 h。
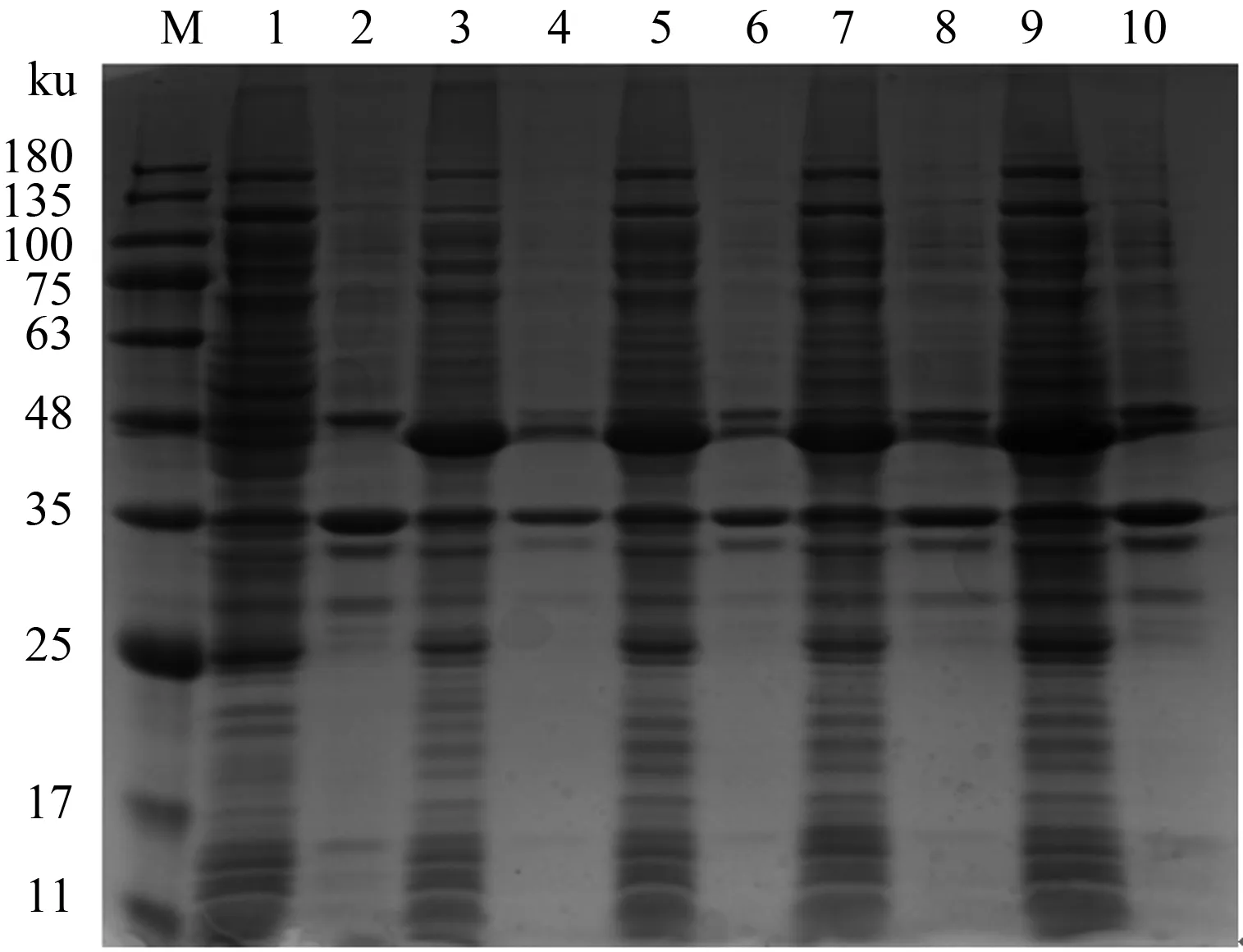
M:蛋白质Marker;1、2分别为诱导8 h后的上清和沉淀;3、4分别为诱导10 h后的上清和沉淀;5、6分别为诱导12 h后的上清和沉淀;7、8分别为诱导14 h后的上清和沉淀;9、10分别为诱导16 h后的上清和沉淀M:Protein Marker;1,2 represent supernatant and precipitate induced at 8 h,respectively;3,4 represent supernatant and precipitate induced at 10 h,respectively;5,6 represent supernatant and precipitate induced at 12 h,respectively;7,8 represent supernatant and precipitate induced at 14 h,respectively;9,10 represent supernatant and precipitate induced at 16 h,respectively图3 26 ℃下重组Sumo-Capc蛋白表达时间优化结果Fig.3 Results of expression time optimization of recombinant Sumo-Capc protein at 26 ℃
2.3 重组Sumo-Capc蛋白的纯化、酶切和Western blot鉴定
SDS-PAGE和Western blot(抗组氨酸标签单克隆抗体为一抗)结果显示,超声破碎后上清经过Ni-NTA亲和层析柱纯化后,能够得到较纯的带有Sumo标签的目的蛋白(图4)。以Sumo蛋白酶对Sumo-Capc蛋白进行酶切去标签,SDS-PAGE结果显示,在约27 ku处显示明显条带,与预期的Capc蛋白大小一致(图5A);Western blot结果显示,该蛋白质能够与鼠源PCV2 Cap单克隆抗体产生反应,证明纯化后的Capc蛋白具有针对PCV2 Cap单克隆抗体的免疫反应性(图5B)。
2.4 PCV2病毒样颗粒物理特性鉴定
从图6可看出,纯化后的Capc蛋白形成了均一的、直径为17~19 nm的蛋白质聚集体。进一步透射电镜下观察发现,纯化后的Capc蛋白能够自组装形成直径为17~19 nm的病毒样颗粒(图7)。
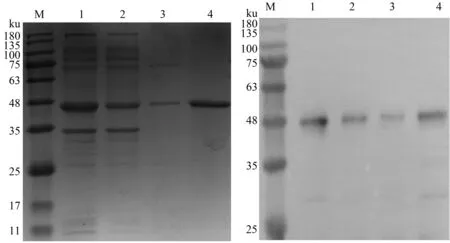
M:蛋白质Marker;1:裂解液上清;2:上样流穿液;3:洗杂液;4:纯化的Sumo-Capc蛋白M:Protein Marker;1:Supernatant after sonication;2:Flow fluid after sample loading;3:Wash flow fluid;4:Purified Sumo-Capc protein图4 纯化重组Sumo-Capc蛋白的SDS-PAGE和Western blot分析Fig.4 SDS-PAGE and Western blot analysis of purified recombinant Sumo-Capc protein
3 结论与讨论
PCV2是引起一系列包括PCV2系统性疾病、PCV2亚临床感染、呼吸性疾病、猪皮炎/肾病综合征等猪圆环病毒相关疾病(PCVD)的主要病原体,对世界范围内生猪养殖业造成了巨大威胁[17]。此外,该病毒具有变异快、传播广、毒力强、难清除等特点,对我国养猪业造成了巨大的经济损失,已成为制约我国生猪产业发展的重要因素之一[7,18]。
疫苗免疫是预防PCVD的主要手段,免疫后能够有效控制PCV2感染和PCVD相关疾病暴发[19]。目前,市场上针对PCV2的疫苗多为灭活疫苗和亚单位疫苗,并且大多数是基于PCV2a型毒株研发[19]。GenBank数据显示,PCV2a型毒株在2003年以前是全球主要流行毒株,之后主要流行毒株变成了PCV2b型[20],而PCV2b毒株感染也成为我国养猪业面临的重要问题。与现有PCV2a的疫苗相比,基于PCV2b研发的疫苗能够为动物提供更好的保护[21]。另外,由于PCV2当前并不能完全从猪场清除[19],针对当前流行的毒株不断提高现有疫苗效力成为PCV2疫苗研发的主要方向。因此,本试验以研究基于PCV2b亚型的亚单位疫苗为目的,以期为控制PCV2感染提供更为高效的疫苗候选。
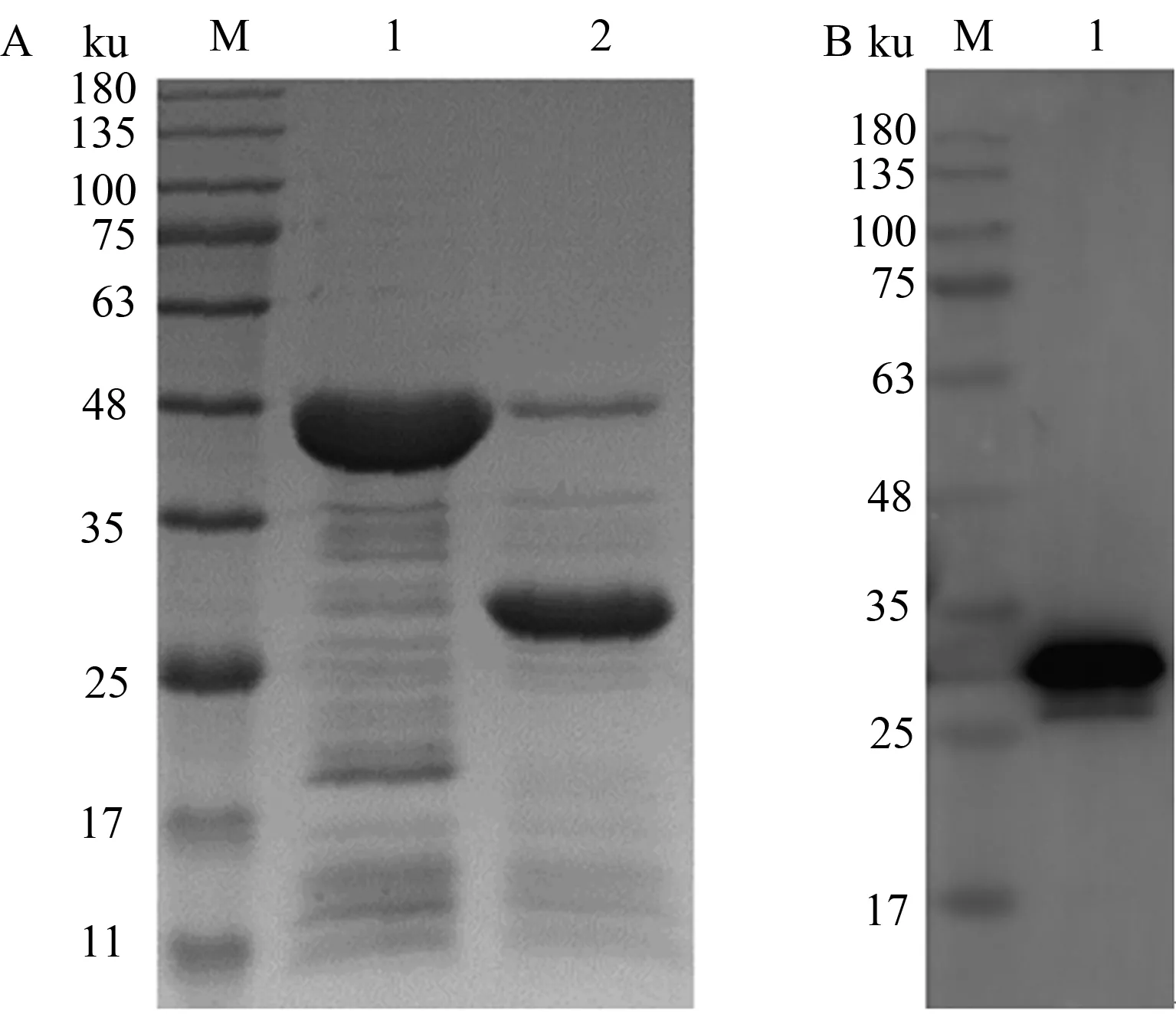
A.Sumo-Capc蛋白的酶切结果(M:蛋白质Marker;1:纯化的Sumo-Capc;2:酶切后的Capc);B.纯化Capc蛋白的 Western blot 鉴定结果(M:蛋白质Marker;1:纯化的Capc)A.Enzyme digestion of Sumo-Capc(M:Protein Marker;1:Purified Sumo-Capc;2:Capc after enzyme digestion);B.Western blot analysis of purified Capc (M:Protein Marker;1:Purified Capc)图5 Sumo-Capc蛋白的酶切和纯化Capc蛋白鉴定Fig.5 Enzyme digestion of Sumo-Capc protein and identification of purified Capc protein
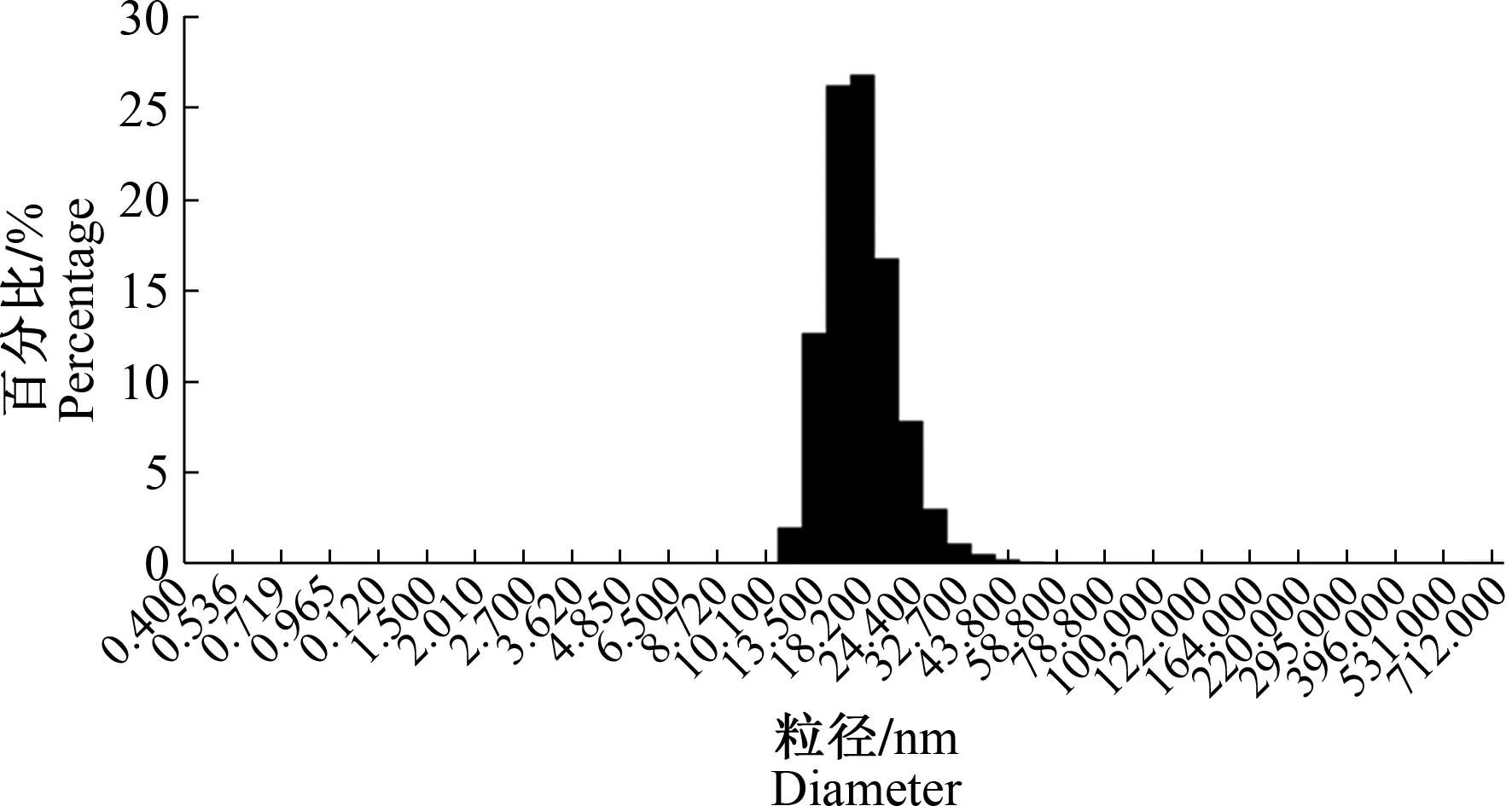
图6 重组Capc蛋白的动态光散射分析Fig.6 Dynamic light scattering analysis of recombinant Capc protein
病毒样颗粒具有与天然病毒形态完全一致的结构,由于缺乏DNA或RNA等遗传物质不具有感染性,并能够同时激活B细胞介导的体液免疫和细胞免疫反应,有效保护靶动物[22-23]。本研究以PCV2b型毒株Cap基因序列为对象,针对大肠杆菌表达系统进行密码子优化,通过PCR将PCV2病毒中和表位226LKDPPLNP233构建于Cap基因C端,将重组载体转化表达菌后,通过诱导表达、纯化,成功得到连接有自身中和表位的嵌合PCV2b病毒样颗粒。重组蛋白Sumo-Capc的最适表达条件为26 ℃下诱导 12 h,该条件下目的蛋白容易富集并且具有较少的背景蛋白。Western blot鉴定结果显示,纯化后的目的蛋白Capc能够与PCV2单克隆抗体产生特异的反应,初步证明该蛋白质具有针对PCV2 Cap蛋白的免疫反应性。动态光散射和透射电镜分析结果显示,目的蛋白形成了直径为17~19 nm的病毒样颗粒,与前人研究结果相符[24-25]。可见,在Cap蛋白C端插入同源中和表位片段对Cap病毒样颗粒的形成能力没有造成影响。前人研究也发现,插入合适长度的异源表位对PCV2病毒样颗粒的形成无影响[22]。本试验得到的病毒样颗粒粒径并不均一,可能与插入表位的长度和理化特性有关。因而,Cap蛋白C端连接不同长度或具有不同理化特性的插入片段对PCV2病毒样颗粒形成大小的影响需要进一步研究。

图7 电镜观察Capc蛋白自组装形成的病毒样颗粒 Fig.7 Transmission electron micrograph of viruslike particles by Capc protein self-assembly
综上,本试验成功表达了串联PCV2b中和表位的嵌合Capc蛋白,该蛋白质能够与PCV2单克隆抗体产生特异的免疫反应性,并能够形成病毒样颗粒,为后续的动物试验奠定了基础,为PCV2相关疫苗的研究提供新的思路。
