泽兰实蝇寄生对四川地区紫茎泽兰种子质量的影响
2020-06-07谌利民曾春艳
王 勤,谌利民,曾春艳,曹 阳,陈 瑶,陈 艳*
(1.绵阳师范学院生态安全与保护四川省重点实验室、生命科学与技术学院,四川绵阳 621006;2.四川省唐家河国家级自然保护区管理处,四川广元 628100)
0 引言
紫茎泽兰(AgeratinaadenophoraSprengel)为菊科泽兰属多年生丛生型半灌木草本植物,是世界性恶性入侵植物之一[1-2].紫茎泽兰原产于墨西哥,约在20世纪40年代由缅甸传入我国云南省,现已广泛分布在我国西南地区的云南、贵州、四川与广西等省.一旦在新的环境中定植成功,紫茎泽兰可迅速形成单优群落,给当地的农林牧业生产造成巨大损失,并对生物多样性和生态环境带来严重的影响[1, 3].紫茎泽兰可以通过根状茎或种子进行繁殖,种子繁殖在其扩散中起到重要作用.紫茎泽兰的种子小又轻,且具冠毛,一株成熟的紫茎泽兰植株每年约产生1万~10万粒种子[4],可随风媒、水媒、动物和车载传播,其中水媒和车载是其进行远距离传播的主要方式[5].
泽兰实蝇(ProcecidocharesutilisStone)是紫茎泽兰的专食性寄生天敌,其幼虫蛀入紫茎泽兰幼嫩的茎枝端部,诱导紫茎泽兰形成膨大虫瘿,在一些地区对紫茎泽兰的生长和繁殖起到了抑制作用[6].自20世纪20年代以来,泽兰实蝇被许多国家用于防治紫茎泽兰,1945年从墨西哥引进到夏威夷,再依次引进到澳大利亚、新西兰与印度等国家的各地区,但只在夏威夷得到了较好的防控效果[2, 7].我国于1984年从西藏将泽兰实蝇引入到云南的部分地区,随后引入到西南地区的四川、贵州等地并形成自然种群[8],但对紫茎泽兰并没有产生理想的控制效果.有研究发现,泽兰实蝇的寄生可以降低紫茎泽兰的种子结实量、种子千粒重与种子活力等[9],还降低了紫茎泽兰种子的萌发率和发芽能力,起到控制紫茎泽兰的蔓延和扩散的作用[10];但也有研究发现泽兰实蝇的寄生对紫茎泽兰的生长和繁殖无明显的抑制作用[11-12].那么在四川地区,泽兰实蝇的寄生对紫茎泽兰的种子质量有怎样的影响呢?我们从被泽兰实蝇寄生和未被寄生的紫茎泽兰枝条上收集成熟种子,比较种子千粒重及种子的萌发特性,旨在探讨泽兰实蝇对紫茎泽兰繁殖的影响,以期为紫茎泽兰的防治提供参考.
1 材料与方法
1.1 种子采集
供试种子于2019年4月下旬采集于汉源县、冕宁县、西昌市和平地镇(攀枝花市仁和区)4个紫茎泽兰自然种群(表1),每个种群沿108国道选择15个采样点,每个样点间隔不少于30 m,采样点为国道两旁的路域生态系统.每个采样点分别选取一枝被泽兰实蝇寄生的和未被寄生的紫茎泽兰枝条,采集枝条上的成熟种子用信封装袋,带回实验室自然风干保存.

表1 研究种群自然概况Tab.1 The Natural Situation of the Sampled Populations
注:年均降水、年均温度来源于www.worldclim.org,为1970—2000年的平均值.
1.2 种子千粒重
每个样本随机选取200粒种子,用SHIMADZU电子天平(精度0.000 1 g)称重,然后计算其千粒重.
1.3 种子萌发
实验前用0.3%NaClO对种子进行表面消毒5 min,并用蒸馏水冲洗3次后,浸泡24 h备用.
采用滤纸法进行种子萌发实验.在直径9 cm的培养皿中垫两层滤纸,加入适量蒸馏水浸润,每个培养皿中均匀放入30粒预处理的种子.
整个实验在光照培养箱中进行,光照周期为12L:12D,温度周期为25:15℃,湿度为65%.种子萌发以种子露出胚根为标准,每天记录种子的萌发情况.连续5d无种子萌发视为实验结束.实验结束用数显式游标卡尺测量萌发幼苗的根长和芽长.
1.4 数据分析
种子重量以千粒重为指标,种子萌发特性包括萌发率、发芽势、发芽指数(衡量种子萌发速度的指标)、幼苗根长、幼苗芽长,其中:
萌发率=发芽总数/供试种子数 ×100%
发芽势[13]=前5天发芽种子数/供试种子数 ×100%
发芽指数[14]=∑Gt/Dt,式中Gt为第t日萌发的种子数,Dt为相应的萌发日数.
使用Shapiro-Wilk检验检验数据分布的正态性,并使用Levene’s检验分析数据的方差齐次性.为分析泽兰实蝇寄生对紫茎泽兰种子重量和萌发特性的影响,首先将四个种群的数据合并,利用线性混合效应模型(Linear Mixed Model,LMM)分析泽兰实蝇寄生与未寄生状态下紫茎泽兰种子千粒重及各萌发指标的差异显著性,检验时将泽兰实蝇寄生与否和种群作为混合效应,将采样点设置为随机效应以去除采样点的影响.
为比较种群间的差异,当数据满足正态分布和方差齐性时,使用单因素方差分析(One-way ANOVA)分析各指标在种群间的差异显著性,并使用最小显著性差异法(Least Significance Difference, LSD)进行种群间的两两比较;否则,使用Kruskal-Wallis检验分析各指标在种群间的差异显著性,使用Wilcoxon秩和检验分析数据的两组成对比较.
为探究各种群内泽兰实蝇寄生的影响,使用配对样本T检验(Paired-sample T test)分析各个种群寄生与未寄生状态下各指标的差异显著性.
所有数据分析在SPSS软件中完成,作图使用Origin软件完成.
2 结果与分析
2.1 泽兰实蝇寄生对紫茎泽兰种子质量的影响
1) 混合四个种群数据,比较被泽兰实蝇寄生与未被寄生的紫茎泽兰枝条的种子千粒重及萌发特性,结果表明(图1).
被寄生枝条的种子千粒重为(0.051±0.001)g,未被寄生的为(0.054±0.001)g,被寄生与未被寄生之间差异显著(LMM,P=0.037).
被寄生枝条的种子萌发率、发芽势、发芽指数及幼苗根长分别为87.66%、77.60%、7.73、25.20 mm,均高于未被寄生的86.08%、73.22%、7.47、24.80 mm,但寄生与未寄生状态下以上指标无显著差异(LMM,P>0.05),而芽长是被寄生的(1.79 mm)略低于未被寄生的(1.80 mm),两者间无显著差异(LMM,P=0.37)(图1).

图1 泽兰实蝇寄生与未寄生的紫茎泽兰枝条的种子千粒重及萌发指标的比较Fig.1 Comparison of 1000-seed Weight and Germination index between the Seeds Collected from Ageratina Adenophora Branches Galled or Un-galled by Procecidochares Utilis
注:图中不同字母表示两组间差异显著(P<0.05)
2) 比较种群内泽兰实蝇寄生对紫茎泽兰种子质量的影响.结果显示(表2):
泽兰实蝇的寄生降低了紫茎泽兰种子的千粒重,但影响仅在种群XC显著(Paired-sample T test,P=0.034),其他种群内寄生与非寄生状态下的差异不显著(Paired-sample T test,P>0.05)(表2).
泽兰实蝇寄生对紫茎泽兰种子萌发特性的影响在不同种群中有差异.在种群MN中,被寄生枝条的种子萌发率、发芽势、发芽指数及幼苗根长均显著高于未寄生枝条的种子(Paired-sample T test,P值分别为0.011、0.013、0.047、0.011),幼苗芽长是被寄生显著比未寄生的短(Paired-sample T test,P=0.044);而在其他种群中,泽兰实蝇的寄生或降低或增高萌发率、发芽势等,但寄生对所有萌发指标的影响都不显著(Paired-sample T test,P>0.05)(表2).

表2 各种群中寄生与未寄生状态下紫茎泽兰种子的重量和萌发特性(mean±SE)Tab.2 The Seed Weight and Germination Indexes of Ageratina Adenophora Seeds Collected from the Branches with or without Galls Induced by Procecidochares Utilis in Each Population
注:检验在各种群内分别进行,表中每个种群内同一列数据后的不同字母表示寄生与未寄生之间差异显著(P<0.05)
2.2 紫茎泽兰种子质量影响的种群差异
比较四个采样种群的种子千粒重及萌发特性,结果显示,种子的重量和萌发特性存在种群间差异(图2).
2.2.1 种子千粒重 泽兰实蝇寄生和未被寄生的紫茎泽兰枝条的种子千粒重在四个种群间均存在显著差异(One-way ANOVA,P值分别为0.028、0.020).多重比较结果显示,泽兰实蝇寄生状态下,种群PD的紫茎泽兰种子显著轻于HY、MN(P<0.05);未被寄生的状态下,PD的种子千粒重显著低于其他三个种群(P<0.05);无论在寄生还是未寄生状态下,其他三个种群的种子重量都不存在显著差异(图2A).
2.2.2 种子萌发率 泽兰实蝇寄生枝条的紫茎泽兰种子萌发率在四个种群间无显著差异(Kruskal-Wallis,P=0.572),而未被寄生枝条的种子存在差异显著(Kruskal-Wallis,P=0.015),其中MN种群的种子萌发率显著低于HY和XC种群(P<0.05)(图2B).
2.2.3 种子发芽势 泽兰实蝇寄生和未被寄生的紫茎泽兰枝条的种子发芽势在四个种群间均存在显著差异(Kruskal-Wallis,P值分别为0.014、0.018),泽兰实蝇寄生状态下,种群HY的种子发芽势显著低于MN和XC种群(P<0.05);未被寄生的状态下,种群XC显著高于其他三个种群(P<0.05)(图2C).
2.2.4 种子发芽指数 泽兰实蝇寄生的紫茎泽兰枝条的种子发芽指数在四个种群间不存在显著差异(One-way ANOVA,P=0.098),而未被寄生枝条的种子存在差异显著(One-way ANOVA,P=0.046),其中种群XC显著高于HY、MN、PD三个种群(P<0.05)(图2D).
2.2.5 幼苗根长 泽兰实蝇寄生枝条的紫茎泽兰幼苗根长在四个种群间差异显著(One-way ANOVA,P=0.001),而未被寄生的在四个种群间不存在显著差异(One-way ANOVA,P=0.943),在泽兰实蝇寄生的状态下,MN种群的幼苗根长最长,且显著高于HY和PD种群(LSD,P<0.05)(图2E).
2.2.6 幼苗芽长 泽兰实蝇寄生和未寄生枝条的紫茎泽兰幼苗芽长在四个种群间均无差异显著(One-way ANOVA,P值分别为0.067、0.196)(图2F).
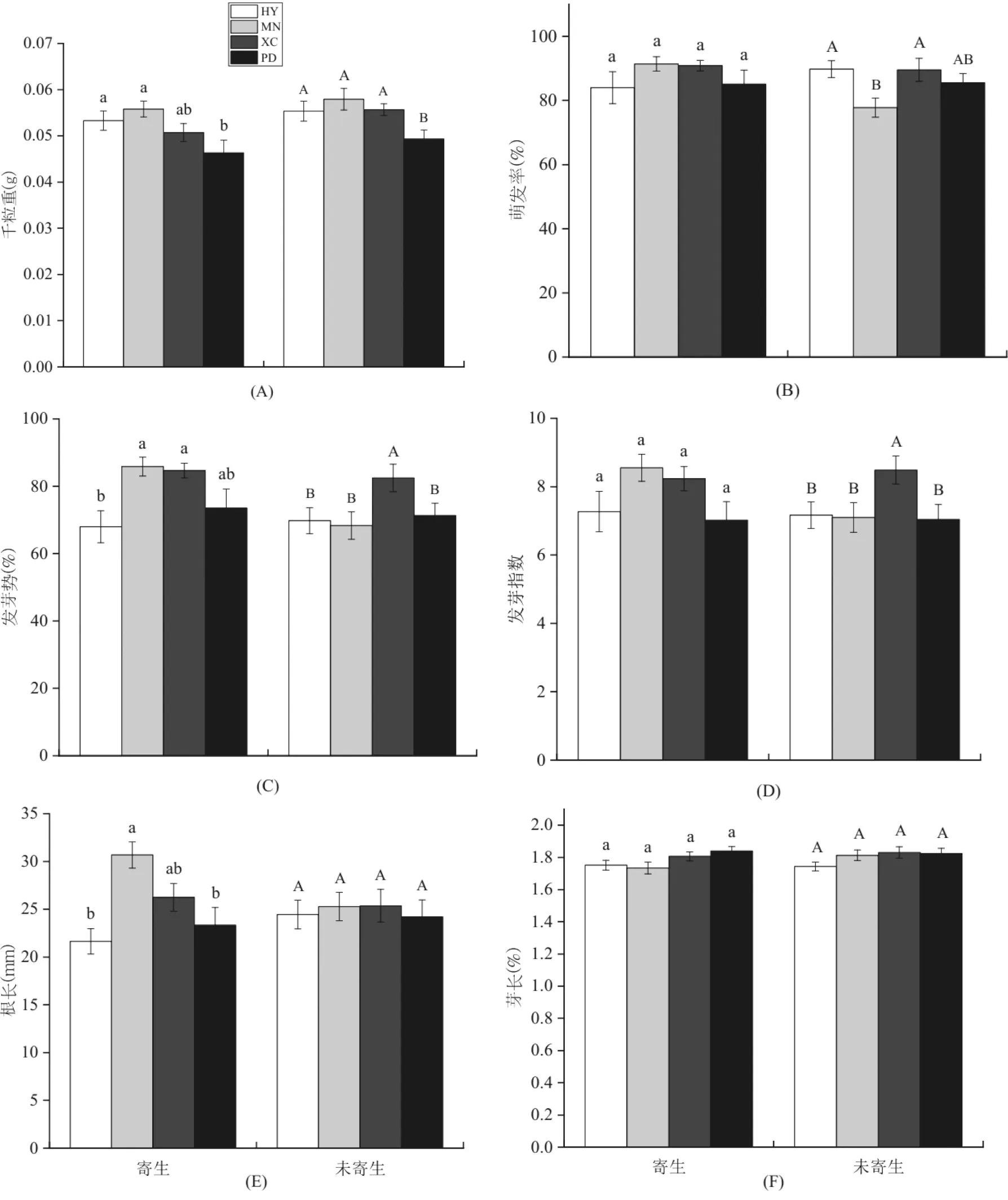
图2 泽兰实蝇寄生与未寄生对紫茎泽兰种子质量影响的种群间差异Fig.2 The Population Differences of Seed Quality of Ageratina Adenophora Affected by Parasitism and Non-parasitism of Procecidochares Utilis
注:图中不同小写字母表示泽兰实蝇寄生状态下的种群间差异在P=0.05水平显著;不同大写字母表示未寄生状态下的种群间差异在P=0.05水平显著.
3 讨论
种子是植物生活史的重要阶段,大多数入侵植物需要依靠种子的传播来建立和扩散种群[15].而紫茎泽兰种子是其快速传播、蔓延的主要途径[16].因此,紫茎泽兰种子的重量和萌发特性对其扩散起着非常重要的作用.
四川地区四个自然种群中,泽兰实蝇的寄生显著降低了被寄生枝条的种子千粒重(图1),其抑制作用在所有种群中均有表现(表2).营养供应会影响种子重量的增加[17].泽兰实蝇的幼虫一般侵入紫茎泽兰幼嫩的茎端、诱导紫茎泽兰形成膨大的虫瘿[18].同位素研究揭示,泽兰实蝇诱导的虫瘿是强烈的营养吸收库,刺激了营养物质的输入[19],从而与花朵和发育种子竞争营养,抑制了种子重量的增加.
种子重量一般会影响种子的萌发,但我们对紫茎泽兰种子萌发特性的比较显示,泽兰实蝇的寄生对被寄生枝条种子的萌发特性并无显著影响(图1),在一些种群内泽兰实蝇的寄生甚至可提高种子的萌发率(表2).这说明,虽然在研究区域内泽兰实蝇的寄生降低了紫茎泽兰种子的千粒重,但对其种子的萌发并未产生影响,在Arunava Datta等的研究结果中也发现紫茎泽兰的千粒重不影响其种子的萌发[20].这可能是因为紫茎泽兰能对泽兰实蝇的寄生产生一定的防御和适应对策.泽兰实蝇的寄生会使得紫茎泽兰自身对其产生较强的生理生化响应[11].姜丽娜等人研究发现泽兰实蝇寄生可能会促使紫茎泽兰改变自身的生长与防御能量分配,减少了营养和生长的投入,将更多的资源投向繁殖和防御[8].而在寄生状态下对繁殖投入的增加,可能在一定程度上弥补了虫瘿对营养的竞争,减少了对种子发育的影响.
在四川的四个种群中,泽兰实蝇寄生对紫茎泽兰的种子质量的影响是存在差异的(图2),尤其体现在MN种群,该种群紫茎泽兰的萌发特性在寄生与未寄生之间均存在显著差异.这一方面可能是因为不同入侵种群的土壤性质、水热环境条件不一样,如赵相健对不同地理种群紫茎泽兰种子萌发率的比较研究发现野外种子质量受到植株所处小生境的影响较大[21];另一方面可能是因为不同生境中泽兰实蝇的寄生强度不同.李爱芳等研究发现湿润生境下泽兰实蝇枝条寄生率(20.27%)显著高于干旱生境下的枝条寄生率(9.33%)[22].
泽兰实蝇作为防治紫茎泽兰的天敌,但利用其作为生防昆虫防治紫茎泽兰的效果受到很多因素的影响,导致不同地区、不同生境的防治效果不同,如有研究显示泽兰实蝇对紫茎泽兰的株高、生物量、结实量及种子发芽能力等生长和繁殖能力具有一定的抑制作用[9, 23],也有研究显示泽兰实蝇的寄生对紫茎泽兰的生长和繁殖并没有显著影响[11-12].从我们的结果看,泽兰实蝇的寄生在四川地区并未显著抑制紫茎泽兰种子的活力,尽管种子的重量受到了抑制,但萌发未受到抑制,这可能也是其入侵成功的原因之一.
