利用CRISPR/Cas9技术构建拟南芥IQM家族基因四突变体
2020-06-03魏丽茵吕天晓范甜周玉萍田长恩
魏丽茵 吕天晓 范甜 周玉萍 田长恩
摘 要之前,本研究组分别对IQM家族基因的功能进行了研究。不过,由于缺乏相应突变体而未能对该家族多基因同时突变所产生的生长发育改变进行研究。因此,利用CRISPR/Cas9基因编辑技术构建IQM家族多基因突变体。分别在IQM家族4个成员的基因组上设计靶点接头,通过同源重组构建成相应小载体sgRNA表达盒:LazAtU3d-AtU3b-AtU3d-AtU3b,并整合成pYLCRISPR/Cas9Pubi-N表达载体后转入拟南芥。经抗生素抗性筛选,共获得70株T0代植株。最后经过编辑靶点测序,得到2个IQM1~IQM4的四突变体株系。
关键词拟南芥;IQM;四突变体;CRISPR/Cas9
IQM家族由6个成员组成,命名为IQM1~IQM6。生物信息学分析表明,IQM蛋白均含有1个IQ基序,其N-端和C-端分别与豌豆重金属诱导蛋白6A和天花粉素具有较高的同源性,是1个全新的含IQ基序的家族(Zhou et al, 2010;田长恩等, 2013)。
本研究组长期从事IQM家族的功能研究,发现IQM1介导 CaM、光和部分非生物胁迫信号,调节气孔运动和根生长(Zhou et al, 2012)。还发现,IQM1可通过与CAT2互作而增强其活性,继而提高JA合成酶ACX2和ACX3的活性,增加JA含量和对灰质芽孢杆菌的抗性(Lv et al,2019)。IQM2可能通过影响光合作用而参与植物对病菌的反应(吴俊等,2017)。IQM3在叶、茎、花和根中的表达较强,但在荚果中很弱,与种子萌发及幼苗子叶膨大有关(周玉萍等,2009);IQM3还可能通过抑制赤霉素的生物合成而参与对幼苗的侧根数量和主根长度的调节(徐浩等,2019)。IQM4在植物非生物胁迫中发挥重要作用(萧文慧等,2016);还与种子休眠和萌发成正相关(Zhou et al, 2018)。
总之,IQM家族不同成员具有不同功能。因此,有必要构建拟南芥IQM家族的多突突变体,为深入研究拟南芥IQM家族基因的功能提供研究材料。
1 材料与方法
1.1 实验材料
以野生型拟南芥Columbia-0(Col-0)为基础进行基因编辑,同时敲除IQM基因家族中的4个成员IQM1~IQM4。质粒pCambia1301和大肠杆菌(菌株 E.Coli DH5α)由本实验室保藏;根瘤农杆菌(菌株 GV3101)由中国科学院华南植物园张明永教授友好转赠的;CRISPR/Cas9基因编辑体系小载体 pYLsgRNA-AtU3d/LacZ、pYLsgRNA-AtU3b和大载体pYLCRISPR/Cas9Pubi-N均由华南农业大学刘耀光教授友好赠送。
1.2 实验方法
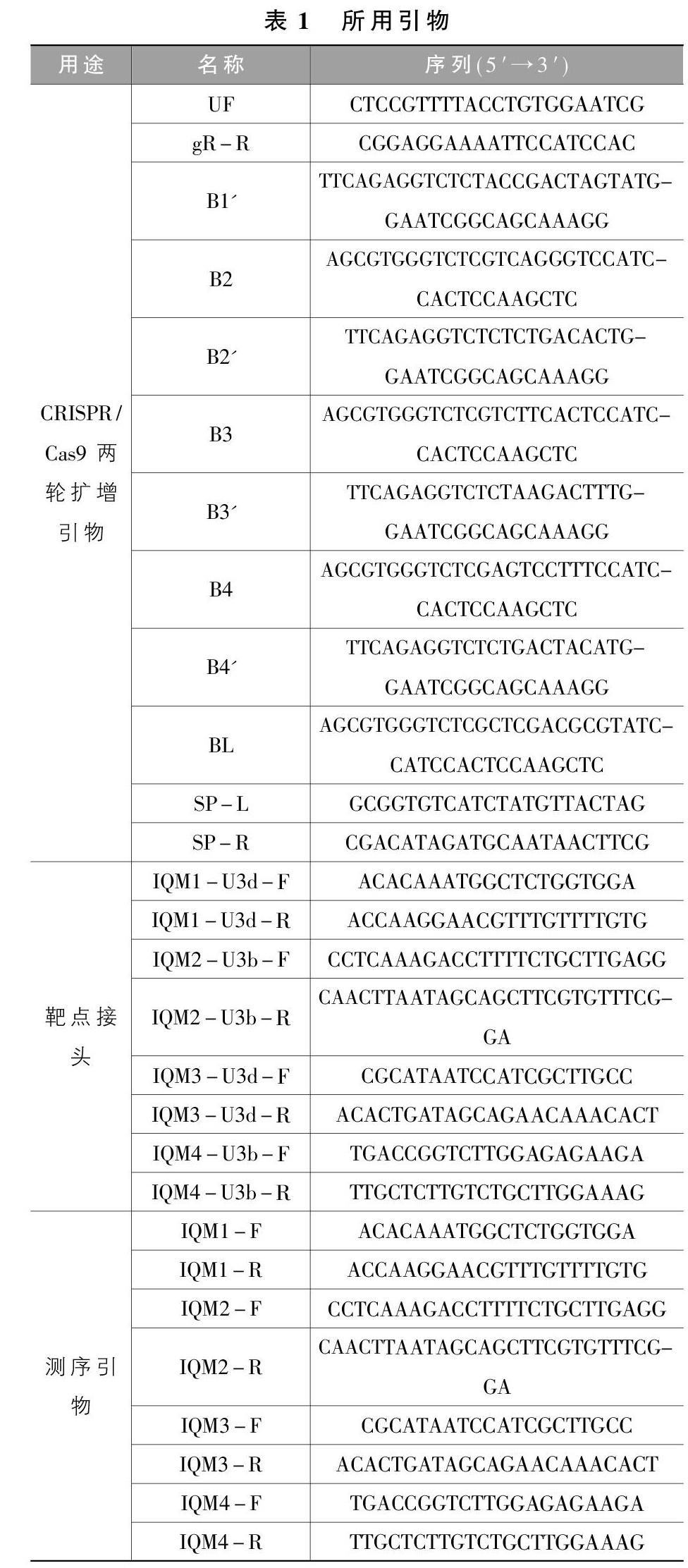
参照Ma和Liu (2016)的CRISPR/Cas9基因编辑技术进行。首先,在IQM1至IQM4的基因组序列中设计4对靶点接头(表1),分别退火连成小片段, 同源重组入经BsaI线性化的載体pYLsgRNA-AtU3d/LacZ和pYLsgRNA-AtU3b中,然后,进行gRNA表达盒扩增,回收预期扩增产物, 连接进双元载体并转化大肠杆菌;继而,进行转化菌落PCR鉴定, 选择阳性克隆测序验证;进一步,将验证正确的质粒转化农杆菌, 并用菌落PCR鉴定得到转化农杆菌;再进一步,通过农杆菌蘸染花蕾法转化野生型拟南芥, 收获种子并在含有卡拉霉素(Kan)的培养基上筛选出抗性植株即候选T0代编辑植株;最后,用靶点上、下游引物(表1)对候选株系进行PCR扩增, 将产物测序, 选出双链都成功编辑的转基因植株留种,供纯合子筛选。
2 结果与分析
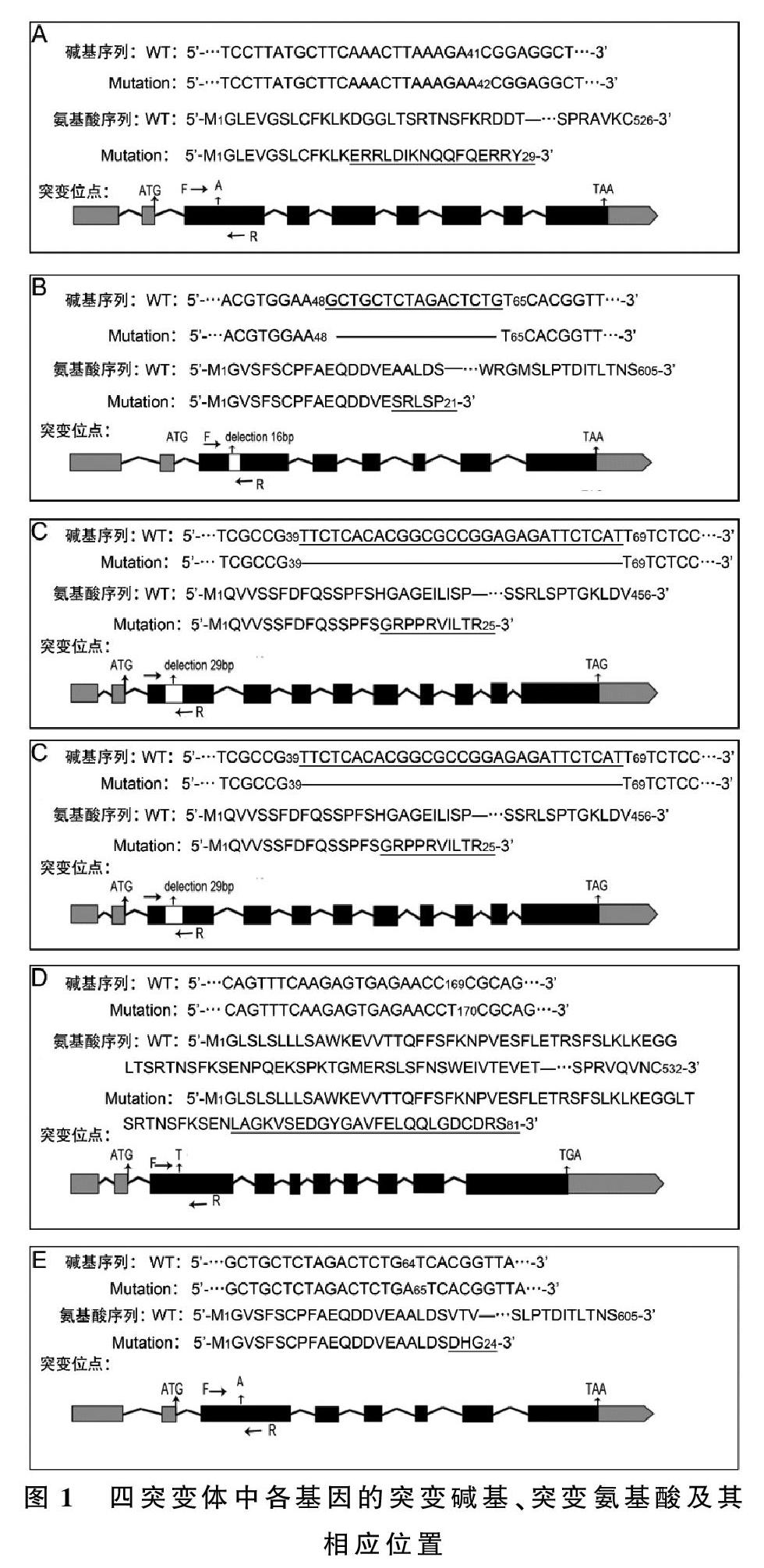
本文利用CRISPR/Cas9对野生型拟南芥的4个基因(IQM1、IQM2、IQM3和IQM4)进行编辑。对编辑的T0代进行抗生素抗性筛选,共获得70株抗性苗;继而,对这些苗及其后代进行跨靶点PCR产品测序,在T4代中筛选到2个四突变体纯合株系(iqm1 iqm2 iqm3 iqm4)-1和(iqm1 iqm2 iqm3 iqm4)-2,其相应的DNA序列和突变蛋白的氨基酸序列(见图1)。
与野生型(WT)相比,四突变体(iqm1 iqm2 iqm3 iqm4)-1的突变位点分别是:iqm1,从起始密码子ATG的A开始,在第41与42碱基之间插入1个A,导致其后序列发生移码突变并提前终止,产生1个只有29个氨基酸残基的突变蛋白,而IQM1共有526个氨基酸残基(图1-A);iqm2:从起始密码子ATG的A开始的第49至64碱基之间缺失16个碱基,导致其后序列发生移码突变并提前终止,产生1个只有21个氨基酸残基的突变蛋白,而IQM2共有605个氨基酸残基(图1-B);iqm3:从起始密码子ATG的A开始的第40至68碱基之间缺失29个碱基,导致其后序列发生移码突变并提前终止,产生1个只有25个氨基酸残基的突变蛋白,而IQM3有456个氨基酸残基(图1-C);iqm4:从起始密码子ATG的A开始在第170与171碱基之间上插入1个T,导致其后序列发生移码突变并提前终止,产生1个只有81个氨基酸残基的突变蛋白, 而野生型有532个氨基酸残基(图1-D)。
与上述突变体比较,另1个株系(iqm1 iqm2 iqm3 iqm4)-2只在IQM2中存在的不同:从起始密码子ATG的A开始在第64与65碱基之间插入1个A,导致其后序列发生移码突变并提前终止, 产生1个只有24个氨基酸残基的突变蛋白, 而IQM2有605个氨基酸残基(图1-E)。
3 讨论
本文利用CRISPR/Cas9技术对拟南芥IQM家族4个基因同时进行编辑,成功获得2个四突变体株系(iqm1 iqm2 iqm3 iqm4)-1和(iqm1 iqm2 iqm3 iqm4)-2。之后,将在上述四突变体的基础上,再编辑该家族其余的IQM5和IQM6基因,以期获得IQM全家族基因的六突变体。为研究IQM家族基因同时突变对植物生长发育以及对环境反应的影响提供材料。
參考文献
[1]田长恩,周玉萍.植物具IQ基序的钙调素结合蛋白的研究进展[J].植物学报, 2013,48:447-460.
[2]吴骏,林显宇,周玉萍,谢楚萍,田长恩.AtIQM2突变影响病菌侵染引起的荧光参数变化[J].广州大学学报(自然科学版),2017,16(3):44-47.
[3]周玉萍,赵军,何滔,程惠贞,段俊,田长恩。拟南芥IQM3基因的表达分析及其突变体的鉴定[J].热带亚热带植物学报,2009,17:365-370.
[4]徐浩,冯奕嘉,范甜,吕天晓,谢楚萍,周玉萍,田长恩.拟南芥IQM3基因突变减少幼苗的侧根数量和增加主根长度[J].植物生理学报, 2019, 55:629-634.
[5]萧文慧,宋俊威,黄小玲,周玉萍,田长恩.非生物胁迫对拟南芥iqm4基因表达的影响[J].科技视界,2016(16):10-11.
[6]Ma XL, Liu YG. CRISPR/Cas9-based multiplex genome editing in monocot and dicot plants[J].Curr Protoc Mol Biol,2016,115:31.6.131.6.21.
[7]Lv TX, Li XM, FT, Luo HT, Xie CP, Zhou YP, Tian CE. The calmodulin-binding protein IQM1 interacts with CATALASE2 to affect pathogen defense[J]. Plant Physiol, 2019, 181:1314-1327.
[8]Zhou YP, Duan J, Chen YZ, Yamamoto KT, Duan J, Tian CE. Sequence and expression analysis of the Arabidopsis IQM family[J]. Acta Physiol Plant, 2010, 32:191–198.
[9]Zhou YP, Duan J, Fujibe T, Yamamoto KT, Tian CE. AtIQM1, a novel calmodulin-binding protein, is involved in stomatal movement in Arabidopsis[J]. Plant Mol Biol, 2012, 79:333-346.
[10]Zhou YP, Wu JH, Xiao WH, Chen W, Chen QH, FT, Xie CP, Tian CE. Arabidopsis IQM4, a novel calmodulin-binding protein, is involved with seed dormancy and germination in arabidopsis[J]. Front Plant Sci, 2018, 9:721.
