天津周边地区烟粉虱隐种鉴定及其系统发育分析
2020-06-03张邓壮张艳玲胡明鑫刘佰明谷希树
张邓壮 张艳玲 胡明鑫 刘佰明 谷希树
摘 要:烟粉虱是一种世界性害虫,入侵世界许多地区,其携带的番茄黄化曲叶病毒(Tomato yellow leaf curl virus, TYLCV)给番茄生产造成了严重的损失。烟粉虱的种类繁多,给绿色防控带来许多困难。为了明确烟粉虱的种类,本试验主要对天津周边,还有山东、山西和湖北等省市10个地区不同寄主共16个烟粉虱种群进行了隐种鉴定及系统发育分析。结果表明,其中13个烟粉虱种群只有MED隐种,3个烟粉虱种群中MEAM1和MED隐种共存。MEAM1隐种烟粉虱的同源性高达100%,MED隐种烟粉虱的同源性为99.6%~100%,入侵我国的MED隐种可能起源于地中海西部地区。本研究明确了监测地区烟粉虱隐种及其进化关系,为当地烟粉虱的防控提供了科学依据。
关键词:烟粉虱;系统树;隐种;系统发育
中图分类号:Q966 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2020.03.004
Species Identification and Phylogenetic Analysis of Bemisia tabaci in Tianjin and Surrounding Areas
ZHANG Dengzhuang1,2, ZHANG Yanling3, HU Mingxin2,4, LIU Baiming2, GU Xishu2
(1.College of Life Science, Tianjin Normal University, Tianjin 300387, China; 2.Tianjin Institute of Plant Protection, Tianjin 300384, China; 3.Vegetable Technology Promotion Station of Ninghe District, Tianjin City,Tianjin 301500, China;4.College of Modern Agriculture and Ecological Environment, Heilongjiang University, Harbin, Heilongjiang 150080, China)
Abstract: As a worldwide pest, Bemisia tabaci has invaded many regions of the world. The tomato yellow leaf curl (TYLCV) virus it carries has caused serious losses. There are many species of Bemisia tabaci, which brings many difficulties to green prevention and control. In order to clarify the species of Bemisia tabaci, in this experiment, a total of 16 species of Bemisia tabaci in different regions of Tianjin and its vicinity, Shandong Shanxi,Hubei were collected to perform species identification and phylogenetic analysis. The results showed that 13 populations only contained Bemisia tabaci MED, 3 populations contained both Bemisia tabaci MEAM1 and Bemisia tabaci MED. Among our collected samples, the homology of Bemisia tabaci MEAM1 was 100%, and The homology of Bemisia tabaci MED was 99.6%~100%.China's MED may originate in the western Mediterranean. This study illustrated the species and evolutionary relationships of Bemisia tabaci in the monitored areas, providing a scientific basis for the prevention and control of Bemisia tabaci in these areas.
Key words: Bemisia tabaci; phylogenetic tree; species; phylogenetic
煙粉虱Bemisia tabaci(Gennadius)是一种多食性农业害虫,能危害600多种寄主植物,为害方式包括通过吸取植物汁液、分泌蜜露影响光合作用直接危害植物,还可以通过传播200多种植物病毒间接危害植物[1-2],番茄黄化曲叶病毒能侵染多种植物,在番茄上的为害尤为严重,严重时导致绝收[3]。烟粉虱有非常复杂的遗传结构,目前认为烟粉虱至少是由36个隐种组成的物种复合体[4-5]。其中MEAM1隐种(B型)和MED隐种(Q型)烟粉虱入侵性最强、为害最重。烟粉虱侵入我国时间较长,在20世纪90年代MEAM1烟粉虱就开始入侵我国,之后迅速蔓延到全国各地。而MED烟粉虱则入侵较晚,于2003年在云南首次发现,但是在最近十年中MED烟粉虱已经取代了MEAM1烟粉虱成为中国主要的烟粉虱类型[6-7]。不同烟粉虱隐种具不同的生物学特性,如MEAM1和MED隐种烟粉虱的传播植物病毒能力和对杀虫剂抗性存在明显差异[8-9]。MED隐种的遗传多样性要比MEAM1更加丰富,因而其适应环境的能力更强,能更好的抵御外界不利因素,例如农药、化学药剂等,MED隐种对新烟碱类杀虫剂的抗性更强[10]。近年来,国内研究者已逐渐重视田间烟粉虱隐种的监测,监测的地区包括新疆、北京、湖南、江苏、陕西、湖北等,为科学防治烟粉虱提供了理论依据[11-15]。本研究主要立足于天津各区及其周边地区,同时与其他省份的地区作比较,采用烟粉虱mtDNA COI基因作为分子标记,对天津各区、北京、山东、山西和武汉等地区不同寄主植株的烟粉虱种群进行隐种鉴定及系统发育分析,旨在为合理的防治天津地区烟粉虱提供科学依据。
1 材料和方法
1.1 材 料
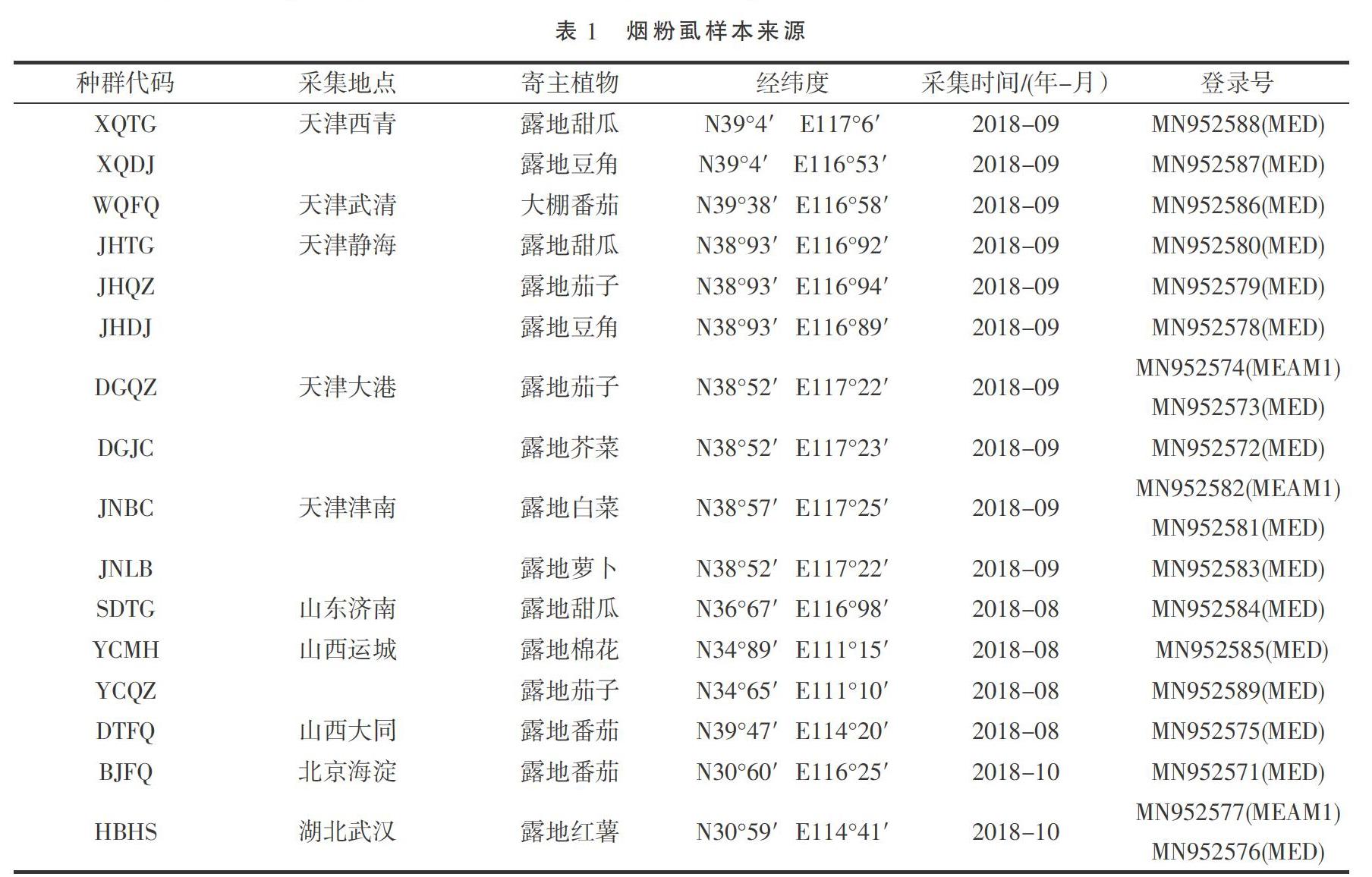
采集了天津(西青、靜海、大港、津南、武清)、北京海淀、山东济南、山西运城、山西大同以及湖北武汉共10个地区不同寄主的16个种群的烟粉虱成虫,所采集到的样品直接浸泡于1.5 mL装有无水乙醇的离心管中,保存在-20 ℃冰箱中备用。具体采集信息及登录号见表1。
1.2 方 法
1.2.1 单头烟粉虱总DNA的提取 参照柳洋[16]的方法,使用KAPA DNA提取试剂盒对烟粉虱DNA进行提取。用昆虫针取一头烟粉虱至2 mL离心管中,加入灭菌后的研磨珠,向离心管中加入30 μL配好的DNA提取混合液,将加入提取液的离心管放入组织破碎仪中研磨1 min,8 000 rpm离心30 s,取上清至新的PCR管中并放入PCR仪中进行酶促反应,反应程序为:蛋白酶75 ℃裂解10 min,95 ℃灭活酶5 min。-20 ℃冰箱长期保存。
1.2.2 mtCOI扩增及酶切鉴定烟粉虱生物型 进行PCR反应扩增mtCOI基因片段,反应程序参照柳洋[16],引物为:C1-J-2195'-TTGATTTTTTGGTCATCCAGAA
GT-3')和R-BQ-2819(5'-CTGAATA TCGRCGAGGCA
TTCC-3'),扩增产物大小为620 bp左右。
参照 Chu 等[17]的方法,应用限制性内切酶 AseI 酶切已扩增到的 620 bp mtCOI 基因片段,通过酶切产物可判断烟粉虱的生物型。
用1.5%琼脂糖凝胶电泳检测PCR产物,用凝胶成像仪观察并记录结果。
1.2.3 mtCOI基因序列测序及数据分析 进行PCR反应,反应程序参照 Simon 等[18]的方法,引物为:C1-J-2195(5'-TTGATTTTTTGGTCATCCAGAAGT-3')和L2-N-3014(5'-TCCAA TGCACTAATCTGCCA
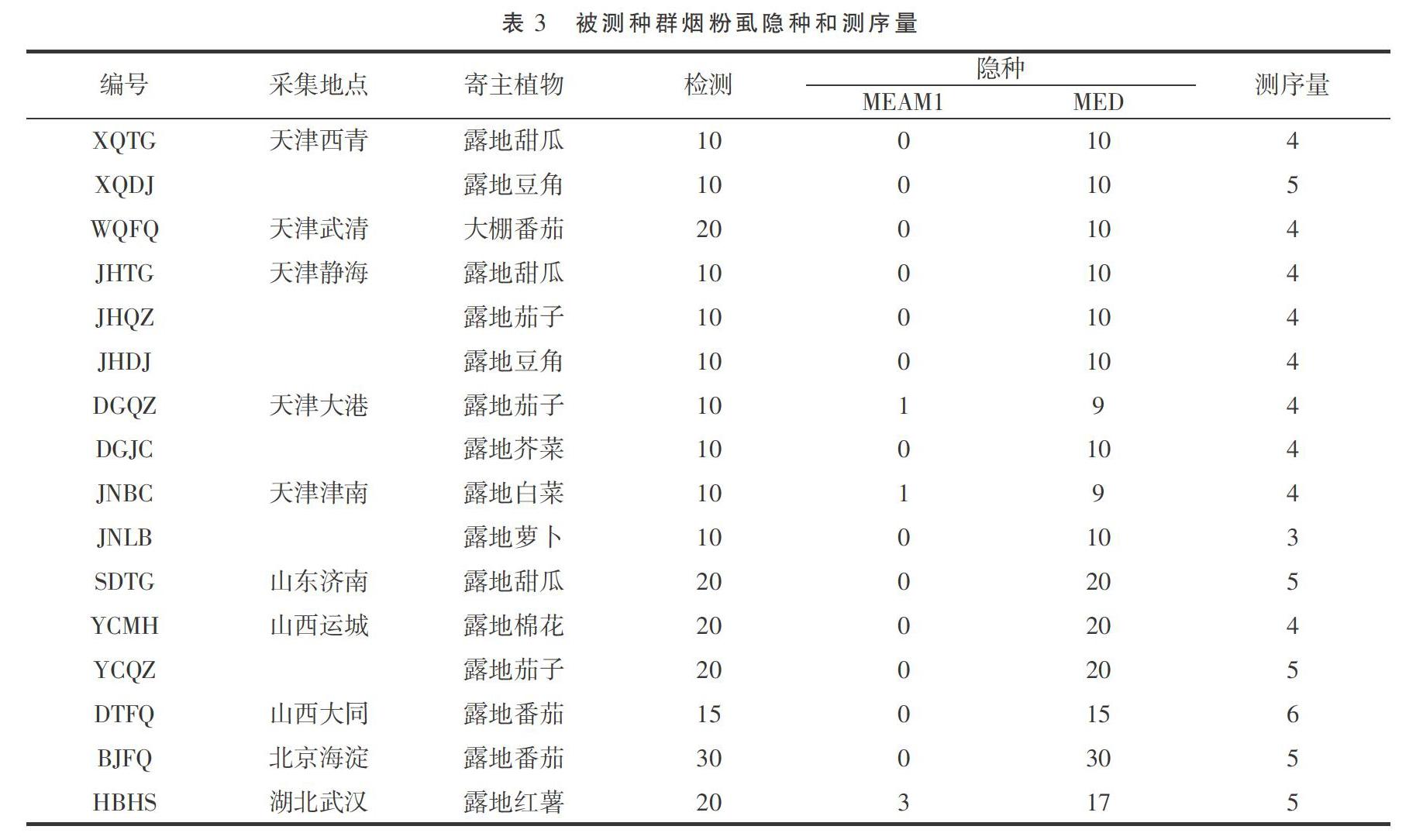
TATTA-3'),扩增产物为COI基因3'末端的部分序列,大小为840 bp左右。将PCR产物纯化后送往天津擎科生物科技有限公司进行双向测序,每个种群至少检测3头烟粉虱。
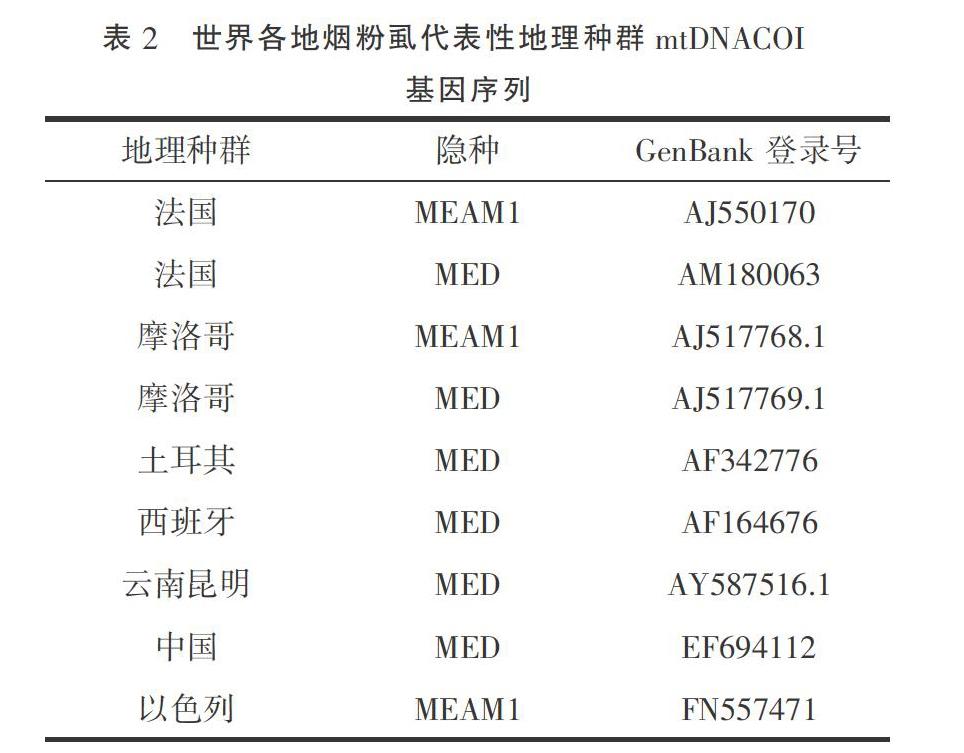
将测序结果提交 GenBank,获取登录号(表1),通过BLAST方法检索有代表性国外烟粉虱COI序列(表2)。在16个种群中,从13个只有MED烟粉虱的种群各抽选一条测序结果;在MEAM1、MED烟粉虱共存的3个种群中,不同的隐种烟粉虱各抽选一条测序结果。以非洲小粉虱[Bemisia afer( Priesner & Hosny)](AF418673.2)为外群[19]。运用MEGA 7软件,根据 Kimura 2- paramter 模型计算出采样地区烟粉虱与国内外不同地理种群及不同生物型烟粉虱的进化分歧矩阵,利用邻位法(neighbor-joining,NJ)和最大似然法(maximum likelihood,ML)分别构建系统发育树,各分支进行1 000次的重复检验。
2 结果与分析
2.1 鉴定不同地区烟粉虱生物型检测结果
MED烟粉虱的 mtCOI 基因片段存在一个 AseI 酶切位点,因而可以被酶切成两个片段(498 bp 和122 bp),而 MEAM1 烟粉虱的 mtCOI 基因由于不存在 AseI 酶切位点,因此不能被切开[10]。
结果显示16个种群中,有13个种群的烟粉虱都是MED型,有3个种群是MEAM1、MED混合,且混合种群中以MED为主(表3)。
2.2 根据mtDNA COI序列构建进化分歧和系统发育树
2.2.1 构建进化分歧 非洲小粉虱和其他种群烟粉虱的遗传距离都在28%以上,MEAM1烟粉虱和MED烟粉虱的遗传距离基本在4.7%~5.6%之间,我们检测地区的MED烟粉虱的遗传距离在0%~0.4%之间,与中国Q型和云南昆明烟粉虱的遗传距离基本在0.2%之间,低于已知MED烟粉虱的内部差异。三个地区的MEAM1烟粉虱同源性高达100%(表4、表5)。
2.2.2基于mtDNA COI基因构建系统发育树 使用NJ法构建系统树,各分支均进行1 000次的重复检验(图1)。从进化树中可以看出, MED烟粉虱以99%的置信度聚为一个分支,且测得的所有MED烟粉虱与来自法国、摩洛哥和西班牙的MED烟粉虱亲缘关系更近,而来自土耳其的则单独聚为一支。所有的MEAM1烟粉虱聚为一支。使用ML法和NJ法构建的系统发育树结果相似(图2)。
3 讨 论
近十年来,我国烟粉虱的优势隐种逐渐由MED代替了MEAM1,其传播的TYLCV对我国大部分地区的番茄造成严重的危害,对杀虫剂抗药性的问题也日益突出[20]。烟粉虱是一个复杂的物种复合体,不同隐种之间传播TYLCV和对杀虫剂抗药性也都有所差异。明确天津各地区烟粉虱的隐种现状,与世界各地不同隐种烟粉虱之间的系统发育分析,可对外来入侵烟粉虱的防治具有重要的指导意义。
本文通过 mtDNA COI基因序列分析鉴定了天津各区及其附近、山东、山西、湖北等不同地区烟粉虱的隐种组成和系统生物发育分析,检测的隐种和系统发育树共同表明在监测的各个地区的不同寄主植株上的烟粉虱绝大部分是都是MED烟粉虱,只有极少数的种群存在MEAM1烟粉虱,这与现在MED烟粉虱是主要隐种的现状是相符的。在2009年,Pan等[7]调查了天津西青区的烟粉虱隐种,烟粉虱全部是MEAM1隐种,而今调查天津各区烟粉虱隐种几乎都是MED,与褚栋等[21]的结论也一致。
许丽丽等[22]推测中国MED烟粉虱可能起源于地中海西部地区,我们调查地区的MED隐种与来自法国、摩洛哥和西班牙的MED烟粉虱亲缘关系更近,土耳其的则单独聚为一支,结果相似,为这一推测再添实例,更进一步的证明推测的正确性。
不同隐种对寄主植株的危害程度是不同的,MED烟粉虱在番茄上的危害程度比MEAM1烟粉虱更加严重[23]。有研究表明,MED烟粉虱比MEAM1烟粉虱对新类烟碱农药具有更高的抗性[24]。这意味着这些地区的烟粉虱种群对新类烟碱农药有较高的抗性,而目前烟粉虱的治理主要通过喷施农药等化学途径,这为防治烟粉虱加大了难度。
本研究中由于采样地点、数量、方式等局限性,不排除13个种群只检测出MED烟粉虱的种群是混合种群的可能,因此需要持续对天津各地区烟粉虱进行隐种鉴定及系统发育分析,明确各地区烟粉虱的详细情况,为科学的防治烟粉虱提供理论依据。
参考文献:
[1]BROWN J K. Phylogenetic biology of the Bemisia tabaci sibling species group[M]//Bemisia: bionomics and management of a global pest. Springer, Dordrecht, 2009: 31-67.
[2]HOGENHOUT S A, AMMAR E D, WHITFIELD A E, et al. Insect vector interactions with persistently transmitted viruses[J]. Annual review of phytopathology, 2008, 46(1):327-359.
[3]PICD B, DIEZ M J, NUEZ F.Viral diseases causing the greatest economic losses to the tomato crop.II. The Tomato yellow leaf curl virus-a review[J]. Scientia horticulturae,1996,67(3):151-196.
[4]HU J, DE BARRO P, ZHAO H, et al. An extensive field survey combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China[J]. PloS one,2011,6:el061.
[5]劉银泉,刘树生.烟粉虱的分类地位及在中国的分布[J].生物安全学报,2012,21(4):247-255.
[6]CHU D, ZHANG Y J, BROWN J K, et al. Theintroduction oftheexotic Q biotypeofBemisia tabacifromthemediterranean REGION INTO CHINA ON ORNAMENTAL CROPS[J]. Florida entomologist, 2006, 89(2):168-174.
[7]PAN H, CHU D, GE D, et al. Further spread of and domination by Bemisia tabaci(Hemiptera: Aleyrodidae) biotype Q on field crops in China[J]. Journal of economic entomology, 2011, 104(3):978-985.
[8]HOROWITZ A R, DENHOLM I, GORMAN K, et al. Biotype Q of Bemisia tabacii dentified in Israel[J]. Phytoparasitica, 2003, 31(1):94-98.
[9]BROWN J K, CZOSNEK H. Whitefly transmission of plant viruses[J]. Advances in botanical research, 2002, 36(2):65-76.
[10]PAN H P, PREISSER E L, CHU D, et al. Insecticides promote viral outbreaks by altering herbivore competition[J]. Ecological applications, 2016, 25(6):1585-1595.
[11]王少丽, 张友军, 李如美,等. 北京和湖南烟粉虱生物型及其抗药性监测[J]. 应用昆虫学报, 2011, 48(1):27-31.
[12]谭永安, 肖留斌, 孙洋,等. 江苏烟粉虱生物型鉴定与危害程度分析[J]. 环境昆虫学报, 2012, 34(3):277-282.
[13]赵瑞华, 邓振山, 贺晓龙. 陕西地区烟粉虱生物型的鉴定及其危害分析陕西地区烟粉虱生物型的鉴定及其危害分析[J]. 延安大学学报(自然科学版), 2015, 34(4): 82-87.
[14]周新改,马伟华,刘美刚. 湖北省棉花主产区烟粉虱生物型分布及系统发育分析[J]. 植物保护, 2011, 37(5):72-76.
[15]曹骞,李晶,买热木古丽·克依木,等. 新疆地区烟粉虱生物型的区域分布及其携带的番茄黄化曲叶病毒检测[J]. 昆虫学报, 2013, 56(6):652-664.
[16]柳洋. 中国烟粉虱生物型分布、带毒率及抗药性监测[D].北京:中国农业科学院,2015:6-9.
[17]CHU D, ZHANG Y J, WAN F H. Cryptic invasion of the exotic Bemisia tabaci biotype Q occurred widespread in Shandong Province of China[J]. Florida entomologist, 2010,93(2): 203-207.
[18]SIMON C, FRATI F, BECKENBACH A, et al. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase Chain reaction primers[J]. Annals of the entomological society of America, 1994, 87(6):651-701.
[19]罗晨.烟粉虱的寄主调查、生物学特性和分子系统学研究[D].北京:中国农业大学,2002.
[20]褚栋, 张友军.近10年我国烟粉虱发生为害及防治研究进展[J]. 植物保护, 2018, 44(5):56-60.
[21]褚栋,潘慧鹏,国栋,等.Q型烟粉虱在中国的入侵生态过程及机制[J].昆虫学报,2012,55(12):1399-1405.
[22]许丽丽,蔡力,沈伟江,杜予州.中国部分地区烟粉虱生物型种类及系统发育关系分析[J].应用生态学报,2014,25(4):1137-1144.
[23]MUNIZ M, NOMBELA G. Differential variation in development of the B-and Q-Biotypes of Bemisia tabaci (Homoptera: Aleyrodidae) on sweet pepper at constant Temperatures[J]. Environmemology, 2001, 30(4):720-727.
[24]RAUCH N, NAUEN R. Identification of biochemical markers linked to neonicotinoid cross resistance in Bemisia tabaci(Hemiptera: Aleyrodidae)[J]. Archives of insect biochemistry & physiology, 2010, 54(4):165-176.
