基于SSR标记的燕山板栗种质资源遗传多样性分析
2020-05-04张馨方张树航王广鹏
张馨方 张树航 李 颖 郭 燕 王广鹏
(河北省农林科学院 昌黎果树研究所,河北 昌黎 066600)
板栗(CastaneamollissimaBL.)是我国的重要经济林树种,广泛分布在大江南北的24个省(自治区、直辖市)。中国北方的燕山区既是我国板栗的主要产区,又是世界上糖炒型板栗的最佳产区,该区域主要包括北京市的怀柔、密云区和河北省的遵化、迁安、迁西、兴隆、青龙和宽城等县(市)[1],其出产的栗果在国内外市场上被统称为燕山板栗。得益于得天独厚的地理条件和资源优势,燕山板栗以口味甘甜、肉质细糯、品质优良以及适宜糖炒而享誉国际市场,是我国极少数具有出口优势的经济林产品。
经过长期的种子实生繁殖、自然和人工选择,燕山区拥有极其丰富的板栗种质资源,这些资源具有不同的变异类型和独特的农艺性状,其中更是蕴含着许多有待进一步发掘的优异基因,是育种工作良好的资源材料,因此,对燕山种质资源进行遗传多样性研究十分必要。目前,利用DNA分子标记技术对栗属植物进行遗传多样性的研究报道越来越多,周连第等[2]采用AFLP技术对86个板栗品种进行遗传多样性研究,证明了以AFLP-荧光法分析板栗品种资源分子水平遗传多样性是可行的方法。向晖等[3]和龚榜初等[4]分别采用SRAP和ISSR分子标记对我国锥栗自然居群进行了遗传多样性分析,并提出了湘西地区可能是锥栗的次生分布中心和现代遗传多样性分布中心。项艳等[5]利用RAPD分子标记手段对14个安徽省板栗品种进行遗传多样性分析,建立了板栗品种的分子检索表,并提出了重点保存品种。艾呈祥等[6]利用SSR引物对26份山东省主栽板栗品种进行遗传多样性分析,探讨了鲁中南地区和泰沂山区不同板栗品种的起源问题。以往的研究,或选用燕山板栗材料数量有限,或取样范围没有涵盖燕山板栗的主产区,整体而言前人对燕山板栗分子水平上的遗传多样性研究相对较粗浅。SSR分子标记技术早被证实具有共显性遗传、操作便捷和结果稳定而准确等优良特点,是遗传多样性分析的重要工具。基于日本栗和欧洲栗[7-10]遗传多样性评价开发的SSR引物在国外早有报道,这批引物在中国板栗、茅栗、锥栗也获得了良好的扩增效果[11],说明它们在栗属植物间具有较高通用性。本研究拟通过筛选多态性较好的SSR引物,对燕山地区7个县域群体以及1个外来群体共151份板栗种质资源进行遗传多样性分析,并构建聚类树状图和主坐标分析图,以期为燕山板栗种质资源的遗传背景明晰和以遗传距离为参考的亲本选择利用提供依据。
1 材料与方法
1.1 材料
试验所用151份板栗种质资源(均是由实生大树选育的地方种系)来自燕山地区的7个不同县域群体,包括青龙县(25份)、遵化市(17份)、昌黎县(14份)、迁西县(29份)、宽城县(23份)、兴隆县(26份)和怀柔区(8份),以及1个太行山区的邢台板栗资源9份,全部定植于河北省农林科学院昌黎果树研究所板栗种质资源圃。2018年5—6月,选取板栗新鲜健康嫩叶,放于装有硅胶的自封袋中干燥,在室温下保存备用。供试材料及原产地位置信息见表1 和表2。
1.2 基因组DNA的提取
采用改良的CTAB法[12]提取板栗叶片基因组DNA,用1%琼脂糖凝胶电泳及微量分光光度计进行DNA质量和浓度检测,合格的DNA样品放置于-20 ℃保存备用。
1.3 SSR反应体系及扩增条件
PCR反应体系20 μL,包括双蒸水13.35 μL,10×Buffer 2.0 μL,MgCl22.0 μL(25 mmol/L),dNTP 1.2 μL(浓度2.5 mmol/L),TaqDNA聚合酶0.2 μL(浓度5 units/μL),引物0.25 μL(20 μmol/L),DNA模板30 ng。扩增条件如下:94 ℃ 预变性5 min;94 ℃变性50 s;55 ℃(因引物不同而定)退火50 s;72 ℃延伸50 s,循环30次;后72 ℃延伸7 min。
1.4 SSR引物筛选及产物检测
为了得到清晰的SSR指纹图谱,从220对引物中筛选出条带清晰、多态性较好的21对引物用于全部样品分析,220对引物来源主要包括板栗及蔷薇科模式植物已公开发表的SSR引物序列,供试引物具体信息见表3[13-19]。
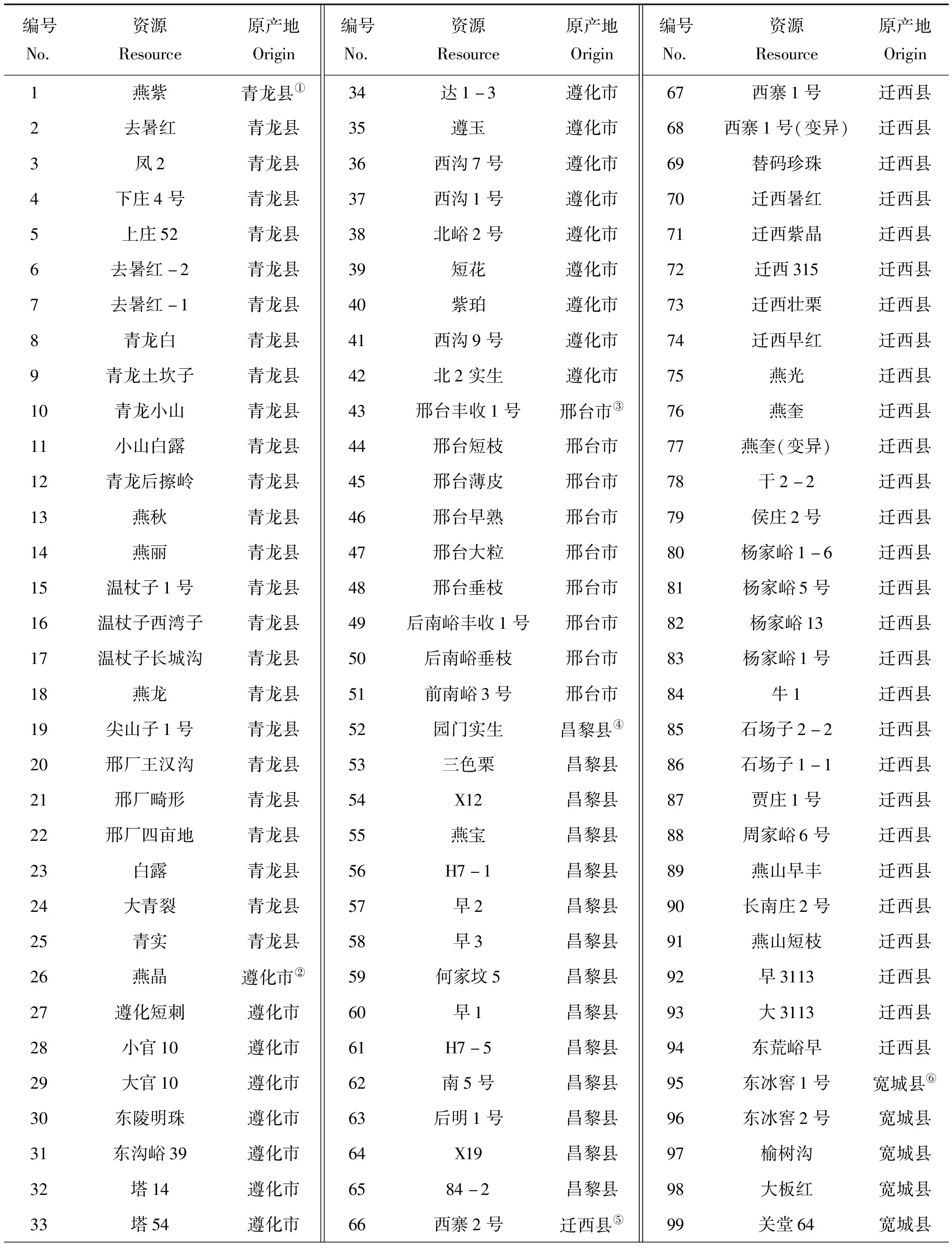
表1 供试板栗材料
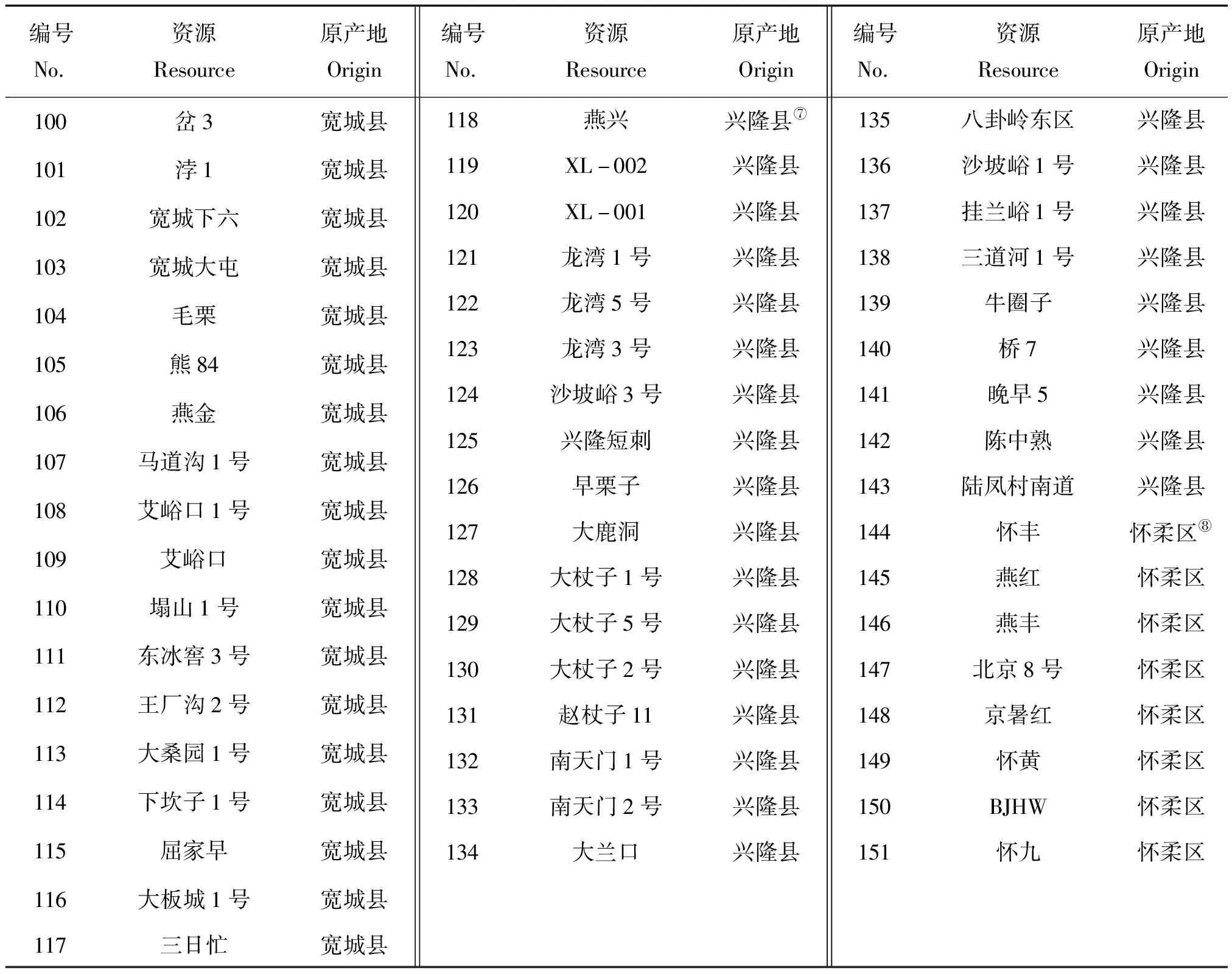
表1(续)
注:①④属河北省秦皇岛市;②③属河北省;⑤属河北省唐山市;⑥⑦属河北省承德市;⑧属北京市。
Note: ①④ belongs to Qinhuangdao, Hebei.② and ③ belong to Hebei Province.⑤ belongs to Tangshan, Hebei.⑥ and ⑦ belong to Chengde, Heibei.⑧ belongs to Beijing.

表2 供试材料原产地位置信息
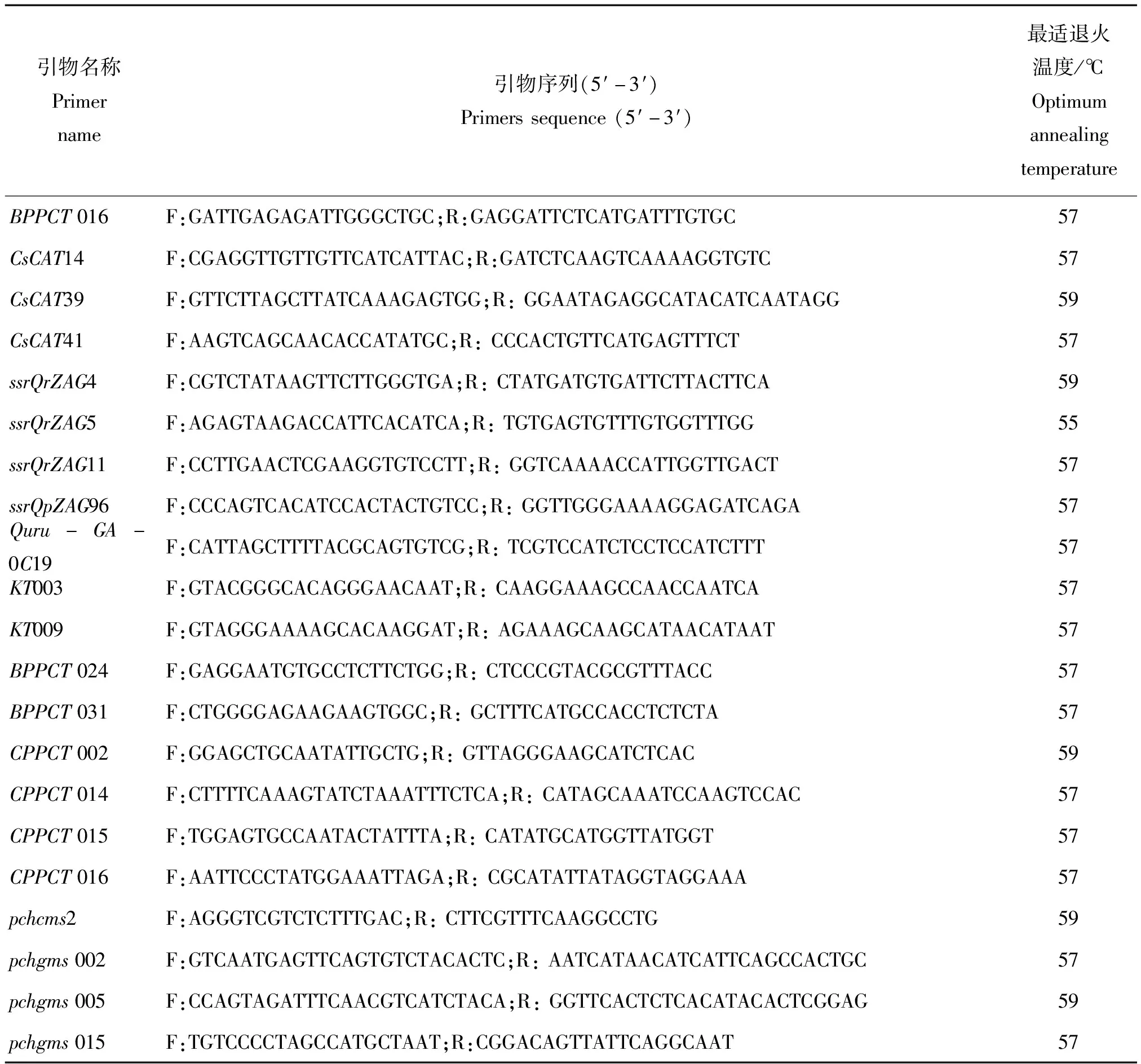
表3 筛选出的21对SSR引物
扩增产物用6%聚丙烯酰胺凝胶分离,在90 W恒功率下电泳40~50 min,银染法[20]染色,待出现清晰条带后置于灯箱上拍照。
1.5 数据统计分析
以0、1统计SSR扩增带型,并建立由“0、1”组成的原始数据矩阵。在相同迁移位置上,有条带记为“1”,无带记为“0”,构建二元数据矩阵。使用POPGENE Version 1.32软件计算观测等位基因数目(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)、Shannon’s信息指数(I)、多态位点数目(Np)和多态位点百分率(PPB),基因流(Nm)和遗传一致度等参数。通过软件PIC-CALC计算引物多态性信息含量(PIC, polymorphism information content)。根据遗传一致度,利用NTSYS Version 2.10软件中的SAHN模块和UPGMA方法进行聚类分析,使用EIGEN模块进行主坐标分析。
2 结果与分析
2.1 SSR分子标记遗传多样性
采用SSR标记对8个群体151份板栗资源遗传多样性进行分析。21对SSR引物共检测到71个等位基因,其中全部为多态性等位基因,多态率达100%,每对引物获得的等位基因数目为2~6,平均3.38个,扩增产物大小在100~300 bp。SSR引物的多态性信息含量(PIC)变幅为0.614 5~0.972 3,平均为0.866 8。部分扩增产物的电泳扫描照片见图1。
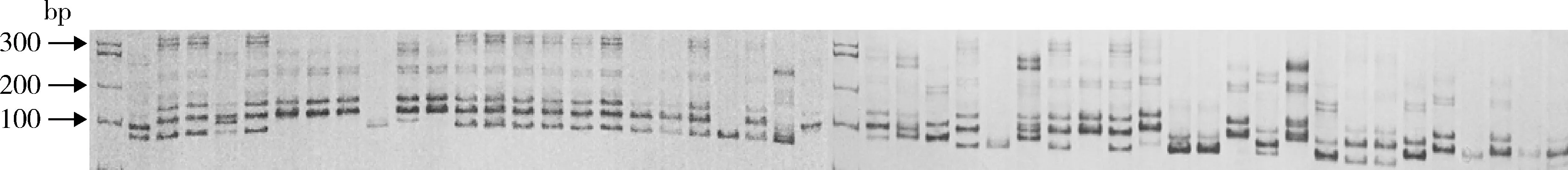
图1 引物KT009扩增结果
2.2 不同板栗群体间遗传多样性比较
计算8个板栗群体各遗传多样性参数(表4)。其中,各群体资源份数为8~29份不等,迁西群体包含资源份数最多,但各项遗传多样性参数均不是最高,而怀柔群体资源量最小,Nei’s多样性指数H=0.309 9,Shannon’s信息指数I=0.468 0均最大,从分子水平证明了资源份数多,遗传多样性不一定最大的论断[21]。多态位点数和多态位点百分率是反映遗传多态性的重要指标,遵化群体多态位点数目(Np=69)、多态位点百分率(PPB=97.18%)及观测等位基因数(Na=1.971 8)、有效等位基因数(Ne=1.521 9)均最大,邢台群体多态位点数目(Np=61)、多态位点百分率(PPB=85.92%)及观测等位基因数(Na=1.859 2)、有效等位基因数(Ne=1.456 3)最小。其中,有7个群体多态性位点百分率>90%,说明调查各群体的遗传多态性普遍较高。有效等位基因数Ne是反映群体遗传变异大小的一个指标,与等位基因的频率有关,与观测等位基因数Na相比,青龙和兴隆群体的有效等位基因数较小,说明青龙和兴隆板栗资源具有的极端频率的等位基因相对较多;而怀柔和邢台群体的有效等位基因数Ne更接近观测等位基因数Na,表明等位基因在这些群体中分布较均匀。Nei’s 多样性指数和 Shannon’s 信息指数都是反映群体遗传多样性水平的重要参数,8个群体中怀柔和遵化板栗群体遗传多样性水平高,而邢台群体遗传多样性水平相对较低;供试板栗资源总的 Nei’s 多样性指数H=0.302 9,Shannon’s 信息指数I=0.465 5,说明燕山板栗种质资源遗传多样性水平普遍较高。其中怀柔区和遵化市是燕山板栗资源遗传多样性最丰富的地区。
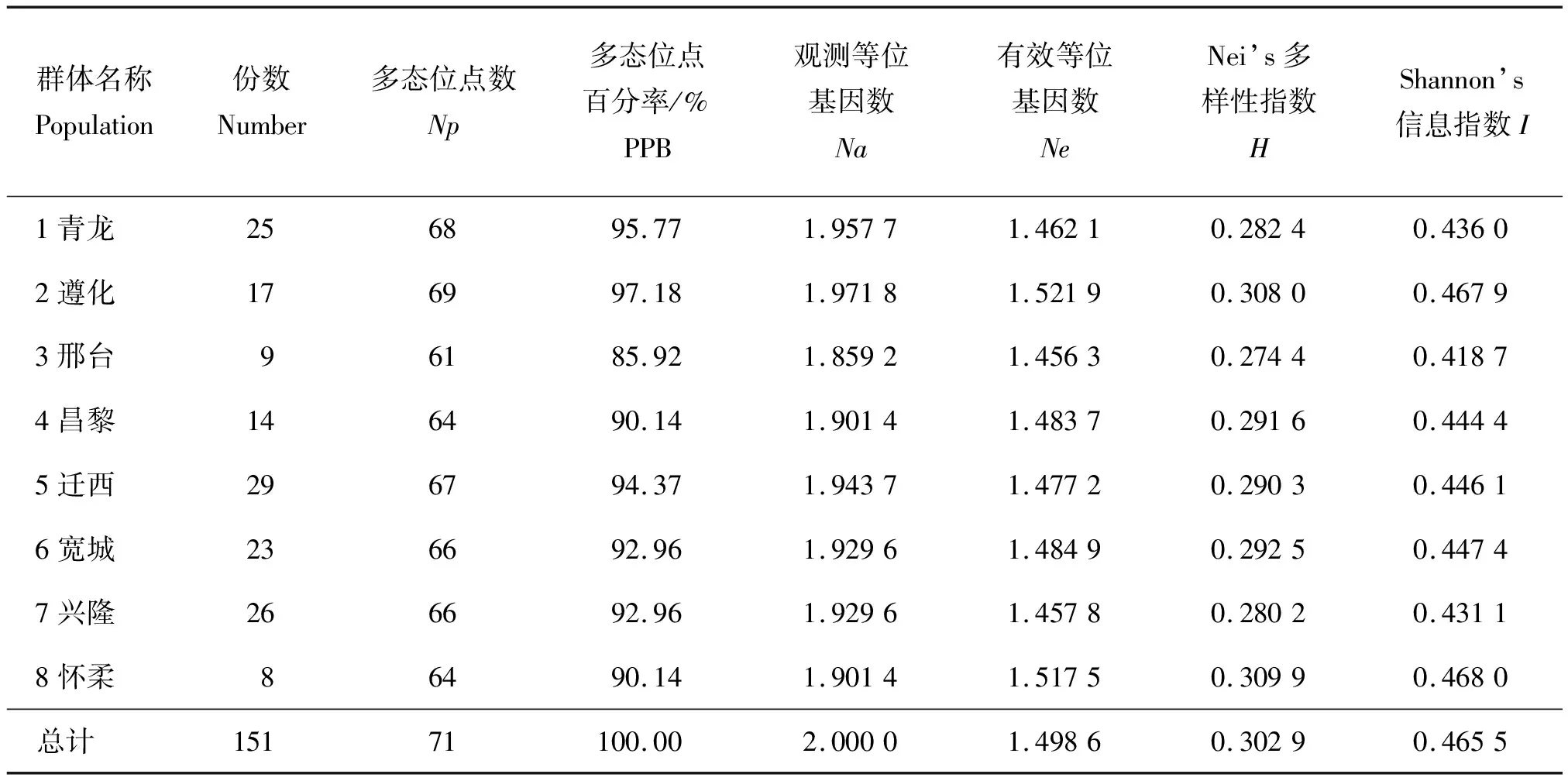
表4 21对SSR引物在8个板栗群体中检测到的遗传多样性参数
2.3 板栗群体遗传分化
供试板栗群体总遗传多样性Ht=0.306 2,群体内遗传多样性Hs=0.291 2,群体间遗传分化系数Gst=0.049 0,即群体间的遗传变异占总变异的4.90%,群体内的遗传变异占总变异的95.10%,表明板栗资源群体内遗传分化大于群体间分化。由于自然或人工选择、随机漂变等原因,板栗群体会出现遗传分化,并且板栗雌雄同株、风媒异花授粉的植物学特性,使得其遗传变异的大多数来自居群内部。板栗群体基因流Nm=9.696 4,说明8个群体间存在丰富的基因交流,防止了因遗传漂变而引起的群体间的遗传分化。
2.4 聚类分析
8个群体间的遗传一致度和遗传距离如表5。遗传一致度和遗传距离分别是从相同和相反方面度量群体间遗传关系的指标。由表5可以看出,各群体间遗传一致度的变化范围为0.959 0(怀柔与邢台群体)~0.987 9(宽城与迁西群体),燕山产区内各群体间的遗传一致度普遍较高,可能与地区之间相互引种有关。遗传距离反映居群间亲缘关系的远近,遗传距离越小,在一定程度上表明二者亲缘关系越近。由表可知,迁西和宽城群体之间的遗传距离最小(为0.012 2),与兴隆种群之间的遗传距离次之(为0.014 9),宽城和兴隆之间遗传距离也很小(为0.016 6),说明迁西、宽城和兴隆群体之间的遗传相似性更趋于一致。邢台和怀柔、邢台和兴隆群体之间的遗传距离相对较大(分别为0.041 9和0.041 0),表明它们之间的亲缘关系相对较远。
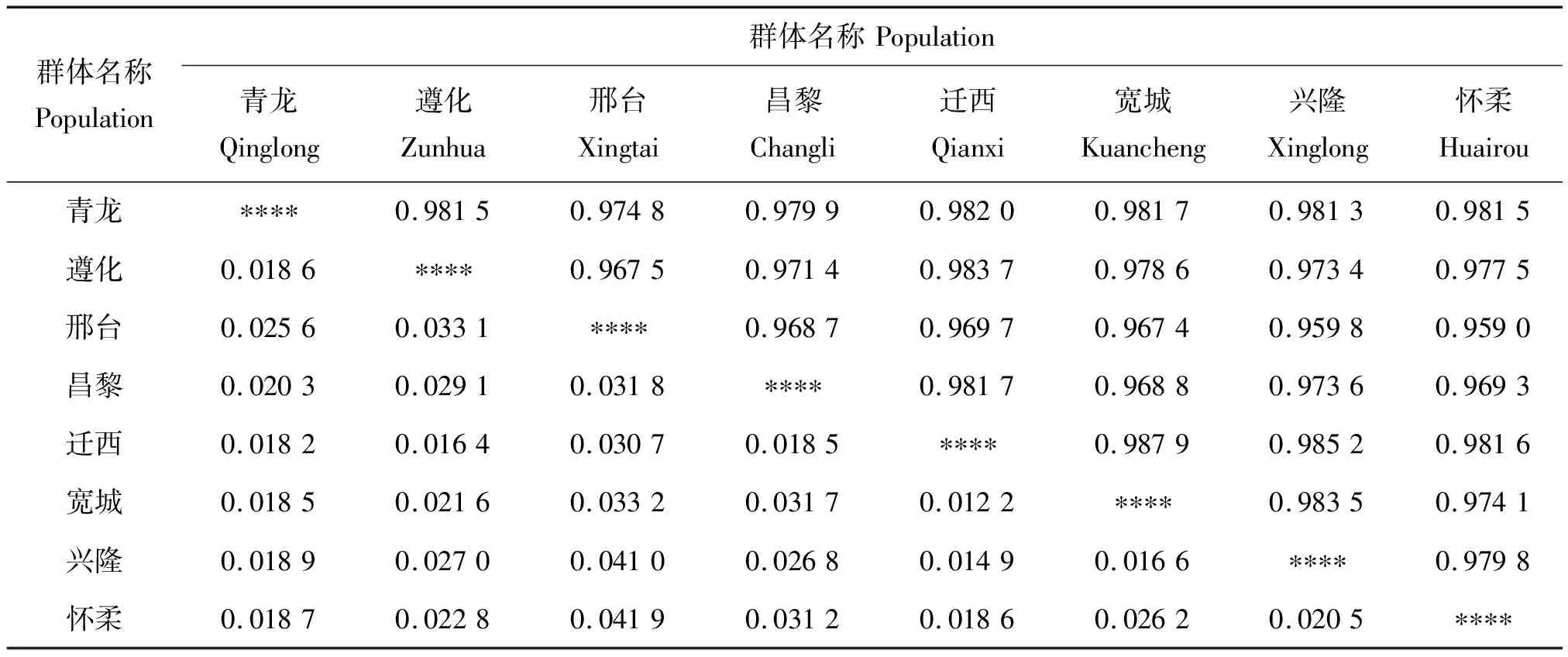
表5 8个群体间的遗传一致度和遗传距离
注:右上三角为遗传一致度,左下三角为遗传距离。
Note: Nei’s genetic identity (the upper right triplet) and genetic distance (the lower left triplet).
根据遗传一致度利用NTSYS(2.10)软件进行聚类(图2)。8个群体间分子遗传相似系数为0.967~0.988,群体的整体相似系数高,说明遗传距离较近。如图2,在遗传相似系数0.977处,可将8个群体分为3个组。其中,第1组包括青龙、迁西、宽城、兴隆、遵化和怀柔群体,而昌黎和邢台群体各为一组。在第1组中,迁西、宽城、兴隆群体遗传距离更近,它们在地理位置上两两相邻,此外,河北省承德市宽城县与秦皇岛市青龙县相邻,唐山市迁西县、承德市兴隆县都与遵化市相邻,它们位于燕山山脉附近,相似的气候条件和地理环境使这些群体的遗传组成趋于一致,遗传距离更近。第2组包括昌黎群体,位于燕山山脉东南方向沿海地区,与第1组遗传距离较远。第3组邢台群体位于太行山脉,地理位置和气候环境与其他两组的群体都不同,所以遗传距离更远,在聚类上也单独一组。
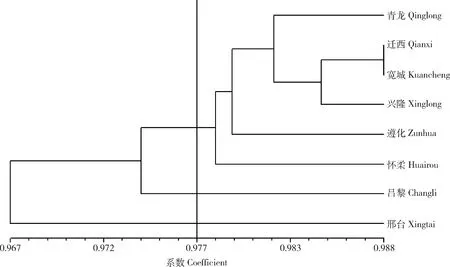
青龙、迁西、宽城、兴隆、遵化、怀柔、昌黎、邢台指来自这些地方的板栗资源组成的群体。
2.5 主坐标分析
根据遗传一致度,作主坐标分析图(图3)。从图3中可以看出8个群体可以分成3个部分,其中,兴隆、迁西、宽城3个群体距离较近,怀柔与兴隆、遵化与迁西、青龙与迁西群体位置靠近,共同组成第1部分。第2部分包括位置稍远的昌黎群体,更远的邢台群体单独组成第3部分。可见,主坐标分析图与聚类图的大体趋势是相一致的。
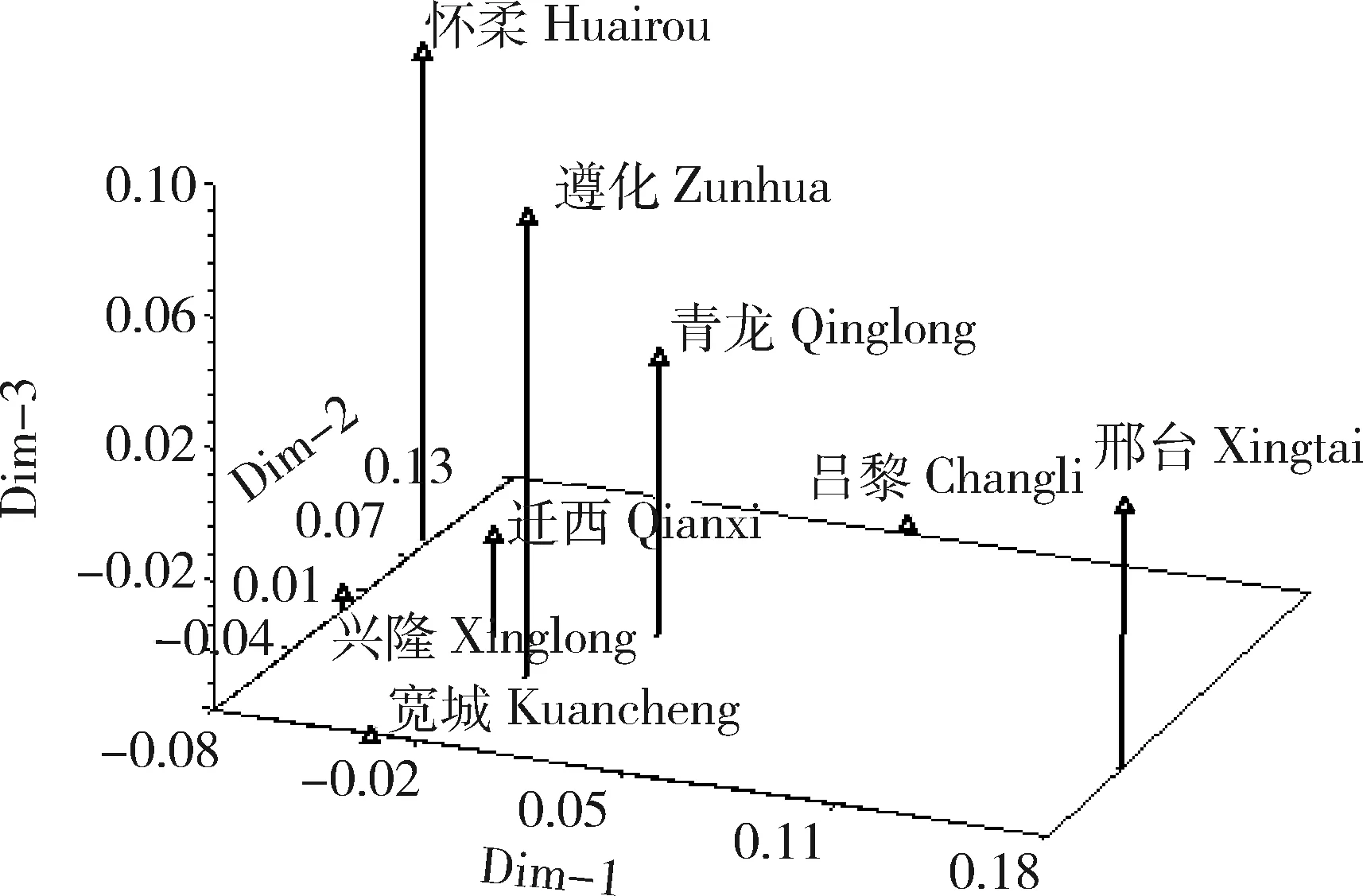
图3 8个板栗群体的主坐标分析三维图
2 讨 论
我国板栗具有较高的遗传多样性水平,已被诸多研究证实[22-23]。王同坤等[24-25]对36份燕山板栗种质资源和8份外来品种进行了AFLP和RAPD分析,结果表明所检测材料中存在着明显的遗传多样性。本研究得到8个板栗群体的有效等位基因数(Ne)、Nei’s 多样性指数(H)、Shannon’s 信息指数(I)分别为1.498 6、0.302 9、0.465 5,表明燕山板栗在居群水平上存在丰富的遗传变异度和较高的遗传多样性水平,这可能与板栗异花授粉的生物学特性有关[26]。本研究统计发现,燕山板栗群体间基因流Nm为9.696 4,表明群体间存在频繁的基因交流,群体遗传多样性丰富,并且种级水平上有95.10%的遗传变异存在于群体内部。有文献报道,异交植物的遗传多样性水平比自交(近交)植物更高[27];并且当基因流Nm值>1时,基因流可以抵制住遗传漂变的影响,防止发生遗传分化[28-29]。板栗具有风媒、虫媒的异交授粉特性,以及人为引种或鸟类迁徙过程中都可能携带种子,减少了地理隔离的影响,使得频繁的基因交流成为可能[3]。
本研究采用SSR分子标记技术对151份种质资源进行遗传多样性分析,共扩增出71个SSR位点,多态性位点百分率为100%,高于王同坤等[24-25]对燕山板栗AFLP分析得到的56.5%多态率和RAPD分析得到的62.9%,造成差异的原因可能有三方面:一是本研究中8个不同群体的151份资源均是来自于燕山各产区由单株实生大树选育而来的地方农家品种、审定品种以及具有特殊性状或潜在价值的实生树材料,本身来源范围广阔、遗传多样性丰富;二是采用人工读取条带的方法进行条带统计,结果较机读更为准确;三是不同的分子标记手段产生的不同试验结果。目前,SSR技术已广泛应于苹果、梨、桃、葡萄、杏、柑橘和樱桃等果树的品种鉴定、遗传图谱构建及遗传多样性研究中[30-36]。房守敏等[37]通过对家蚕RAPD、AFLP和SSR标记的比较分析,认为SSR分子标记由几个碱基对组成核苷酸重复序列,通过检测转录区的序列变异,进一步反映功能上的多样性,能真实说明种质之间的遗传多样性,更适用于种质资源的遗传多样性评价。本研究结果也表明SSR比AFLP和RAPD的信息含量高,更适合用于板栗种质资源的遗传多样性分析。
在本研究所分析的8个板栗群体中,遗传距离为0.012 2~0.041 9,遗传距离较小,遗传相似系数高,说明群体间的亲缘关系较近。供试的8个群体位于我国临近的燕山山脉和太行山山脉周边,所在地理位置相接,相似的自然环境和栽培条件以及频繁的品种交流,都会使这些地区的群体在遗传组成上大体趋于一致;此外,长期的人工干预选择,也会使燕山板栗的遗传基础趋于单一[24]。聚类图和主坐标分析图显示8个板栗群体被分成三类,燕山板栗主产区青龙、迁西、宽城、兴隆、遵化和怀柔群体划为1类,而处于燕山区的昌黎群体和处于太行山区邢台群体各为1类,其原因可能是河北省秦皇岛市昌黎县虽处于燕山山脉,但其位于燕山山脉东南方向沿海地区,气候类型和环境条件与燕山山脉北部燕山板栗主产区县市有明显差异,导致该群体资源在长期的环境适应和进化过程中形成了适应当地环境的一些基因性状;而邢台群体位于太行山山脉一带,与供试燕山板栗群体在地理位置上有明显距离,空间距离上的阻隔是造成这一群体和燕山板栗群体具有明显遗传上的差异的主要原因。
