人类活动对山区溪流鱼类多样性的影响
2020-04-09韦宏军
韦宏军


摘 要:确定人类活动对溪流鱼类多样性的影响及其作用机理,是保护溪流鱼类多样性保护的科学基础。该研究选取了安徽省九龙峰自然保护区境内的殷溪河及其周边的焦村河为研究水域,分别于2018年2、5、9和11月4次取样,比较分析了2条溪流的局域栖息地、鱼类群落结构及其多样性的异同,进而探讨了人类活动对溪流鱼类多样性的影响及其机理。结果显示,相对于殷溪河而言,焦村河在丰水期(5月和9月)栖息地的空间异质性略高,而在枯水期(2月和11月)的栖息地空间异质性相对较低。尽管2条溪流间的鱼类物种数和个体数无显著差异,但焦村河的β多样性显著高于殷溪河;同殷溪河相比,光唇鱼、原缨口鳅等亲流性敏感鱼类的数量明显下降,而泥鳅、麦穗鱼等缓流/静水性耐受鱼类的数量明显增多。说明城镇化、水坝建设等人类活动可破坏河流生态系统原有的栖息地环境,导致溪流鱼类物种群落多样性发生变化。
关键词:溪流鱼类;群落结构;α和β多样性;人为干扰
中图分类号 Q958.8;X824文献标识码 A文章编号 1007-7731(2020)05-0081-05
溪流生态系统具有极高的空间异质性,同大型河流相比,河源溪流具栖息地条件简单且环境容纳量小等特点,故溪流鱼类的α多样性(局域群落的物种多寡)较低[1]。相比于生物群落α多样性而言,β多样性度量则是群落间的种类组成变化幅度[2]。面对环境变化或人类干扰,群落间的物种组成变化表现为物种获得或物种丢失;而局域群落多样性(如物种数)的变化则取决于丢失物种和获得物种的相对多寡[2]。受人类活动的影响,全球的生物多样性正在以前所未有的速度发生变化:广布物种(cosmopolitan species)不断扩散和被引种、土著物种(endemic species)日益濒危和灭绝;广布物种对土著物种的取代过程降低了生物群落的特质并提高了群落间的相似性,进而导致“生物同质化”(biotic homogenization)[3]。
当前,人类活动导致的栖息地碎化、外来种入侵等因素所导致的生物多样性丧失已经成为当今环境问题的焦点[4,5]。溪流生态系统中的人类干扰形式多样,如:水坝切断了溪流原有的连通性,改变溪流的水源补给和营养输入,严重破坏了原有的栖息地条件[6,7];城镇化等土地利用使得水体沉积物增加且随地表径流汇入溪流系统,并降低了溪流鱼类的生物完整性及物种数量,导致很多地方性敏感物种减少甚至局域性灭绝[8-9]。
在溪流鱼类多样性持续受威胁的背景下,深入认识人为干扰引起的鱼类多样性变化及其机制对于生物多样性保护至关重要。因此,本研究以安徽九龙峰自然保护区境内的殷溪河与周边的焦村河为研究区域,基于全年4个季度的野外调查数据,比较分析了2条溪流中的局域栖息地和鱼类多样性的异同,进而探讨了人类活动对溪流鱼类多样性的影响及其机理,以期为研究区域所在的黄山山区溪流鱼类多样性的保护积累基础资料。
1 材料与方法
1.1 研究区域 本研究选取殷溪河和焦村河作为研究区域,这2条河流位置接近、大小相似,但受到的人为干扰影响不同。殷溪河发源于安徽省黄山市东部地区的九龙峰自然保护区,自南向北经舒溪河和太平湖汇入青弋江,因而是青弋江流域的河源支流之一;焦村河毗邻殷溪河,自南向北经穰溪河和太平湖汇入青弋江,因而也是青弋江流域的河源支流。地处我国亚热带季风气候区,其气温和降水的季节变化明显:累年各月平均气温范围4.2℃~27.9℃;年平均降水量1800mm,但约80%的降水集中在4—9月。
1.2 野外取样 本研究针对殷溪河和焦村河,自上而下分别设置3个样点(图1),每样点调查河段长度统一为100m。2018年,按季度(2月、5月、9月和11月)对上述设置样点进行野外调查和取样。以背式电鱼器(CWB-2000P,输入12V,输出250V)作为渔具,取样时间统一为30min。对野外采样获得的新鲜渔获物鉴定至种,统计并记录渔获物的物种数和个体数;由于检索工具的限制,吻鰕虎鱼属鱼类仅鉴定至属的水平。疑难种以8%甲醛固定,带回实验室进行室内鉴定,其余活体标本放回采集地。
针对每个调查样点,现场测量6类局域栖息地因子。将采样水域分为3个等距的截面,且每截面分为3个等距的截點,于每截面使用测距仪测量水宽(WW,m);于每截点测量水深(WD,cm),并在60%水深处便携式流速仪(FP111,美国)测量流速(CV,m/s);同时使用YSI便携式水文仪现场测度水温(WT,℃)、电导率(Do,mS/cm)和溶氧(DO,mg/L)。
1.3 多样性指数计算 以每样点的物种数和个体数代表α多样性。根据两两样点间的群落组成,计算其β多样性(βs?r),并参照张东的方法,将其分解为周转(βsim)和嵌套(βnes)2个成分。以某样点与其他各样点的β多样性平均值代表该样点的β多样性大小。上述β多样性指数及其周转和嵌套成分的计算在R软件中进行。
1.4 数据分析 根据某物种的出现样点数与总样点数的比值确定该物种的出现频率(Fi),Fi≥40%、10%≤Fi<40%、Fi<10%的物种分别被视为常见种、偶见种和稀有种;Fi=(Si/S)×100,式中,Si为物种i的出现样点数,S为全部样点数。根据某物种的个体数量与该样点的全部渔获物数量的比值确定相对多度(Pi):Pi=(Ni/N)×100,式中,Ni为物种i的个体数,N为全部物种的个体数。
运用多元统计解析受试组和对照组的栖息地条件变异情况,且根据标准化的栖息地变量计算相应的欧式距离矩阵。基于两类样点栖息地变量的欧氏距离,运用主坐标分析(PCoA)获取各调查样点的散点分布情况,根据样点间的差异性将各样点进行欧式距离排序;并运用基于距离的冗余分析(dbRDA)解析栖息地变量的相对重要性及其解释百分比。运用主坐标典型分析(CAP)检验两类样点间栖息地条件的平均差异。最后,运用置换多元分散分析(PERMDISP)检验两类样点间栖息地条件的异质性,可进一步运用方差分析F-统计来比较样点间的差异显著性。上述分析分别考虑整体情况(忽略季节因素)和各季节内的受试组和对照组的栖息地条件差异。
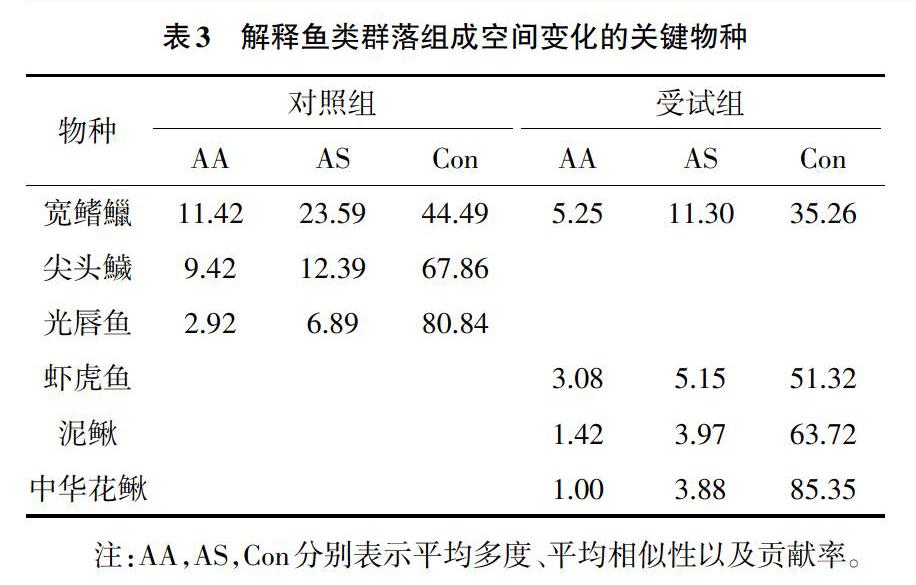
利用Primer5.0软件,运用双因素交叉相似性分析(Two-way crossed ANOSIM) 检验栖息地类型和季节对鱼类群落结构的影响,根据R值确定鱼类群落的分离程度:R>0.75,群落完全分离;0.5 利用SPSS 22.0软件,运用双因素方差分析分别检验不同栖息地类型和季节对局域环境变量和鱼类群落多样性的影响,5个多样性指数独立进行(物种数、个体数、βsor、βnes、βsim)。上述统计分析中,原始数据均经log(X+1)转换后用于统计分析,以满足数据的正态分布和方差齐性要求。 2 结果与分析 2.1 鱼类物种的组成 由表1可知,共采集鱼类673尾,隶属3目6科15种。鲤形目物种数最多,占全部物种的80%。各样点的平均物种数为4.54。殷溪河中共采集鱼类10种,共471尾,焦村河中鱼类13种,共计202尾。中华花鳅、泥鳅、马口鱼、麦穗鱼、高体鳑鲏仅见于焦村河,台湾白甲鱼仅出现于殷溪河。宽鳍鱲的出现频率和相对多度均最大,分别为89.50%和29.72%,虾虎鱼次之,麦穗鱼出现次数和数量最小,分别为4.17%和0.15%。 2.2 局域栖息地条件 运用双因素方差分析检验溪流和季节对各局域栖息地变量的影响,结果显示,溶氧、pH、水温和流速的季节变化显著(P<0.05,F=25.98,溶氧;F=17.59,pH;F=51.98,水温;F=4.845,流速),而仅溶氧存在显著的溪流间差异(P<0.05,F=12.28)。此外,pH和水温受到溪流和季节显著的交互影响(P<0.05,F=3.54,pH;F=4.04,水温)。 分别考虑4个季节内受试组和对照组的局域栖息地条件差异,发现dbRDA的第1轴和第2轴分别解释局域栖息地变异在各季节中存在差异(48.1%和28.9%,2月;47.6%和25.1%,5月;48.2%和39.3%,9月;52.9%和23.4%,11月)(图2)。此外,与dbRDA 2个轴的相关变量在各季节内也存在差异(表2)。CAP分析显示,2类生境间的栖息地同质性在各季节内均无显著性差异(P>0.05,trace=0.66(2月),trace=0.25(5月),trace=0.97(9月),trace=0.99(11月));同时,PERMDISP分析也显示,这两类生境间的栖息地同质性也无显著性差异(P>0.05,F=0.26(2月),F=5.82(5月),F=1.00(9月),F=3.45(11月)),但相对对照组而言,2月和11月的受试组各样点间的平均欧式距离略低(2月:1.87±0.24(受试组),2.02±0.16(对照组);11月:1.60±0.18(受试组),2.31±0.34(对照组)),而5月和9月的受试组各样点间的平均欧式距离略高(5月:2.48±0.11(受试组),1.97±0.18(对照组);9月:2.16±0.40(受试组),1.72±0.20(对照组))。 2.3 鱼类群落结构的时空变化 双因素交叉相似性分析结果显示,对照组和受试组鱼类群落存在明显重叠但仍可分离(Global R=0.269;P<0.05);但两者均不受季节因素的影响(对照组:Global R=0.108,P>0.05;受试组:Global R =0.024,P>0.05)。根據鱼类群落非度量多维标度排序(NMS)双标图,同样也得到以上结果(图3)。运用相似性百分比分析确定造成其群落组成差异的关键物种。结果显示,维持对照组群落组成相似性的关键物种为宽鳍鱲、尖头鱥、光唇鱼,而受试组的关键种则是宽鳍鱲、虾虎鱼、泥鳅和中华花鳅等(表3)。 2.4 鱼类群落多样性的时空变化 双因素方差分析显示,α多样性和β多样性的季节变化均无显著性(P>0.05);就空间(溪流间)变化而言,尽管鱼类物种数和个体数不存在显著的溪流间差异(P>0.05),但β多样性及其嵌套和周转成分都存在显著的溪流间变化(P<0.05):同殷溪河相比,焦村河中的鱼类β多样性及其周转和嵌套成分都显著高于殷溪河(图4);各个多样性指数均不受季节和溪流的显著性交互影响(P>0.05)。 3 讨论 本研究的2条大小相似、位置接近的河源溪流——焦村河和殷溪河,前者在保护区外,其局域栖息地环境受到的人为干扰程度相对较大,而后者则位于保护区内,其栖息地条件接近自然状态。已有报道表明,诸如城镇化发展、水利建设等人类活动,严重破坏河源溪流的栖息地条件[4,6]。类似地,本研究发现,焦村河和殷溪河的溶氧量具有显著性差异,同时pH和水温也受到季节和溪流的联合影响。人类活动不仅能改变局域尺度上的栖息地特征,还能通过改变水文情势、水流流态等栖息地条件的空间异质性而发生栖息地同质化或异质化[3,10]。尽管各季节内焦村河和殷溪河局域栖息地条件的平均差异和空间异质性并无显著性,但在丰水期(5月和9月)时,相对于殷溪河而言,焦村河栖息地变量的平均欧氏距离略高,其空间异质性相对较大,即发生轻微的栖息地异质化;在枯水期(2月和11月)时,焦村河栖息地变量的平均欧氏距离略低,即发生轻微的栖息地同质化现象。本文的焦村河受到较重的人为干扰,存在不同的土地利用方式,诸如城镇发展、农业用地、建设水坝等,导致该区域的空间栖息地异质性相对较高,这与已有的报道结果类似[11]。因此,在丰水期时,焦村河和殷溪河的水流流态相似,而焦村河受到人为活动的影响,其局域栖息地条件的空间异质性较大,易发生栖息地异质化现象。反之,2条溪流在枯水期的水流流态却具有差异,主要因为焦村河建设大量具有蓄水作用的低头坝,样点间的栖息地条件差异性较小,而殷溪河的水文情势接近自然状态,枯水期容易断流增加样点间的栖息地条件空间异质性,因此焦村河易发生栖息地同质化。殷溪河和焦村河中鱼类群落结构及其β多样性存在显著差异。就鱼类群落结构而言,尽管两条河流的鱼类群落结构存在部分重叠,但具有显著差异。经相似性百分比分析发现,殷溪河其维持群落相似性的关键物种为宽鳍鱲、光唇鱼、原缨口鳅等,且这些物种的相对多度较高;而焦村河中的关键物种则多为泥鳅、中华花鳅等。就群落多样性而言,虽然2条河流中鱼类的物种数和个体数无显著差异,但两者的β多样性差异显著。由于本研究所选取的2条河流地理位置、河流大小等具有一致性,因此2条河流间鱼类群落结构及其多样性的差异应体现在它们对当前人为活动的响应。殷溪河位于安徽省省级自然保护区内,其干流的植被覆盖相对完好,农业面积较小、人为活动较少;而焦村河中游有焦村,人口居住密度大,农业占地面积较大。
已有大量研究表明,土地利用[12]、城镇化[13]、水坝建设[6]等人类活动对溪流生态系统的栖息地条件及生物群落结构有着严重影响和危害。比如:人为干扰可降低溪流的栖息地多样性和复杂性,破坏溪流中原有的“急滩-深潭”交替格局,扰乱了溪流中原有的水流流态和水文,减少溪流与沿岸陆地的临界面[12,14]。栖息地条件的变化对溪流鱼类的物种组成及其多样性同样有深远影响,栖息地适合度的下降可引起溪流中原有的急流鱼类物种及其数量的减少甚至是局域性灭绝[15];与此同时,溪流中的栖息地变化还可引起一些外来入侵种和本地入侵种数量的增加。有研究表明,青弋江河源溪流中的低头坝对鱼类群落的影响,同远离水坝的对照点相比,坝上蓄水区的鱼类多样性显著下降且物种组成也明显变化—宽鳍鱲、光唇鱼等急流鱼类数量减少,而麦穗鱼、高体鳑鲏等缓流鱼类数量增多[6]。本研究结果中,同殷溪河相比,焦村河中出现泥鳅、麦穗鱼、马口鱼等喜缓流鱼类。而尖头岁、台湾白甲鱼均仅出现在对照组殷溪河中。两者鱼类群落结构的差异体现了河流生态系统对周边人类活动的影响;尖头岁、台湾白甲鱼为高海拔喜冷水的物种,光唇鱼、原缨口鳅等物种的生态位宽度也较为狭窄,均多栖息于山涧溪流中,属于典型的急流鱼类,对环境变化即为敏感;而泥鳅、麦穗鱼、马口鱼等物种往往多适应大型河流的缓流环境或湖泊、水库等静水环境,不仅分布范围较广且生态耐受性较高。因此,本研究结果进一步证明了人类活动导致溪流中敏感物种减少、广布耐受物种增多。有报道表明,人为活动显著改变溪流鱼类群落α和β多样性,人为干扰在溪流下游地区较为强烈,导致鱼类群落β多样性上升[16]。本研究发现,虽然殷溪河和焦村河间的鱼类α多样性(物种数和个体数)无显著差异,但焦村河中鱼类β多样性显著高于殷溪河中。有文献报道,人为活动显著改变溪流鱼类群落α和β多样性,人为干扰在溪流下游地区较为强烈,导致鱼类群落β多样性上升。引起这一变化的原因,可能是由于人为干扰导致原有的特有种减少,而泛化种增多[16]。本研究中焦村河的β多样性显著高于殷溪河,由于焦村河坡度较小且无高山瀑布等明显的地理障碍,下游的鱼类很容易入侵至中上游地区。
参考文献
[1]王文剑,储玲,司春.秋浦河源国家湿地公园溪流鱼类群落的时空格局[J].动物学研究,2013,34(4):417~428.
[2]张东,宛凤英,储玲,等.青弋江鱼类分类群和功能群的α和β多样性纵向梯度格局[J].生物多样性,2018,26(1):1-13.
[3]卜倩婷,李献,朱仁,等.低头坝驱动山区溪流局域栖息地和鱼类群落的同质化[J].生物多样性,2017,25(8),830-839.
[4]储玲,王文剑,闫莉莉,等.黄山陈村水库上游河源溪流的鱼类群落及其纵向梯度格局[J].生态学报,2015,35(3),900-910.
[5]Zhu R,Li Q,Wang WJ,et al. Effects of local,river-network and catchment factors on fish assemblages in the headwater streams of the Xin′an basin,China.[J]. Journal of Freshwater Ecology,2017,32:300-313
[6]Yan Y,Wang H,Zhu R,et al. Influences of low-head dams on the fish assemblages in the headwater streams of the Qingyi watershed,china.[J] Environmental Biology of Fishes,2013,96:495-506.
[7]李强,张东,宛凤英,等.溪流鱼类群落对低水头坝的大小及功能的响应——以皖南山区河源溪流为例[J].水生生物学报,2018,42(5):965-974.
[8]钱红,严云志,储玲,等.巢湖流域河流鱼类群落的时空分布[J].长江流域资源与环境,2016,25(2):257-264.
[9]李其芳,严云志,储玲,等.太湖流域河流鱼类群落的时空分布[J].湖泊科学,2016,28(6):1371-1380.
[10]Liu Y,Wang Y,Zhu Q,et al. Effects of low-head dams on fish assemblages in subtropical streams:Context dependence on species category and data type.[J]. River Research and Applications,2019,35:396-404.
[11]Thompson K,Austin KC,Smith RM,et al. Urban domestic gardens(I):putting small-scale plant diversity in context[J].J Veg Sci,2003,14:71–78.
[12]Allan JD. Landscapes and riverscapes:the influence of land use on stream ecosystems. Annual Review of Ecology [J] .Evolution and Systematics,2004,35:257–284.
[13]Wang L,Lyons J,Kanehl P,et al. Impacts of urbanization on stream habitat and fish across multiple spatial scales [J]. Environmental Management,28(2),2001,255-266.
[14]Poff NL,Hart DD. How dams vary and why it matters for the emerging science of dam removal[J]. BioScience,2002,52:659–668.
[15]Yan YZ,Xiang XY,Chu L,et al. Influences of local habitat and stream spatial position on fish assemblages in a dammed watershed,the Qingyi Stream,China[J]. Ecology of Freshwater Fish,2011,20(2):199-208.
[16]Li YR,Tao J,Chu L,et al. Effects of anthropogenic disturbances on α and β diversity of fish assemblages and their longitudinal patterns in subtropical streams,China[J]. Ecology of Freshwater Fish,2018,27:433–441.
(責编:张 丽)
