3个茶用菊花品种应对碱胁迫的生理响应及耐碱性评价
2020-03-24杨欢李锦馨姚爱彬王晨轩
杨欢 李锦馨 姚爱彬 王晨轩


摘要:【目的】探究堿胁迫对不同茶用菊花品种花芽分化期生长和生理生化指标的影响,并确定菊花耐碱评价和筛选的主要指标,为菊花抗碱新品种选育和鉴定提供科学依据。【方法】用添加不同浓度(0、20、40、60 mmol/L)Na2CO3溶液的Hoagland营养液对3个菊花品种进行碱胁迫处理,测定不同处理条件下夏妆、祁黄菊和雪映霞光3个茶用菊花新品种的生长指标及生理生化指标,比较分析品种间及品种内不同胁迫浓度处理随胁迫周期的变化规律,以确定菊花品种的耐碱性强弱。【结果】随着碱胁迫时间的延长,3个菊花品种叶片的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性及脯氨酸(Pro)和可溶性糖(SS)含量均呈先增加后降低的变化趋势,丙二醛(MDA)含量、相对电导率(RE)则呈现逐渐增加的变化趋势;而叶绿素相对含量(SPAD)及叶片相对含水量(WC)呈逐渐降低的变化趋势。运用主成分分析,碱胁迫下第1、第2主成分累计贡献率达92.853%,提炼出SOD活性、POD活性、SS含量和Pro含量4个生理指标作为耐碱评价的关键指标。综合公因子载荷矩阵耐碱性评价和隶属函数值耐碱性评价分析得出,3个菊花品种耐碱性为雪映霞光>祁黄菊>夏妆。【结论】在碱胁迫下,3个菊花品种能在一定程度上维持细胞代谢平衡及体内化学物质的合成积累平衡,并进行活性氧的清除,以缓解并适应碱胁迫对其的伤害。但菊花的这种自卫能力有一定的局限性,当超过承受范围时,各品种菊花呈现不同程度碱害症状,雪映霞光更能迅速响应胁迫,其碱害症状明显小于祁黄菊和夏妆。
关键词: 菊花;碱胁迫;生理生化指标;耐碱性评价
中图分类号: S682.11 文献标志码: A 文章编号:2095-1191(2020)12-2993-11
Abstract:【Objective】The effects of alkali stress on the growth and physiological and biochemical indexes of diffe-rent tea chrysanthemum varieties during the flower bud differentiation period were explored, and the main indicators for evaluation and screening of chrysanthemum alkali resistance were determined to provide scientific basis for the selection and identification of new chrysanthemum alkali resistant varieties. 【Method】Three chrysanthemum varieties were subjec-ted to alkaline stress treatment with Hoagland nutrient solution supplemented with different concentrations(0,20,40, 60 mmol/L) of Na2CO3 solution, and the three varieties of Xiazhuang, Qihuangju, and Xueyingxiaguang were tested under different treatment conditions. The growth indicators and physiological and biochemical indicators of the three chrysanthemum varieties were detected, compared and analyzed the variation regulations between varieties and within varieties in different stress concentration treatments with stress cycles to determine the alkali tolerance of chrysanthemum varieties. 【Result】With the prolongation of alkaline stress, the superoxide dismutase(SOD), peroxidase(POD) and catalase (CAT) activities, proline(Pro) and soluble sugar(SS) content first increased and then decreased. Malondialdehyde(MDA) and relative conductivity(RE) showed a gradual increase trend; while relative chlorophyll content(SPAD) and relative leaf water content(WC) showed gradual decreasing trend of change. Using principal component analysis, the cumulative contribution rate of the first and second principal components under alkali stress reached 92.853%, and four physiological indicators of SOD activity, POD activity, SS content, and Pro content were extracted as key indicators for alkali resistance evaluation. According to the evaluation of alkali resistance of common factor loading matrix and the evaluation of membership function value, it was concluded that the alkali resistance of the three chrysanthemum varieties was Xueyingxiaguang>Qihuangju>Xiazhuang. 【Conclusion】Under alkali stress, the three chrysanthemum varieties can maintain the ba-lance of cell metabolism and the synthesis and accumulation of chemical substances in the body to a certain extent, and carry out the elimination of reactive oxygen species to alleviate and adapt to the damage of alkali stress. However, the self-defense ability of chrysanthemum has certain limitations. When it exceeds the tolerance range, various chrysanthemum varieties show different degrees of alkali damage symptoms. Xueyingxiaguang can respond more quickly to the stress, and its alkali damage symptoms are significantly less than that of Qihuangju and Xiazhuang.
Key words: chrysanthemum; alkali stress; physiological and biochemical indexes; alkali tolerance evaluation
Foundation item: Science and Technology Department Key Research and Development Project of Ningxia Hui Autonomous Region(2019BBF02029)
0 引言
【研究意义】菊花[Dendranthema morifolium(Ramat) Tzvelev]是菊科菊属多年生宿根草本植物(管志勇等,2010),是我国十大名花之一,名列世界著名的四大切花(李荣华等,2012)。菊花种质资源极其丰富,在我国乃至世界花卉市场中一直占有重要的主导地位(孙静,2012)。其中,茶用菊既具有观赏价值,又因其多重保健功能而具有巨大的经济效益,推广应用前景广阔。我国碱土和碱化土壤的形成大部分与土壤中碳酸盐的累积有关,因而碱化度普遍较高,严重的盐碱土壤地区植物几乎不能生存。研究菊花的耐碱性并筛选出耐碱菊花品种,对盐碱地的菊花开发利用具有重要意义。【前人研究进展】盐碱胁迫及盐碱地利用已成为研究热点。除离子毒害和渗透胁迫是碱胁迫主要迫害因素外,还包括高pH对植物的胁迫(Wang et al.,2015)。与盐胁迫相比,碱胁迫对植物离子毒性的综合作用危害更大(刘杰,2011),其对植物不同层次(细胞、组织和器官等)的养分吸收,以及有机酸,离子稳态和新陈代谢的平衡均有迫害作用,更严重地会阻碍植物的生长(Liu et al.,2018)。高强度碱胁迫对光合速率的抑制程度大于盐胁迫(Gong et al.,2013)。在碱胁迫下,以脯氨酸(Pro)对植物进行叶面喷施,可以刺激植物茎、叶和根部的生长,提高植物干重和鲜重,从而提高光合作用的速率(Butt et al.,2016)。在碱胁迫过程中发生氧化胁迫反应,植物活性氧(ROS)增加,由于细胞受损,相对电解质渗漏物(REL)含量和丙二醛(MDA)含量增加,植物依靠抗氧化酶系统来清除ROS,以减少碱性胁迫对植物的伤害(楚乐乐等,2019)。不同植物中不同抗氧化酶的活性变化不同(Zhao et al.,2016)。目前,关于水稻(赵海新,2012)、油菜(陶顺仙等,2019)、辣椒(张海英,2019)等作物的相关研究表明,随着碱胁迫浓度的增加,植物抗氧化酶活性呈先升后降的变化趋势。【本研究切入点】关于菊花的研究主要集中在花期调控及优良品种选育上,针对抗逆性方面的研究主要集中在水分和温度胁迫方面,碱胁迫方面的研究较少。【拟解决的关键问题】对碱胁迫下夏妆、祁黄菊和雪映霞光3个茶用菊花新品种叶片的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性及Pro和可溶性糖(SS)含量等9项生理指标及株高、冠幅2项生长指标进行测定、分析研究和评价,运用主成分分析法筛选出耐碱评价的关键性生理指标;综合各项指标分析讨论及采用隶属函数分析、公因子载荷矩阵分析3个品种的耐碱性强弱,筛选出耐碱性品种,为盐碱地地区茶用菊花品种的推广提供理论依据。
1 材料与方法
1. 1 试验材料
供试材料为当年生6叶龄观赏兼茶用菊花夏妆、祁黄菊和雪映霞光扦插苗。其中,夏妆和雪映霞光为培育的新品种,祁黄菊为传统品种。所有材料均来自中国农业大学。前期对同批22个品种做盐碱胁迫预试验,结果表明这3个品种在预试验过程中长势相对较好。
1. 2 试验方法
1. 2. 1 试验设计 试验于2019年6—11月进行。2019年6月初,选择生长一致且生长健壮的3个菊花品种扦插苗,栽植于上口径20 cm、下口径18 cm、高25 cm的塑料花盆内,盆底配套托盘。栽培基质由蛭石、草炭和珍珠岩按1∶1∶1的比例配制而成,养护条件一致,以清水灌溉。于8月上旬花芽分化期选择生长状态一致的植株进行碱胁迫处理。以Na2CO3进行碱胁迫模拟,设置分别添加20、40和60 mmol/L Na2CO3 3個碱浓度的Hoagland培养液,以0 mmol/L Na2CO3碱液的Hoagland培养液为对照组(CK),每个处理组重复3次,每个重复5盆。每天上午8:00—9:00时进行胁迫灌溉,每个胁迫组灌溉100 mL相应浓度的碱性营养液(预试验确定100 mL完全灌溉了基质,但渗到托盘的溶液不会流出)。为避免盐碱冲击的影响,胁迫组从最低碱度梯度进行浇灌,以后每次浇灌时增大一个碱度梯度,直到达到设定碱浓度,达到设定浓度当天为胁迫0 d。分别在达到胁迫浓度的第0 d(第1周)、7 d(第2周)、14 d(第3周)、21 d(第4周)和28 d(第5周)选取植物上部第4~6片完全展开叶,采样用于测定各项生理指标。
1. 2. 2 指标测定及方法 叶片含水量(WC)采取浸泡法(高俊凤,2006)测定;MDA含量采取硫代巴比妥酸法(张治安等,2004)测定;SS含量采取蒽酮比色法(郝建军等,2007)测定;Pro含量采取水合茚三酮法(郝建军等,2007)测定。使用SPAD-502Plus便携式叶绿素仪测定叶片的叶绿素含量(SPAD),相对电导率(RE)采用雷磁DDS-307进行测定。SOD活性用氮蓝四唑法(王学奎,2006)测定;POD活性采用愈创木酚显色法(曹翠玲和麻鹏达,2016)测定;CAT活性参考李合生(2000)的方法进行测定。
1. 3 统计分析
用SPSS 23.0进行数据统计分析;用Origin 2018制图;利用Duncans新复极差法进行差异显著性检验。
2 结果与分析
2. 1 碱胁迫对菊花生长指标的影响
当植物遭遇盐碱迫害时,外在形态的变化最为直观,如图1所示,经过28 d的碱胁迫处理,夏妆、祁黄菊和雪映霞光3个菊花品种植株的株高和冠幅变化十分明显。由图2可看出,随着碱胁迫浓度的升高,3个菊花品种的株高和冠幅均呈逐渐减少的变化趋势,且各浓度处理间达显著差异(P<0.05,下同)。
由图2-A可看出,在碱胁迫浓度为20 mmol/L时,夏妆、祁黄菊和雪映霞光的株高分别比CK低9.87%、6.74%和3.44%,其中,夏妆的株高降幅最大,为3.0 cm,雪映霞光的株高降幅最小,为1.0 cm;碱胁迫浓度升至40 mmol/L时,夏妆、祁黄菊和雪映霞光的株高分别比CK低17.43%、12.36%和6.53%,其中,夏妆的株高降幅最大,为5.3 cm,雪映霞光的株高降幅最小,为1.9 cm;碱胁迫浓度为60 mmol/L时,夏妆、祁黄菊和雪映霞光的株高分别比CK低22.37%、14.23%和9.62%,其中,夏妆的株高降幅最大,为6.8 cm,雪映霞光的株高降幅最小,为2.8 cm。由此可见,Na2CO3浓度越高,对菊花株高的抑制作用越明显。
由图2-B可看出,在碱胁迫浓度为20 mmol/L时,夏妆、祁黄菊和雪映霞光的冠幅分别比CK减少9.46%、3.78%和2.17%,其中,夏妆的冠幅降幅最大,为2.1 cm,雪映霞光的株高相比CK差值最小,仅0.6 cm;碱胁迫浓度升至40 mmol/L时,夏妆、祁黄菊和雪映霞光的冠幅分别比CK减少13.51%、7.57%和3.26%,其中,夏妆的冠幅降幅最大,为3.0 cm,雪映霞光的冠幅降幅最小,为0.9 cm;在碱浓度胁迫达60 mmol/L时,夏妆的冠幅减少16.67%,最大冠幅差3.7 cm,祁黄菊和雪映霞光分别减少9.19%和3.99%。可见,3个菊花品种中夏妆对碱胁迫的响应程度最大。综上所述,碱胁迫对菊花的株高和冠幅有明显影响。
2. 2 碱胁迫对菊花叶片可溶性渗透物含量的影响
2. 2. 1 Pro含量 由图3可看出,随着碱胁迫时间的延长,3个菊花品种的叶片Pro含量均呈先增加后减少的变化趋势。其中,夏妆在第3周时达最高值,而祁黄菊和雪映霞光在第4周时达最高值,此时胁迫浓度均为40 mmol/L,说明夏妆比另外2个品种对碱胁迫更敏感。胁迫第5周时,与CK相比,夏妆、祁黄菊和雪映霞光的叶片Pro含量在胁迫浓度为20 mmol/L时分别增加129.42%、43.43%和86.71%,在胁迫浓度为40 mmol/L时分别增加160.89%、82.39%和116.87%,在胁迫浓度为60 mmol/L时分别增加149.18%、72.38%和99.03%。可见,在低浓度胁迫下,菊花叶片Pro含量随胁迫浓度增加而增加;而当胁迫浓度过高时,Pro含量反而减少,其中,夏妆的叶片Pro含量增幅最小,雪映霞光的叶片Pro含量增幅最大。综上所述,在碱胁迫下菊花叶片Pro起着重要的渗透调节作用,细胞Pro含量的增加不仅维持了细胞的膨压,同时也使保护酶和膜系统免受碱胁迫毒害。
2. 2. 2 SS含量 由图4可知,随着胁迫时间的延长,3个菊花品种的叶片SS含量也呈先增后减的变化趋势,胁迫4周时达最高值,此时胁迫浓度为40 mmol/L;且不同胁迫浓度下叶片SS含量差异显著。胁迫第4周时,与CK相比,夏妆、祁黄菊和雪映霞光的叶片SS含量在胁迫浓度为20 mmol/L时分别增加38.27%、40.27%和39.87%,在胁迫浓度为40 mmol/L时分别增加49.27%、52.82%和52.95%,在胁迫浓度为60 mmol/L时分别增加35.76%、45.06%和46.45%。总体上夏妆增幅最小,雪映霞光增幅最大。说明碱胁迫下,菊花进行自我保护使SS含量增加,让植株更易于维持正常的细胞膨压,以保证植物体内代谢活动的正常进行,但随着胁迫浓度增大及胁迫时间的延长,菊花的自我保护能力达到极限而使SS含量呈现降低趋势。
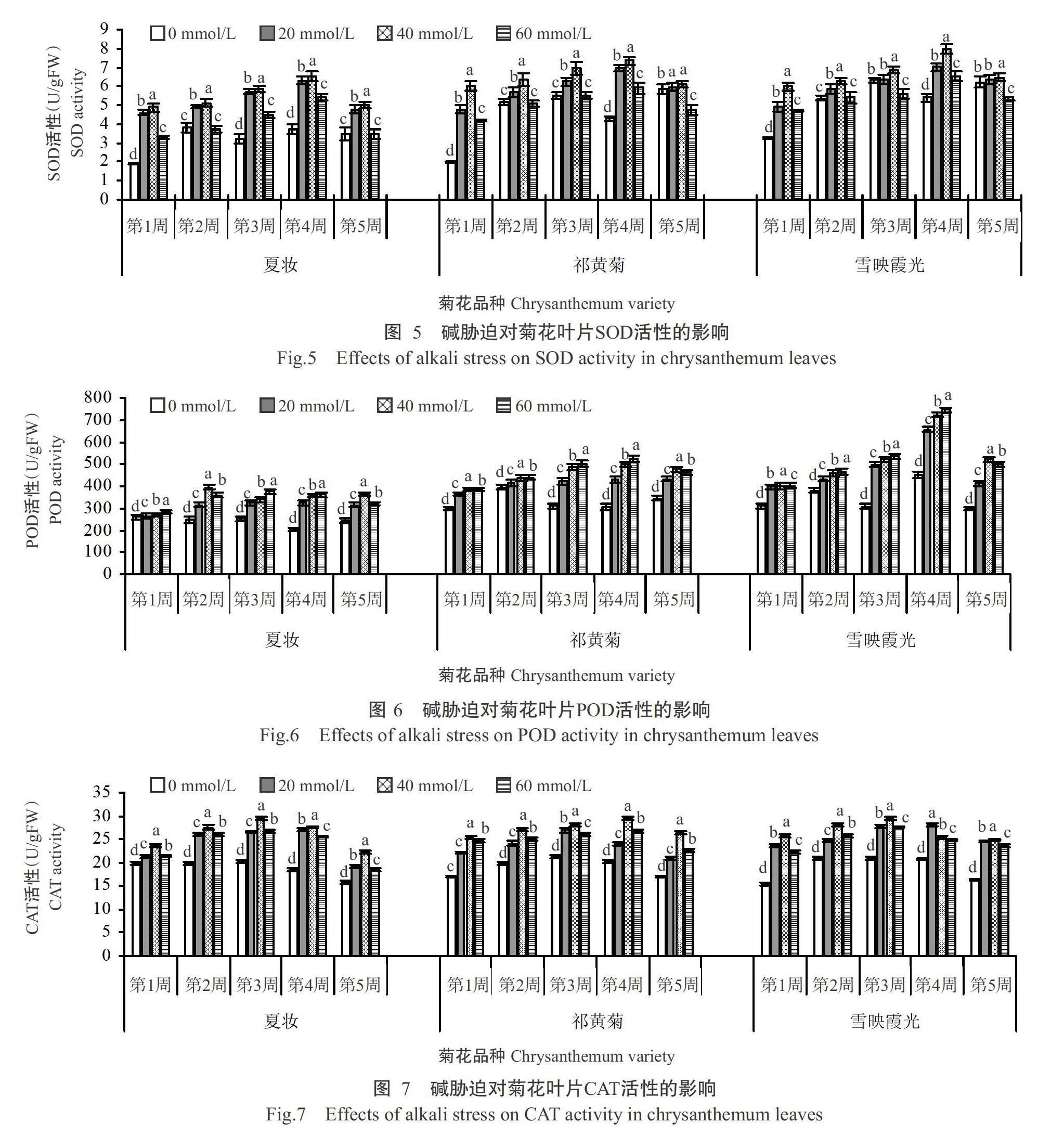
2. 3 碱胁迫对菊花叶片抗氧化酶活性的影响
2. 3. 1 SOD活性 由图5可知,菊花叶片SOD活性随碱胁迫浓度的升高及胁迫时间的延长呈先升后降的变化趋势,并于第4周时达最高值。其中,与CK相比,夏妆、祁黄菊和雪映霞光3个菊花品种叶片SOD活性的增幅在胁迫浓度为20 mmol/L时分别为69.34%、61.56%和30.21%,在脅迫浓度为40 mmol/L时分别为75.53%、71.72%和47.42%,在胁迫浓度为60 mmol/L时分别为46.03%、37.77%和21.68%。可见,不同浓度胁迫下3个菊花品种的叶片SOD活性变化差异性显著,夏妆增幅最大,雪映霞光增幅最小。
2. 3. 2 POD活性 由图6可知,3个菊花品种的叶片POD活性在胁迫前4周随着胁迫浓度的升高而持续升高。胁迫第5周时,与CK相比,胁迫浓度为20 mmol/L时,夏妆叶片POD活性的增幅最小(29.01%),雪映霞光增幅最大(38.23%);夏妆、祁黄菊和雪映霞光叶片POD活性的增幅在胁迫浓度为40 mmol/L时分别为48.96%、38.83%和74.73%,在胁迫浓度为60 mmol/L时分别为31.88%、34.24%和67.24%。不同浓度胁迫下,各品种的叶片POD活性变化差异性显著,夏妆增幅最小,雪映霞光增幅最大,说明夏妆耐碱性最弱,雪映霞光耐碱性最强。
2. 3. 3 CAT活性 由图7可知,3个菊花品种的叶片CAT活性随碱胁迫浓度的升高及胁迫时间的延长呈先升后降的变化趋势。夏妆和雪映霞光在第3周时达最高值,祁黄菊在第4周时达最高值。胁迫第5周时,与CK相比,夏妆、祁黄菊和雪映霞光叶片CAT活性的增幅在胁迫浓度为20 mmol/L时分别为21.68%、22.49%和50.10%,在胁迫浓度为40 mmol/L时分别为40.67%、54.94%和52.05%,在胁迫浓度为60 mmol/L时分别为15.74%、33.60%和44.09%。不同浓度胁迫下,各菊花品种的叶片CAT活性变化差异性显著,雪映霞光的叶片CAT活性增幅最大,夏妆增幅最小。
综上所述,不同品种菊花在碱胁迫下可诱导抗氧化酶的产生,使抗氧化酶活性增强的高峰时间不同,这也说明不同品种菊花随着胁迫时间的延长,其叶片SOD、POD和CAT活性最终整体上均降低,其中,在胁迫的第3周开始,叶片CAT活性呈下降趋势,在胁迫的第5周开始,叶片SOD和POD活性均呈下降趋势,说明菊花叶片抗氧化能力衰退,已不能有效地保护膜系统,抗氧化酶活性下降可能是菊花受碱害的主要原因。
2. 4 碱胁迫对膜脂过氧化指标MDA含量的影响
由图8可知,胁迫第5周时,与CK相比,夏妆、祁黄菊和雪映霞光叶片MDA含量的增幅在胁迫浓度为20 mmol/L时分别为40.88%、41.91%和22.83%,在胁迫浓度为40 mmol/L时分别为81.39%、65.68%和54.87%,在胁迫浓度为60 mmol/L时分别为104.39%、78.67%和65.27%;可见,夏妆增幅最大,雪映霞光增幅最小。由图8还可知,3个菊花品种的叶片MDA含量均在60 mmol/L胁迫下最大,在0 mmol/L时最小,不同浓度处理间差异显著。植株的MDA含量随着碱胁迫浓度的增加、时间的延长而增加,其中,雪映霞光的叶片MDA含量最小祁黄菊次之,夏妆最大,说明雪映霞光的耐碱性最强。
2. 5 碱胁迫对质膜透性的影响
在植物遭遇碱害后,质膜的渗透性增加;电解质外渗的速率是细胞膜稳定性的重要生理指标之一。如图9所示,菊花的叶片RE在胁迫浓度为0 mmol/L时最小、在60 mmol/L时最大,且均随着胁迫时间的延长及胁迫浓度的增加而增大。在胁迫第5周时,与CK相比,夏妆、祁黄菊和雪映霞光叶片RE的增幅在胁迫浓度为20 mmol/L时分别为6.54%、14.13%和3.33%,在胁迫浓度为40 mmol/L时分别为11.51%、22.19%和10.38%,在胁迫浓度为60 mmol/L时分别为22.39%、28.11%和15.49%。不同碱胁迫浓度下菊花的叶片RE存在着显著性差异,其中,雪映霞光和祁黄菊在受到碱胁迫時,植物细胞膜内电解质外渗较少,而夏妆植物细胞膜内电解质外渗较多,RE较高,碱害程度高,说明该品种自身耐碱性弱。
2. 6 碱胁迫对菊花SPAD值的影响
当植物受到碱胁迫时,SPAD也会受到影响,其减少的幅度可用以比较碱害的程度。由图10可看出,在碱浓度为60 mmol/L时均呈现3个菊花品种的叶片SPAD最小,0 mmol/L时SPAD最大,植株的SPAD随碱胁迫浓度的增加而减小,且差异显著。其中,在胁迫第5周时,夏妆的叶片SPAD下降幅度最大,最高浓度胁迫下仅为13.2,较CK下降60.28%,表明其叶绿体受到的损伤较大,耐碱能力较弱;而祁黄菊和雪映霞光的叶片SPAD下降幅度较缓,最高浓度胁迫下分别较CK下降54.01%和50.53%,说明其叶绿体受到伤害较小,耐碱能力较强。
2. 7 碱胁迫对菊花叶片WC的影响
植物叶片WC能很好地反映植物组织细胞的水分生理状态,可作为评定品种耐碱性的生理指标之一。由图11可知,在碱胁迫下,叶片WC随着胁迫时间的延长和胁迫浓度的增大均呈逐渐减少的趋势。与CK相比,各品种菊花在胁迫浓度为60 mmol/L时,叶片WC降幅最大,受碱害最严重。胁迫第5周时,与CK相比,夏妆、祁黄菊和雪映霞光叶片WC的降幅,在胁迫浓度为20 mmol/L时分别为13.69%、6.43%和1.26%,在胁迫浓度为40 mmol/L时分别为20.72%、13.36%、3.79%,在胁迫浓度为60 mmol/L时分别为23.85%、15.33%和8.59%。可见,祁黄菊和雪映霞光的叶片WC下降幅度较缓,说明其耐碱能力较强;3个菊花品种中雪映霞光耐碱性最强,夏妆耐碱性最弱。
2. 8 各指标的主成分分析
对不同品种菊花在不同浓度Na2CO3处理下9个生理指标的平均值进行主成分分析,结果如表1所示。第1和第2主成分的累计贡献率达92.853%,其概括了9个指标的大部分信息。
由表2可知,第1主成分中SS含量、SPAD、CAT活性和Pro含量有较大的得分系数,第2主成分中SOD活性、WC、POD活性和MDA含量有较大的得分系数。综合表1、表2携带的信息,可用SOD活性、POD活性、SS含量及Pro含量4个指标来概括9个指标中的大多数信息。
经过方差最大正交旋转得到旋转后因子的载荷矩阵(表3)。第1公因子主要由Pro含量、POD活性和SS含量3个因子决定,其因子载荷分别为0.961、0.959和0.940,可称其为Pro含量、POD活性和SS含量因子;第2公因子中叶片WC因子载荷最大,为0.929,可称之为叶片WC因子。
2. 9 碱胁迫下3个品种菊花公因子载荷矩阵耐碱性评价
根据表3所得旋转后的公因子载荷系数和表1的因子贡献率,得到2个公因子的表达式,如下所示:
?1=(0.961χ1+0.940χ2+0.690χ3+0.959χ4+0.928χ5+
0.237χ6+0.263χ7-0.408χ8+0.180χ9)×48.786%
?2=(-0.159χ1-0.231χ2+0.677χ3-0.008χ4-0.205χ5-
0.935χ6-0.922χ7+0.895χ8+0.929χ9)×44.066%
式中,F1和F2分别代表菊花样本公因子1和公因子2。χ1、χ2、χ3、χ4、χ5、χ6、χ7、χ8和χ9分别代表测定指标Pro含量、SS含量、SOD活性、POD活性、CAT活性、MDA含量、RE、SPAD和叶片WC。通过比较3个品种菊花的因子总值大小,最终确定其耐碱性排序,即雪映霞光>祁黄菊>夏妆(表4)。
2. 10 隶属函数分析
由表5和表6隶属函数值综合得分及排名可看出,3个菊花品种的耐碱性为雪映霞光最强,祁黄菊次之,夏妆最弱。
3 讨论
近年来,伴随着人类活动的影响土壤盐碱化程度加剧,部分地区土壤的pH达10以上,形成明显的碱斑,损害植物的生长和生产(Ma et al.,2018)。不同植物在碱胁迫下的形态结构变化表现不同,植物形态上的变化是植物对于胁迫条件适应性变化最直观的体现(Parida and Das,2005)。本研究中碱胁迫对3个品种菊花株高冠幅均有抑制作用,在较低浓度时植株会适当自我调节,随着胁迫浓度及胁迫周期的延长,植物的正常生长代谢被破坏,株高和冠幅生长减缓。
可溶性渗透物质Pro和SS在碱害胁迫条件下的积累中起到对细胞质渗透压调节剂的自我保护作用,同时也能保护体内氧化酶及细胞膜系统,从而使胁迫伤害得到缓解(雷蕾,2017)。在本研究中,碱胁迫下SS的积累反映了不同品种菊花在碱胁迫下的适应性反应,随着胁迫浓度增加及时间的延长,SS含量呈现的先升后降变化趋势与Na2CO3胁迫下甜菊叶片中可溶性渗透物的变化趋势相似(绳仁立等,2011),说明植物对于碱胁迫的自卫响应有一定的限度,当超过自卫能力范围后,SS积累会呈现降低现象(王旭明等,2018),这应该是植物受到碱害的原因之一。在碱胁迫下菊花由于蛋白质的合成受阻,因此通过分解蛋白质以抵抗毒害的能力增强,从而导致植物体内的氨基酸含量随之增加,其中Pro含量的增加尤为明显。Pro含量的提高是植物在逆境胁迫下的自我保护行为(赵海新,2020)。
碱胁迫通过增加细胞内产生的氧自由基离子对细胞进行迫害,从而阻碍植物正常生长发育(戴睿,2019)。因此植物对碱胁迫的适应过程依赖于抗氧化防御功能的增强(李璇等,2013)。植物在受到碱胁迫后,致使自由基积累,使得细胞膜中不饱和脂肪酸发生氧化作用,对膜脂系统产生影响,MDA即为这种膜脂过氧化反应的最终分解产物。MDA含量的高低可表示膜损伤程度的大小。因此,植物遭遇碱害后,细胞内O2-和H2O2等活性氧(ROS)(郭瑞等,2016;)及MDA含量增加,同時相对电解质泄漏量也增大(Ge et al.,2010)。本研究中,随着碱胁迫程度的加强,MDA含量及RE随之增大充分证实了这点。植物抗氧化酶系统主要由SOD、POD和CAT组成。本研究中这3种酶的活性均随碱胁迫程度的增强呈先升高后降低的变化趋势,表明菊花的抗氧化酶系统对碱胁迫有一定的响应,且不同品种对碱胁迫响应程度不同。当碱胁迫强度过大而使细胞无法很好的适应时,植物体内抗氧化酶活性会有所降低(郭楠楠等,2015)。
许多植物在碱性胁迫条件下光合作用降低(如蒸腾速率降低和气孔导度减小)(Wang et al.,2018)。本研究中随着碱胁迫时间的延长,各品种菊花SPAD值逐渐降低,说明叶片叶绿体光合膜系统被破坏,雪映霞光在胁迫第2周时出现比第1周降幅减小的情况,可能是因为在适应碱胁迫一段时间后叶片叶绿体光合膜系统得到一定程度的修复(刘佳,2017),叶绿素合成能力增强。
植物耐盐碱性是一个极其复杂的生理过程,任何单项机理的研究都不能全面而精准地评价某一植物的耐碱性(闫晶秋子等,2019),因此对植物的耐碱性评价应从研究其形态、生理及基因等综合指标的变化规律入手,从中筛选出主要的指标和成分来对植物进行耐碱性评价。本研究采用主成分降维分析法可将多个综合因子变量的减少转化为若干主要综合因子,起到采集浓缩数据的作用。综上所述,本研究通过这种耐碱性分析方法初步确定了菊花耐碱性指标筛选的有效方法和生理生化评价的指标,为菊花品种的耐碱性筛选奠定了基础。
4 结论
在碱胁迫下,不同品种菊花能在一定程度上维持细胞代谢平衡及体内化学物质的合成积累平衡,并进行活性氧的清除,以缓解并适应碱胁迫的伤害,但菊花的这种自卫能力有一定的局限性,当超过承受范围时,各品种菊花呈现不同程度碱害症状。3个菊花品种中,雪映霞光更能迅速响应胁迫,激活抗氧化系统,维持体内的氧化还原平衡,碱害症状明显小于祁黄菊和夏妆,对碱胁迫的耐受性最强,祁黄菊次之,夏妆耐碱能力最弱。
参考文献:
曹翠玲,麻鹏达. 2016. 植物生理学教学实验指导[M]. 杨凌:西北农林科技大学出版社. [Cao C L,Ma P D. 2016. Physiological teaching experiment guidance[M]. Yangling: Northwest A&F University Press.]
楚乐乐,罗成科,田蕾,张银霞,杨淑琴,李培富. 2019. 植物对碱胁迫适应机制的研究进展[J].植物遗传资源学报,20(4): 836-844. [Chu L L,Luo C K,Tian L,Zhang Y X,Yang S Q,Li P F. 2019. Research advance in Plants ada-ptation to alkali stress[J]. Journal of Plant Genetic Resources,20(4): 836-844.]
戴睿. 2019. 盐碱胁迫对植物生长的影响及应对措施[J]. 现代农业科技,(23): 58. [Dai R. 2019. Effects of saline-alkali stress on plant growth and countermeasures[J]. Mo-dern Agricultural Science and Technology,(23): 58.]
高俊凤. 2006. 植物生理学实验指导[M]. 北京: 高等教育出版社: 15-30.[Gao J F. 2006. Experimental guidance on plant physiology[M]. Beijing: Higher Education Press: 15-30.]
管志勇,陳发棣,滕年军,陈素梅,刘浦生. 2010. 5种菊花近缘种属植物的耐盐性比较[J]. 中国农业科学,43(4): 787-794. [Guan Z Y,Chen F D,Teng N J,Chen S M,Liu P S. 2010. Study on the NaCl tolerance in five plant species from Dendranthema and its relatives[J]. Scientia Agricultura Sinica,43(4): 787-794.]
郭楠楠,陈学林,张继,陈金元,朱愿军,丁映童. 2015. 柽柳组培苗抗氧化酶及渗透调节物质对NaCl胁迫的响应[J]. 西北植物学报,35(8):1620-1625. [Guo N N,Chen X L,Zhang J,Chen J Y,Zhu Y J,Ding Y T. 2015. Changes in antioxidase activity and osmotic adjusting substance of Tamarix chinensis seedlings under NaCl stress[J]. Acta Botanica Boreali-Occidentalia Sinica,35(8): 1620-1625.]
郭瑞,李峰,周际,李昊儒,夏旭,刘琪. 2016. 亚麻响应盐、碱胁迫的生理特征[J]. 植物生态学报,40(1): 69-79. [Guo R,Li F,Zhou J,Li H R,Xia X,Liu Q. 2016. Eco-physiological responses of linseed(Linum usitatissimum) to salt and alkali stresses[J]. Chinese Journal of Plant Eco-logy,40(1): 69-79.]
郝建军,康宗利,于洋. 2007. 植物生理学实验技术[M]. 北京: 化学工业出版社. [Hao J J,Kang Z L,Yu Y. 2007. Experimental techniques of plant physiology[M]. Beijing: Chemical Industry Press.]
雷蕾. 2017. 黄连花开花生物学和抗旱、抗盐碱研究[D]. 东北林业大学. [Lei L. 2017. The study on flowering biology
and drought and salt resistance of Lysimachica davurica
[D]. Harbin: Northeast Forestry University.]
李合生. 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社: 220-223. [Li H S. 2000. Principles and techniques of plant physiology and biochemistry experiment[M]. Beijing: Higher Education Press: 220-223.]
李荣华,阎旭东,赵松山. 2012. 名贵菊花品种耐盐性筛选浓度的确定[J]. 安徽农业科学,40(6): 3263-3264. [Li R H,Yan X D,Zhao S S. 2012. Confirmation of the concentration of salt-tolerance screening for famous and precious chrysanthemum[Dendronthema×Grandiflora(Ramat.)] varieties[J]. Journal of Anhui Agricultural Scien-ces,40(6): 3263-3264.]
李璇,岳红,王升,黄璐琦,马炯,郭兰萍. 2013. 影响植物抗氧化酶活性的因素及其研究热点和现状[J]. 中国中药杂志,38(7): 973-978. [Li X,Yue H,Wang S,Huang L Q,Ma J,Guo L P. 2013. Research of different effects on activity of plant antioxidant enzymes[J]. China Journal of Chinese Materia Medica,38(7): 973-978.]
刘杰. 2011. 向日葵对碱胁迫和盐胁迫适应机制比较[D]. 长春:东北师范大学. [Liu J. 2011. Comparison of adaptation mechanism to alkali stress and salt stress in sunflower(Helianthus annuus L.)[D]. Changchun: Northeast Normal University.]
刘佳. 2017. 李砧木榆叶梅对碱胁迫的响应机理研究[D]. 雅安:四川农业大学. [Liu J. 2017. Study on response mechanism of Prunus triloba Lindl. to alkali stress[D]. Yaan: Sichuan Agricultural University.]
绳仁立,原海燕,黄苏珍. 2011. Na2CO3胁迫对甜菊不同碱耐性品种生理代谢的影响[J]. 安徽农业科学,39(33): 20306-20309. [Sheng R L,Yuan H Y,Huang S Z. 2011. Influences of Na2CO3 stress on the physiological metabolisms of different Stevia rebaudiana alkali tolerance varie-ties[J]. Journal of Anhui Agricultural Sciences,39(33): 20306-20309.]
孫静. 2012. 切花菊抗旱性评价及抗旱机理研究[D]. 南京:南京农业大学. [Sun J. 2012. Drought tolerance evaluation and mechanisms involved in drought tolerance of cut chrysanthemum[D]. Nanjing: Nanjing Agricultural University.]
陶顺仙,张燕,李萍,燕佳琦,徐爱遐,黄镇. 2019. NaHCO3胁迫对甘蓝型油菜幼苗生长的影响及其耐盐候选基因预测[J]. 西北农业学报,28(12): 1978-1986. [Tao S X,Zhang Y,Li P,Yan J Q,Xu A X,Huang Z. 2019. Effects of NaHCO3 stress on growth of Brassica napus seedlings and prediction of candidate genes related to salt tolerance[J]. Acta Agriculturae Boreali-occidentalis Sinica,28(12): 1978-1986.]
王学奎. 2006. 植物生理生化实验原理和技术[M]. 第2版. 北京:高等教育出版社. [Wang X K. 2006. Principles and techniques of plant physiological and biochemical experiment[M]. The 2nd Edition. Beijing: Higher Education Press.]
王旭明,赵夏夏,陈景阳,龚茂健,杨善,谢平,莫俊杰,黄永相,叶昌辉,周鸿凯. 2018. 盐胁迫下水稻孕穗期SS和SPS活性与糖积累的响应及其相关性分析[J]. 江苏农业学报,34(3): 481-486. [Wang X M,Zhao X X,Chen J Y,Gong M J,Yang S,Xie P,Mo J J,Huang Y X,Ye C H,Zhou H K. 2018. The response and correlations between carbohydrate accumulation and activities of SPS,SS at booting stage of rice under salt stress[J]. Jiangsu Journal of Agricultural Sciences,34(3): 481-486.]
闫晶秋子,李钢铁,刘玉军,麻云霞,杨颖. 2019. 基于主成分分析及隶属函数法的巨菌草幼苗耐盐碱性评价[J]. 浙江农业学报,31(9): 1531-1540. [Yan J Q Z,Li G T,Liu Y J,Ma Y X,Yang Y. 2019. Evaluation of salt and alkali resistance of Pennisetum giganteum Z. X. Lin seedlings based on principal component analysis and membership function method[J]. Acta Agriculturae Zhejiangensis,31(9): 1531-1540.]
张海英. 2019. 盐胁迫和碱胁迫对制干辣椒生长发育及果实品质的影响[D]. 石河子:石河子大学. [Zhang H Y. 2019. Effects of salt stress and alkali salt stress on the growth and fruit quality of the industry pepper[D]. Shihezi:Shihezi University.]
张治安,张美善,蔚荣海. 2004. 植物生理学实验指导[M]. 北京: 中国农业科学技术出版社. [Zhang Z A,Zhang M S,Wei R H. 2004. Experimental guidance of plant physio-logy[M]. Beijing: China Agricultural Science and Technology Press.]
赵海新. 2012. 寒地水稻对碱胁迫的响应及鉴定指标评价[D]. 沈阳:沈阳农业大学. [Zhao H X. 2012. Identification indexes evaluation and response to alkali stress of early Japonica rice in cold region[D]. Shenyang:Shenyang Agricultural University.]
赵海新. 2020. 碱胁迫对水稻叶绿素及叶片脯氨酸和可溶性糖含量的影响[J]. 作物杂志,(1): 98-102. [Zhao H X. 2020. Effects of alkali stress on chlorophyll and the contents of proline and soluble sugar in rice[J]. Crops,(1): 98-102.]
Butt M,Ayyub C M,Amjad M,Ahmad R. 2016. Proline application enhances growth of chilli by improving physiological and biochemical attributes under salt stress[J]. Pakistan Journal of Agricultural Sciences,53(1): 43-49.
Ge Y,Li Y,Zhu Y M,Bai X,Lv D K,Guo D J,Ji W,Cai H. 2010. Global transcriptome profiling of wild soybean(Glycine soja) roots under NaHCO3,treatment[J]. BMC Plant Biology. doi: 10.1186/1471-2229-10-153.
Gong B,Wen D,Vandenlangenberg K,Wei M,Yang F J,Shi Q H,Wang X F. 2013. Comparative effects of NaCl and NaHCO3,stress on photosynthetic parameters,nutrient metabolism,and the antioxidant system in tomato leaves[J]. Scientia Horticulturae,157: 1-12.
Liu A L,Xiao Z X,Li M W,Wong F L,Yung W S,Ku Y S,Wang Q W,Wang X,Xie M,Yim A,Chan T F,Lam H M. 2018. Transcriptomic reprogramming in soybean seedlings under salt stress: Soybean transcriptome under salt stress[J]. Plant Cell & Environment. doi: 10.1111/pce. 13186.
Ma H Y,Li J P,Yang F,Lü X T,Pan Y P,Liang Z W. 2018. Regenerative role of soil seed banks of different successional stages in a saline-alkaline grassland in northeast China[J]. Chinese Geographical Science. doi: 10.1007/s11769-018-0966-2.
Parida A K,Das A B. 2005. Salt tolerance and salinity effects on plants: A review[J]. Ecotoxicology and Environmental Safety,60(3):324-349.
Wang L X,Fang C,Wang K. 2015. Physiological responses of Leymus Chinensis to long-term salt,alkali and mixed salt-alkali stresses[J]. Journal of Plant Nutrition,38(4): 526-540.
Wang Y X,Hu Y,Zhu Y F,Baloch A W,Jia X M,Guo A X. 2018. Transcriptional and physiological analyses of short-term Iron deficiency response in apple seedlings provide insight into the regulation involved in photosynthesis[J]. BMC Genomics. doi: 10.1186/s12864-018-4846-z.
Zhao Q,Suo J W,Chen S X,Jin Y D,Ma X L,Yin Z P,Zhang Y H,Wang T,Luo J,Jin W H,Zhang X,Zhou Z Q,Dai S J. 2016. Na2CO3-responsive mechanisms in halophyte Puccinellia tenuiflora roots revealed by physiological and proteomic analyses[J]. Scientific Reports. doi: 10.1038/srep32717.
(責任编辑 邓慧灵)
