红花檵木LcDRF1和LcDRF2基因克隆及亚细胞定位分析
2020-03-24张邦跃李彩虹刘旋廖晓珊荣朵艳
张邦跃 李彩虹 刘旋 廖晓珊 荣朵艳


張邦跃(1985-),博士,主要从事植物分子育种研究工作。先后主持或作为主要成员参与湖南省重点研发项目“高产类黄酮红花檵木种质资源的筛选研究”和“百合抗尖孢镰刀菌的分子机理研究及抗病品种的推广应用”、湖南省教育厅项目“红花檵木类黄酮生物合成途径调节因子MYB的分子克隆及功能鉴定”、株洲市科技计划项目“红花檵木叶色控制关键基因筛选及转基因研究”等科研项目8项。获海南省科学技术进步奖三等奖1项、湖南省科技厅成果登记证书1项、国家发明专利1项;参与培育百合新品种2个;在《Plant Physiology》《Molecules and Cells》《南方农业学报》《西北植物学报》《西北农业学报》等国内外期刊上发表学术论文10余篇,其中第一作者或通讯作者7篇(SCI收录期刊2篇)。
摘要:【目的】克隆红花檵木二氢黄酮醇-4-还原酶(DFR)基因(LcDFR1和LcDFR2),并对其进行亚细胞定位,为揭示红花檵木花青苷的分子合成机理提供理论依据。【方法】基于红花檵木转录组数据,以红花檵木叶片cDNA为模板,RT-PCR克隆LcDFR1和LcDFR2基因开放阅读框(ORF)序列,利用生物信息学软件对其进行分析,并通过烟草叶片瞬时表达方法观察蛋白的亚细胞定位情况。【结果】克隆获得的红花檵木LcDFR1和LcDFR2基因ORF序列分别为1014和993 bp,分别编码337和330个氨基酸残基。LcDFR1与LcDFR2的氨基酸序列相似性为77%,二者与其他物种DFRs氨基酸序列的相似性均较高,其中与葡萄、山核桃、拟南芥等双子叶植物的DFRs氨基酸序列相似性为64%~84%,而与单子叶植物玉米和水稻的DFRs氨基酸序列相似性为58%~61%,表明不同物种DFRs氨基酸序列具有较高的保守性。在基于DFRs氨基酸序列相似性构建的系统发育进化树上,LcDFR1与牡丹和芍药的DFRs聚为一类,而LcDFR2与烟草和番茄的DFRs聚为一类。结合前人研究结果推测LcDFR1和LcDFR2的氨基酸序列中均包含保守的NADP结合域和底物结合域。但三级结构预测结果均未预测到二者的底物结合位点,也未预测到LcDFR2的NADP结合位点。LcDFR1和LcDFR2亚细胞定位于细胞质,在细胞质中行使催化功能。【结论】LcDFR1和LcDFR2在红花檵木细胞质中催化花青苷物质的生物合成,但二者在进化过程中产生底物偏好性、催化能力等功能差异。
关键词: 红花檵木;二氢黄酮醇-4-还原酶(DFR);基因克隆;生物信息学分析;亚细胞定位
中图分类号: S687 文献标志码: A 文章编号:2095-1191(2020)12-2865-10
Abstract:【Objective】Cloning and subcellular localization of two dihydroflavonol-4-reductase(DFR) genes LcDFR1 and LcDFR2 from Loropetalum chinense var. rubrum were conducted to provide theoretical basis for further study of molecular mechanism of anthocyanin biosynthesis in L. chinense. 【Method】The open reading frame(ORF) sequences of two LcDFR1 and LcDFR2 were cloned with the templates of cDNA from the leaves of L. chinense using RT-PCR method based on the previous transcriptome data. Their amino acid sequences were analyzed by bioinformatics methods. The subcellular localization of LcDFRs proteins were detected by transient expression in tobacco leaves. 【Result】LcDFR1 and LcDFR2 ORF sequences cloned from L. chinense were 1014 and 993 bp, encoding 337 and 330 amino acids residues respectively. The amino acid sequences similarity between LcDFR1 and LcDFR2 was 77%. The identities of two LcDFRs amino acid sequences with DFRs amino acid sequence of other species were high. The similarity of two LcDFRs amino acid sequences with dicotyledon such as Vitis vinifera(NP_001268144.1), Paeonia lactiflora(QIC54081.1) and P. suffruticosa(AMW36065.1) was ranging from 64% to 84%, while their similarities with monocotyledon such as Zea mays (NP_001152467.2) and Oryza sativa (CAA69253.1) were from 58% to 61%, indicating that DFRs amino acid sequences from different species were highly conserved. Phylogenetic tree constructed based on DFRs amino acid sequence similarity showed that LcDFR1 was clustered into the same group with DFRs of P. lactiflora and P. suffruticosa, while LcDFR2 was clustered into the same group with DFRs of Nicotiana tabacum and Solanum lycopersicum. The amino acid sequence of both LcDFR1 and LcDFR2 contained conserved NADP binding motif and substrate binding domain, while their tertiary structure prediction results showed no available substrate binding site in both LcDFR and NADP binding site in LcDFR2. The subcellular localization studies have revealed that LcDFR1 and LcDFR2 were mainly cytoplasmic proteins, indicating that LcDFRs catalyzed substrates in cytoplasm. 【Conclusion】Both LcDFR1 and LcDFR2 may be involved in anthocyanin biosynthesis in cytoplasm of L. chinense. However, they may obtain different functions such as substrate prefe-rence and catalytic ability during evolution.
Key words:Loropetalum chinense var. rubrum; dihydroflavonol-4-reductase(DFR); gene cloning; bioinformatics analysis; subcellular localization
Foundation item: Project of Hunan Department of Science and Technology(2016NK2097); Hunan Natural Science Foundation(2019JJ50135); Project of Zhuzhou Science and Technology Bureau(Zhukefa〔2017〕68); Zhuzhou Science and Technology Talent Support Project(2019TJ-06)
0 引言
【研究意义】红花檵木(Loropetalum chinense var. rubrum)是金缕梅科(Hamamelidaceae)檵木属(Loropetalum)植物,为我国特有的观花观叶灌木,其叶子长时间保持红色,且植株易繁殖、耐修剪,是我国南方地区常用的园林观赏植物(彭闰珉和于晓英,2012)。叶片是否呈红色,主要取决于叶片中花青苷物质的含量,因此可将叶片中花青苷物质的含量及其稳定性作为新品种选育的核心指标(彭闰珉和于晓英,2012)。植物花青苷物质生物合成涉及到多种合成酶,其中,二氢黄酮醇-4-还原酶(DFR)是生物合成途径后期的第一个关键酶,可催化底物二氫黄酮醇流向花青苷代谢途径(Santos-Buelga et al.,2014;李亚丽等,2018)。因此,开展红花檵木DFRs基因克隆及功能研究,深入了解红花檵木该类同源基因在花青苷物质生物合成的作用,对促进红花檵木品种选育具有重要意义。【前人研究进展】DFR是烟酰胺腺嘌呤二核苷酸磷酸(NADP)依赖的还原酶家族成员,能利用NADP辅因子催化二氢黄酮醇还原形成无色、不稳定的无色花青素,以其为前体进一步合成有色花青苷物质(Petit et al.,2007)。大量研究证实,DFR在部分经济作物和观赏植物组织颜色形成过程中发挥着十分重要的作用。马铃薯(Solanum tuberosum L.)块茎中生成红色花青苷物质需要StDFR基因(又称R基因座)高效表达,如在白色块茎品种中过表达StDFR基因能完全恢复块茎花青苷物质的合成能力(De Jong et al.,2003;Zhang et al.,2009)。通过RNAi技术下调紫薯(Ipomoea batatas L. cv. Ayamurasaki)IbDFR基因的表达水平,结果发现其嫩叶、茎及贮藏根中的花青苷含量均显著减少,整株颜色变浅(Wang et al.,2013)。利用病毒诱导基因沉默(Virus-induced gene silencing,VIGS)技术瞬时沉默红叶海棠(Malus spp.)叶片中的McDFR基因,结果发现受到干扰的叶片由红色变绿色,花青苷物质含量显著降低,而在绿叶海棠中瞬时过表达McDFR基因会诱导叶片由绿色变红色,花青苷物质含量显著增加(Tian et al.,2015)。利用CRISPR/cas9技术对牵牛花(Ipomoea nil)的InDFR-B基因进行编辑,结果显示DFR-B双等位基因同时突变会导致花朵中花青苷物质消失,花由紫罗兰色变成白色(Watanabe et al.,2017)。通过在烟草中异源过表达其他植物的DFRs基因,包括苜蓿(Medicago trunca-tula)的MtDFR1(Xie et al.,2004)、玫瑰(Rosa rugosa)的RrDFR1(Luo et al.,2016)、矮牵牛(Petunia hybrida)的PhDFR(Luo et al.,2016)、美丽葡萄(Vitis bellula)的VbDFR(Zhu et al.,2018)和麝香兰(Muscari spp.)的MaDFR(Liu et al.,2019),结果发现这些植物的DFRs基因均参与花青苷物质的积累,也间接说明DFRs在进化过程中保持了生物学功能的保守性。此外,观赏植物DFRs基因克隆及功能研究也较多,如牡丹(Paeonia suffruticosa Andr.)(周琳等,2011)、百合(Lilium)(覃仁娟,2014)、香雪兰(Freesia hybrida)(Li et al.,2017)和一串红(Euphorbia pulcherrima willd. ex Klotzsch)(Gu et al.,2018)等,结果均显示这些观赏植物DFRs基因与花青苷物质的积累紧密相关。【本研究切入点】目前,关于红花檵木DFRs基因的相关研究鲜见报道。【拟解决的关键问题】基于前期红花檵木转录组数据,通过RT-PCR克隆出LcDFR1和LcDFR2基因cDNA序列,利用生物信息学软件对其编码的氨基酸序列进行功能分析,构建35S::LcDFR1-GFP和35S::LcDFR2-GFP表达载体并检测编码蛋白的亚细胞定位,为深入了解LcDFR1和LcDFR2基因在红花檵木花青苷合成过程中的调控作用提供参考依据。
1 材料与方法
1. 1 试验材料
供试红花檵木为4年生扦插苗,盆栽于温室内;用于瞬时表达的本氏烟草种植于植物光照培养箱中。植物多糖多酚总RNA提取试剂盒购自北京艾德莱生物科技有限公司,PrimeSTAR GXL DNA聚合酶购自宝日医生物技术(北京)有限公司;大肠杆菌DH5α感受态细胞、PCR产物磁珠回收试剂盒、无缝连接试剂盒和M-MuLV逆转录酶购自生工生物工程(上海)股份有限公司;AGL-0根癌农杆菌感受态细胞为自制保存;pCAMBIA1305-GFP为湖南工业大学生命科学与化学学院实验室改造保存。主要仪器设备:Fusion FX5-XT凝胶成像系统(Vilber,法国)和SP8激光共聚焦显微镜(Leica,德国)。
1. 2 RNA提取和第一链cDNA合成
利用植物RNA提取试剂盒提取红花檵木叶片总RNA,并用1%琼脂糖凝胶电泳进行检测。以总RNA为模板,利用Oligo dT18引物和M-MuLV逆转录酶合成第一链cDNA,于-20 ℃保存备用。
1. 3 LcDFR1和LcDFR2基因克隆
基于红花檵木转录组测序结果,从中筛选到3个DFRs同源cDNA序列,经序列分析发现仅有2个基因包含有完整的开放阅读框(ORF),长度分别为1014和993 bp,分别命名为LcDFR1和LcDFR2。利用Primer Premier 5.0设计这2个基因5'-非编码区(5'-UTR)和3'-UTR区域的特异引物DFR1-F/DFR1-R和DFR2-F/DFR2-R(表1),用于扩增LcDFR1和LcDFR2基因cDNA序列(包含部分5'-UTR、3'-UTR及完整ORF)。以cDNA第一链为模板,利用PrimeSTAR GXL DNA聚合酶扩增LcDFR1和LcDFR2基因cDNA序列。反应体系25.0 μL:5×PrimeSTAR GXL Bu-ffer 5.0 μL,2.5 mmol/L dNTP Mixture 2.0 μL,10 mmol/L上、下游引物各0.5 μL,cDNA模板0.3 μL,PrimeSTAR GXL DNA聚合酶0.5 μL,灭菌水补足至25.0 μL。扩增程序:95 ℃预变性2 min;98 ℃ 10 s,55 ℃ 15 s,68 ℃ 80 s,进行35个循环;72 ℃延伸5 min,10 ℃保存。PCR产物经琼脂糖凝胶电泳检测后,挑取条带大小正确的目的片段送至生工生物工程(上海)股份有限公司进行双向测通,以確定转录组数据中LcDFR1和LcDFR2基因cDNA序列的正确性。
1. 4 生物信息学分析
利用DNAMAN 9.0对LcDFR1和LcDFR2基因的ORF及其编码氨基酸序列进行分析。利用NCBI数据库的BLASTp(https://blast.ncbi.nlm.nih.gov/Blast.cgi)对LcDFR1和LcDFR2的同源氨基酸序列进行搜索。利用DNAMAN 9.0对不同物种DFRs氨基酸序列进行多重比对,并利用MEGA 6.0的邻接法(Neighbor-joining)构建系统发育进化树。最后,使用SWISS-MODEL(https://swissmodel.expasy.org/)进行三级结构预测。
1. 5 植物表达载体构建及亚细胞定位
根据红花檵木LcDFR1和LcDFR2基因的ORF序列,利用Primer Premier 5.0设计扩增ORF片段(不包含终止密码子)的特异引物DFR1-GFP-F/DFR1-GFP-R和DFR2-GFP-F/DFR2-GFP-R(表1)。利用PrimeSTAR GXL DNA聚合酶对LcDFR1和LcDFR2基因ORF片段进行扩增,PCR产物经磁珠法回收后利用无缝连接试剂盒连接到Xba I酶切线性化的pCAMBIA1305-GFP载体上,获得重组植物表达载体35S::LcDFR1-GFP和35S::LcDFR2-GFP,转化大肠杆菌DH5α感受态细胞,经菌液PCR鉴定后挑取阳性菌送至生工生物工程(上海)股份有限公司测序。将测序正确的35S::LcDFR1-GFP和35S::LcDFR2-GFP重组植物表达载体,利用热激法分别转化AGL-0根癌农杆菌感受态细胞,经PCR鉴定获得阳性单克隆菌,将其接种至LB液体培养基中,经振荡培养后离心积菌用渗透液重悬,用注射器吸取5 mL菌液侵染3周龄本氏烟草叶片进行瞬时表达,培养3 d后用SP8激光共聚焦显微镜观察荧光信号(王晰等,2018)。
2 结果与分析
2. 1 LcDFR1和LcDFR2基因克隆及序列分析结果
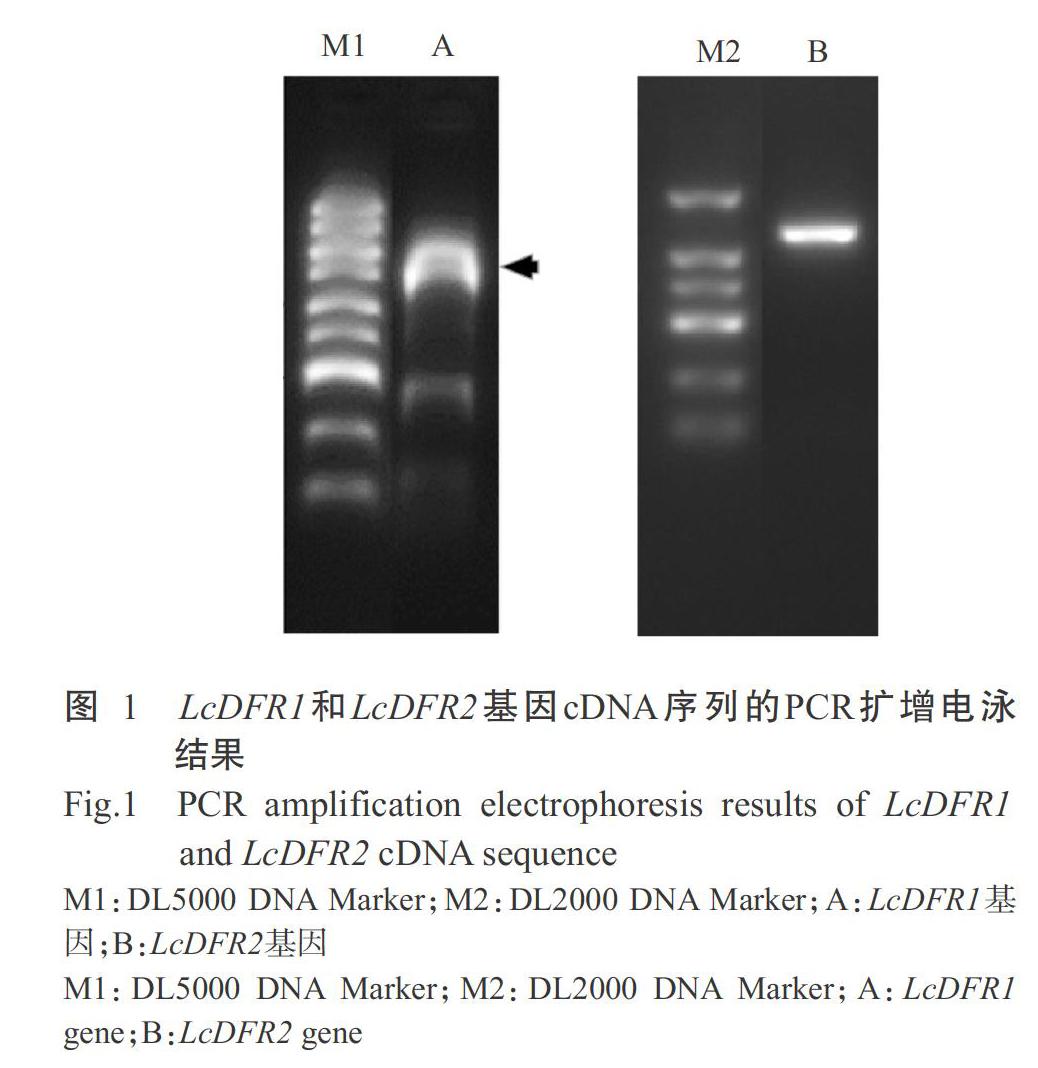
以红花檵木叶片的cDNA第一链为模板,分别用特异引物DFR1-F/DFR1-R和DFR2-F/DFR2-R进行PCR扩增,获得LcDFR1和LcDFR2基因cDNA序列,PCR扩增产物用1%琼脂糖凝胶电泳检测,结果显示目标扩增条带均约1100 bp,与预期结果相符,其中LcDFR1基因的扩增产物还存在非特异性扩增(图1)。测序结果显示,LcDFR1和LcDFR2基因cDNA序列与转录组测序结果一致,包含完整ORF序列,其中LcDFR1基因ORF全长为1014 bp,编码337个氨基酸残基(图2),LcDFR2基因ORF全长为993 bp,编码330个氨基酸残基(图3)。
2. 2 同源性比对分析及系统进化树构建结果
将LcDFR1和LcDFR2氨基酸序列与其他植物的DFRs氨基酸序列进行比对,包括葡萄(Vitis vini-fera)VvDFR(NP_001268144.1)、山核桃(Carya illinoinensis)CiDFR(QCT85679.1)、芍药(Paeonia lactiflora)PlDFR(QIC54081.1)、牡丹(P. suffruticosa)PsDFR(AMW36065.1)、拟南芥(Arabidopsis thalia-na)AtDFR(NP_199094)、烟草(Nicotiana tabacum)NtDFR(NP_001312661)、玉米(Zea mays)ZmDFR(NP_001152467.2)和水稻(Oryza sativa)OsDFR(CAA 69253.1),结果如图4所示。LcDFR1与LcDFR2的氨基酸序列相似性为77%,二者与其他物种DFRs氨基酸序列的相似性均较高,其中与葡萄、山核桃、拟南芥等双子叶植物的DFRs氨基酸序列相似性为64%~84%,而与单子叶植物玉米和水稻的DFRs氨基酸序列相似性为58%~61%。通过与其他相关文献(Lacombe et al.,1997;Li et al.,2017;Liu et al.,2019)中注释的DFRs特征结合域氨基酸序列进行比对分析,结果发现LcDFR1和LcDFR2的氨基酸序列均包含有NADP结合域和底物结合域,且与底物特异性相关的2个氨基酸残基高度保守(图4)。由图5可知,LcDFR1与牡丹和芍药的DFRs聚为一类,而LcDFR2与烟草和番茄(Solanum lycopersicum)的DFRs聚为一类。
2. 3 LcDFR1和LcDFR2蛋白三级结构预测结果
利用SWISS-MODEL对LcDFR1和LcDFR2蛋白的三级结构进行预测,结果(图6)发现二者均与葡萄的VvDFR(2c29.1.A)为同源模板,其中,LcDFR1蛋白与VvDFR(2c29.1.A)的氨基酸序列相似性达84.48%,GMQE达0.96,QMEAN为0.66,说明其与同源模板的匹配度较高,且三级结构预测结果显示,其含有1个保守的NADP结合位点,但未预测到底物结合位点(图6-A);LcDFR2蛋白与VvDFR(2c29.1.A)的氨基酸序列相似性为79.27%,GMQE达0.95,QMEAN为0.52,说明匹配度也较高。此外,LcDFR2与LcDFR1蛋白的三级结构重叠率高,空间结构相似(图6-B),但其三级结构未预测到保守的NADP结合位点和底物结合位点。
2. 4 植物表达载体构建及亚细胞定位结果
分别以DFR1-GFP-F/DFR1-GFP-R和DFR2-GFP-F/DFR2-GFP-R为引物,PCR扩增出LcDFR1和LcDFR2基因的ORF序列,长度在1000 bp左右(图7-A和图7-B),并将其与线性化的pCMBIA1305-GFP进行连接,获得重组植物表达载体35S::LcDFR1-GFP和35S::LcDFR2-GFP(图8),转化大肠杆菌DH5α感受态细胞,PCR鉴定及测序验证后,提取阳性质粒并转化到AGL-0根癌农杆菌,PCR鉴定出阳性单克隆菌(图7-C和图7-D)。采用叶片注射法用将阳性单克隆菌液侵染本氏烟草叶片下表皮,培养3 d培养后,利用Leica SP8激光共聚焦显微镜观察烟草表皮细胞中的荧光信号,结果如图9所示,LcDFR1和LcDFR2蛋白的亚细胞定位结果比较相似,主要定位于烟草表皮细胞的细胞质中。
3 讨论
红花檵木是我国特有的观花观叶植物,而花青苷物质形成分子机制研究是红花檵木分子育种的核心内容。DFR是植物花青苷物质生物合成途径后期的第一个关键酶,能引导底物二氢黄酮醇流向花青苷代谢途径,对植物组织的颜色形成起关键作用。已有研究表明,马铃薯(De Jong et al.,2003;Zhang et al.,2009)、紫薯(Wang et al.,2013)、海棠(Tian et al.,2015)、牵牛花(Watanabe et al.,2017)等植物的DFRs在花青苷物质生物合成途径中是必需催化酶,若DFRs基因缺失或被干扰则会导致花青苷物质合成障碍。在烟草中异源表达苜蓿(Xie et al.,2004)、玫瑰(Luo et al.,2016)、矮牵牛(Luo et al.,2016)、美丽葡萄(Zhu et al.,2018)、麝香兰(Liu et al.,2019)等植物DFRs基因能显著提升花青苷物质的合成。
本研究克隆获得LcDFR1和LcDFR2基因cDNA序列,分别编码337和330个氨基酸残基,与多种木本植物DFRs氨基酸序列长度接近,包括葡萄VvDFR(337个氨基酸残基)、枫香树LfDFR(337个氨基酸残基)和山核桃CiDFR(339个氨基酸殘基)。前人研究表明,DFRs作为NADP依赖的家族成员,其氨基酸序列N端有一段约含21个氨基酸残基的NADP结合域特征序列(Lacombe et al.,1997;Li et al.,2017;Liu et al.,2019)。葡萄VvDFR的NADP结合域序列为VTGASGFIGSWLVMRLLERGY,LcDFR1和LcDFR2的NADP结合域序列与其相比分别存在1和3个氨基酸残基的差异,说明不同物种DFRs的NADP结合域序列较保守。DFR的底物是二氢黄酮醇类,包括二氢槲皮素(DHQ)、二氢山奈酚(DHK)和二氢杨梅素(DHM)。DFR对底物的特异识别主要依赖于一段约28个氨基酸残基的底物结合域序列,某个特定氨基酸残基的变化均会导致该酶对底物识别的改变(Johnson et al.,2001;Chen et al.,2020)。如非洲菊(Gerbera jamesonii)GjDFR的N134突变为L134,导致该酶只催化DHK、不催化DHQ和DHM;E145突变成L145则导致该酶彻底失去催化活性。红盖鳞毛蕨(Dryopteris erythrosora)DeDFR1和DeDFR2的底物特异识别氨基酸残基之一为R(对应于GjDFR的N134),决定了这2个酶只能催化DHK和DHQ,不能催化DHM(Chen et al.,2020)。虽然不同植物DFRs底物识别序列存在一定差异,但决定底物识别能力的2个氨基酸残基保守性非常高,如VvDFR的N133和E144,这种情况也出现在LcDFR1和LcDFR2中。由于不同DFRs在催化底物时可能表现出不同的底物偏好性,如香雪兰的3个FhDFRs和麝香兰的MaDFR均偏好催化DHM,导致这2种植物花朵主要累积飞燕草色素而显蓝色(Li et al.,2017;Liu et al.,2019)。LcDFR1和LcDFR2对底物的偏好性还需进一步通过体内和体外试验进行探究。
本研究的LcDFR1和LcDFR2蛋白三级结构预测结果显示,二者与葡萄的VvDFR(2c29.1.A)(Petit et al.,2007)晶体结构相似性最高。蛋白三级结构预测是基于已有的同源蛋白晶体三维结构数据进行分析,从而获得目标蛋白的三维模型,比基于氨基酸序列进行一级结构分析结果更复杂和多样化。在VvDFR的晶体结构中,NADP和DHQ位于三级空间结构的催化腔中,且NADP配体与VvDFR蛋白上19个非连续氨基酸残基相互作用(包括疏水作用、氢键等),而底物DHQ与VvDFR蛋白上8个非连续氨基酸残基相互作用(Petit et al.,2007),完全不同于通过比对氨基酸序列一级结构中的NADP特征结合域(约21个连续氨基酸序列)和底物特征结合域(约27个连续氨基酸序列)(Li et al.,2017;Liu et al.,2019)。LcDFR1与VvDFR的氨基酸序列相似性为84%,高于LcDFR2与VvDFR的氨基酸序列相似性(79%),由于氨基酸序列相似性越高,蛋白三级结构预测质量越高,故LcDFR1的QMEAN为0.66,高于LcDFR2的QMEAN(0.52)。相比LcDFR2的预测结果,LcDFR1所形成的NADP结合空间结构可能更接近于模板蛋白,因此仅LcDFR1预测到NADP结合位点;二者底物结合位点空间结构可能与模板蛋白间均存在一定差异,造成底物结合位点的预测缺失。但蛋白预测结果并不完全反映真实的蛋白结构,蛋白配体/底物结合位点的预测缺失可能是由于数据库中可用的DFR蛋白—配体结合晶体模型太少(仅有葡萄VvDFR),不能反映出特征结合位点空间结构及不同配体结合的多样性,从而不利于LcDFR1和LcDFR2三级结构中结合位点的预测。今后应通过构建更多DFR蛋白—配体的晶体结构数据,以利于提高其他未知DFRs的三级结构预测的准确性。此外,本研究发现LcDFR1和LcDFR2蛋白三级结构预测结果重叠率很高,但系统发育进化树上二者被分到不同的分支中,其中LcDFR1与牡丹和芍药聚为一类,而LcDFR2与烟草和番茄聚为一类,可能是由于进化过程中产生了底物偏好性、催化能力等功能差异。
经前人研究证实,花青苷物质首先在细胞质中合成,随后通过转运蛋白或微自噬介导运输至液泡中(Gomez et al.,2011;Chanoca et al.,2015),推测DFR蛋白作为花青苷物质生物合成的关键酶,定位于细胞质中。为进一步验证DFRs亚细胞定位情况,本研究构建植物表达载体35S::LcDFR1-GFP和35S::LcDFR2-GFP(GFP位于LcDFRs的C端,以避免其对N端位置NADP结合域的功能影响),通过根癌农杆菌介导转入本氏烟草叶片进行瞬时表达,在SP8激光共聚焦显微镜观察荧光信号,结果发现,LcDFR1和LcDFR2蛋白主要定位于细胞质中,与美丽葡萄VbDFR-GFP的亚细胞定位相同(Zhu et al.,2018),表明LcDFR1和LcDFR2是在细胞质中催化底物,符合花青苷物质合成的特点。今后可通过体内、体外试验验证LcDFR1和LcDFR2的催化活性和底物特异性,从而进一步分析其功能,为红花檵木或其他观赏植物的彩色育种提供参考。
4 结论
LcDFR1和LcDFR2在红花檵木细胞质中催化花青苷物质的生物合成,但二者在进化过程中产生底物偏好性、催化能力等功能差异
参考文献:
李亚丽,李欣,肖婕,李瑞玲,杨华丽,孙勃,汤浩茹. 2018. 二氢黄酮醇-4-还原酶在花青素合成中的功能及调控研究进展[J]. 西北植物学报,38(1):187-196. [Li Y L,Li X,Xiao J,Li R L,Yang H L,Sun B,Tang H R. 2018. Function and regulation characterization of dihydroflavonol 4-reductase in anthocyanin biosynthesis[J]. Acta Botanica Boreali-Occidentalia Sinica,38(1):187-196.]
彭闰珉,于晓英. 2012. 湖南檵木属植物的园林应用现状调查与分析[J]. 天津农业科学,18(6): 152-155. [Peng R M,Yu X Y. 2012. Investigation and analysis of the landscape application present situation of Loropetalum in Hunan Province[J]. Tianjin Agricultural Sciences,18(6):152-155.]
覃仁娟. 2014. 百合DFR基因克隆及功能初步鉴定[D]. 雅安:四川农业大学. [Qin R J. 2014. Cloning and functional initial charaterization of the gene encoding dihydroflavonol 4-reductase(DFR) from Lily[D]. Yaan:Sichuan Agricultural University.]
王晰,丁文杰,李婭,刘家伟,王良桂,岳远征. 2018. 长筒石蒜花青素合成酶基因LlANS的克隆与表达分析[J]. 河南农业大学学报,52(4):611-617. [Wang X,Ding W J,Li Y,Liu J W,Wang L G,Yue Y Z. 2018. Cloning and expre-ssion analysis of anthocyanidin synthase(LlANS) in Lycoris longituba[J]. Journal of Henan Agricultural University,52(4):611-617.]
周琳,王雁,任磊,彭镇华. 2011. 牡丹二氢黄酮醇4-还原酶基因PsDFR1的克隆及表达分析[J]. 植物生理学报,47(9): 885-892. [Zhou L,Wang Y,Ren L,Peng Z H. 2011. Cloning and expression analysis of dihydroflavonol 4-reductase gene PsDFR1 from tree peony(Paeonia su-ffruticosa Andr.)[J]. Plant Physiology Journal,47(9):885-892.]
Chanoca A,Kovinich N,Burkel B,Stecha S,Bohorquez-Restrepo A,Ueda T,Eliceiri K W,Grotewold E,Otegui M S. 2015. Anthocyanin vacuolar inclusions form by a microautophagy mechanism[J]. The Plant Cell,27(9):2545-2559.
Chen X F,Liu W L,Huang X Y,Fu H H,Wang Q X,Wang Y F,Cao J G. 2020. Arg-type dihydroflavonol 4-reductase genes from the fern Dryopteris erythrosora play important roles in the biosynthesis of anthocyanins[J]. PLoS One,15(5):e0232090.
De Jong W S,De Jong D M,De Jong H,Kalazich J,Bodis M. 2003. An allele of dihydroflavonol 4-reductase associa-ted with the ability to produce red anthocyanin pigments in potato(Solanum tuberosum L.)[J]. Theoretical and App-lied Genetics,107(8):1375-1383.
Gomez C,Conejero G,Torregrosa L,Cheynier V,Terrier N,Ageorges A. 2011. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATE transporters and GST[J]. The Plant Journal,67(7):960-970.
Gu Z Y,Chen H,Yang R N,Ran M H. 2018. Identification of DFR as a promoter of anthocyanin accumulation in poinsettia(Euphorbia pulcherrima,willd. ex Klotzsch) bracts under short-day conditions[J]. Scientia Horticulturae,236:158-165.
Johnson E T,Ryu S,Yi H,Shin B,Cheong H,Choi G. 2001. Alteration of a single amino acid changes the substrate specificity of dihydroflavonol 4-reductase[J]. The Plant Journal,25(3):325-333.
Lacombe E,Hawkins S,van Doorsselaere J,Piquemal J,Goffner D,Poeydomenge O,Boudet A M,Grima-Pettenati J. 1997. Cinnamoyl CoA reductase,the first committed enzyme of the lignin branch biosynthetic pathway:Cloning,expression and phylogenetic relationships[J]. The Plant Journal,11(3):429-441.
Li Y Q,Liu X X,Cai X Q,Shan X T,Gao R F,Yang S,Han T T,Wang S C,Wang L,Gao X. 2017. Dihydroflavonol 4-reductase genes from Freesia hybrida play important and partially overlapping roles in the biosynthesis of flavonoids[J]. Frontiers in Plant Science,8:428.
Liu H L,Lou Q,Ma J R,Su B B,Gao Z Z,Liu Y L. 2019. Cloning and functional characterization of dihydroflavonol 4-reductase gene involved in anthocyanidin biosynthesis of grape hyacinth[J]. International Journal of Molecular Sciences,20(19):4743.
Luo P,Ning G H,Wang Z,Shen Y X,Jin H N,Li P H,Huang S S,Zhao J,Bao M Z. 2016. Disequilibrium of flavonol synthase and dihydroflavonol-4-reductase expre-ssion associated tightly to white vs. red color flower formation in plants[J]. Frontiers in Plant Science,6:1257.
Petit P,Granier T,dEstaintot B L,Manigand C,Bathany K,Schmitter J M,Lauvergeat V,Hamdi S,Gallois B. 2007. Crystal structure of grape dihydroflavonol 4-reductase,a key enzyme in flavonoid biosynthesis[J]. Journal of Molecular Biology,368(5):1345-1357.
Santos-Buelga C,Mateus N,De Freitas V. 2014. Anthocyanins. Plant pigments and beyond[J]. Journal of Agricultural and Food Chemistry,62(29):6879-6884.
Tian J,Han Z Y,Zhang J,Hu Y J,Song T T,Yao Y C. 2015. The balance of expression of dihydroflavonol 4-reductase and flavonol synthase regulates flavonoid biosynthesis and red foliage coloration in crabapples[J]. Scientific Reports,5:12228.
Wang H X,Fan W J,Li H,Yang J,Huang J R,Zhang P. 2013. Functional characterization of dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses[J]. PLoS One,8(11):e78484.
Watanabe K,Kobayashi A,Endo M,Sage-Ono K,Toki S,Ono M. 2017. CRISPR/Cas9-mediated mutagenesis of the dihydroflavonol-4-reductase-B(DFR-B) locus in the Japanese morning glory Ipomoea(Pharbitis) nil[J]. Scientific Reports,7:10028.
Xie D Y,Jackson L A,Cooper J D,Ferreira D,Paiva N L. 2004. Molecular and biochemical analysis of two cDNA clones encoding dihydroflavonol-4-reductase from Medicago truncatula[J]. Plant Physiology,134(3):979-994.
Zhang Y F,Cheng S P,De Jong D,Griffths H,Halitschke R,De Jong W. 2009. The potato R locus codes for dihydroflavonol 4-reductase[J]. Theoretical and Applied Gene-tics,119(5):931-937.
Zhu Y,Peng Q Z,Li K G,Xie D Y. 2018. Molecular cloning and functional characterization of a dihydroflavonol 4-reductase from Vitis bellula[J]. Molecules,23(4):861.
(責任编辑 陈 燕)
