常绿阔叶次生林不同经营方式对土壤微生物呼吸及其熵值的影响
2020-03-08张丽纪娇娇杨智杰
张丽 纪娇娇 杨智杰

摘 要 :次生林在森林碳匯中发挥着重要作用,其合理经营对于提高森林生产力和土壤碳汇功能具有重要的意义。以福建三明地区两种经营方式的常绿阔叶次生林(天然更新、人工促进天然更新)为对象,研究次生林不同经营方式对土壤微生物呼吸及其熵值的影响。结果表明:(1)次生林不同经营方式下的土壤微生物呼吸速率均随土层深度的增加而下降,人工促进天然更新(简称人促更新)林土壤微生物呼吸速率比天然更新林约高15%;(2)不同经营方式次生林表层土层土壤微生物生物量(MBC)含量显著高于深层,天然更新林除40~60 cm土层MBC含量低于人促更新林外,其余各土层MBC含量均高于人促更新林;(3)天然更新林深层土壤微生物熵(qMB)显著高于人促更新林,但表层低于人促更新林;(4)人促更新林各土层微生物代谢熵(qCO2)均高于天然更新林。研究结果表明,常绿阔叶林自然更新与人促更新的表层土壤有机碳活性与利用效率没有显著差异,但是自然更新模式下深层土壤微生物的活性程度高于人促更新方式,且微生物对碳源的利用效率也较高,有利于土壤有机碳固持,提升土壤质量。因此,深层土壤有机碳对不同经营方式的响应研究对于提升土壤有机碳吸存具有重要意义。
关键词:常绿阔叶次生林;微生物呼吸;微生物熵;微生物代谢熵
中图分类号:S718.4 文献标志码:A 文章编号:0253-2301(2020)11-0041-08
DOI: 10.13651/j.cnki.fjnykj.2020.11.007
Abstract: The secondary forest plays an important role in the forest carbon sequestration, and its rational management is of great significance to improve the forest productivity and soil carbon sequestration capacity. In this study, the effects of different management methods of the secondary forest on the soil microbial respiration and entropy were studied, by taking the evergreen broad-leaved secondary forest with two management modes (natural regeneration, and natural regeneration with artificial aid) in Sanming of Fujian as the object. The results showed that: (1) The soil microbial respiration rate under different management modes of secondary forest decreased with the increase of soil depth, and the soil microbial respiration rate of the human-assisted naturally regenerated secondary forest was about 15% higher than that of the natural regenerated secondary forest. (2) The content of soil microbial biomass carbon (MBC) in the surface layer of the secondary forest with different management modes was significantly higher than that in the deep layer. Except that the MBC content in 40-60 cm soil layer of natural regenerated secondary forest was lower than that of human-assisted naturally regenerated secondary forest, the MBC content of other soil layers were higher than that of human-assisted naturally regenerated secondary forest. (3)The soil microbial entropy (qMB) in the deep layer of natural regenerated secondary forest was significantly higher than that of human-assisted naturally regenerated secondary forest, while the soil microbial entropy (qMB) in the surface layer of natural regenerated secondary forest was lower than that of human-assisted naturally regenerated secondary forest. (4) The microbial metabolic quotient (qCO2) in each soil layer of human-assisted naturally regenerated secondary forest was higher than that of natural regenerated secondary forest. This study showed that there was no significant difference in the soil organic carbon activity and utilization efficiency in the surface layer between the natural regeneration and human-assisted naturally regeneration of evergreen broad-leaved forest, but the activity of deep soil microorganisms under the mode of natural regeneration was higher than that of human-assisted naturally regeneration mode. And the microbial utilization efficiency of carbon source was also higher, which was beneficial to the immobilization of soil organic carbon and the promotion of soil quality. Therefore, it would be of great significance to study the response of deep soil organic carbon to different management modes for improving the absorption and storage of soil organic carbon.
Key words: Evergreen broad-leaved secondary forest; Microbial respiration; Microbial entropy; Microbial metabolic quotient
常绿阔叶次生林是我国最重要的速生林类型之一[1],发挥着比人工林更强的固碳能力[2],其不同更新方式将影响地上植被覆盖度和林窗大小,導致林分结构和土壤性质产生差异,林下微气候发生变化,土壤中的微生物对碳源的利用能力也各不相同[3],从而对土壤碳吸收、分配、固存和排放产生重要影响。因此,了解常绿阔叶林的次生更新方式对土壤微生物利用效率的影响具有重要意义。
土壤微生物是植物生长养分的来源之一和土壤养分的储存库之一[4],同时是土壤有机质和养分转化与循环的动力。微生物生物量碳和基础呼吸是土壤质量、地下过程和对气候变化响应的重要指标[5]。土壤微生物熵和代谢熵是土壤有机质质量变化的敏感指标[6],与传统土壤微生物评价指标相比,可以更敏感地反映土壤微生物对环境变化的响应[7],其变化影响土壤碳吸存效率,对模拟土壤有机碳储量有潜在重要作用[8]。与传统的土壤养分相比,这些变化相对较快的土壤微生物学性质可以有效地评价森林经营措施及土地利用变化的短期效应[9]。目前国内外研究主要关注于次生林对地上和地下碳库的影响,如南美热带森林转换、温带红松林等[10-11],但有关次生林的经营探讨还缺乏研究。因此如何优化次生林经营模式以及提高森林碳储量已成为研究者越来越关注的问题。
米槠Castanopsis carlesii是我国东部湿润亚热带山地海拔<500 m的顶极群落建群种,其天然林采伐迹地经自然更新后将形成以米槠为主的次生林[12]。项目组前期已对亚热带常绿阔叶次生林的土壤养分、可溶性有机质数量和光谱特征、酶化学计量等方面进行了研究[13-14]。本研究选取中亚热带山区(福建三明)不同经营方式常绿阔叶次生林为研究对象,对不同经营方式常绿阔叶林土壤微生物呼吸及其熵值的变化进行研究,为进一步探讨次生林经营对土壤碳吸存的影响机制提供基础数据,也为我国亚热带地区常绿阔叶次生林合理经营、发挥最大碳汇效益提供科学依据。
1 材料与方法
1.1 研究区域与样地概况
研究区位于三明森林生态系统与全球变化福建省野外科学研究站(117°36′E,26°19′N),该区属中亚热带海洋性季风气候,年均温20.1℃,降水多集中在3月至8月,多年平均降水量1656 mm,相对湿度81%。样地平均海拔高度330 m,坡度35°~45°。土壤由花岗岩发育的红壤组成,土层厚度超过1 m。
本研究中亚热带常绿阔叶次生林系以米槠为主的常绿阔叶林采伐迹地,形成了两种主要的更新模式:米槠天然林经强度择伐并历时40多年的封山育林后,天然更新形成的米槠天然更新林;以及经过强度择伐后天然更新,并在更新过程中人为去除非目的树种形成的米储人工促进天然更新(简称人促更新)林。乔木层主要树种为米槠Castanopsis carlesii、闽粤栲Castanopsis fissa、黄丹木姜子Litsea elongata、新木姜子Neolitsea aurata等,以米槠为优势树种,林下灌木主要有鼠刺Itea chinensis、黄瑞木Adinandra millettii、山矾Symplocos caudata、大叶紫珠Callicarpa macrophylla和山血丹Ardisia lindleyana等构成。草本有黑莎草Gahnia tristis、剑叶耳草Hedyotis caudatifolia Merr: et Metcalf、芒萁Dicramopteris dichotama和毛冬青Ilex pubescens等。不同更新方式试验地基本特征见表1。
1.2 样品采集与处理
2017年4月,分别在米槠天然林、米槠天然更新林和人工促进天然更新林随机布设3个20 m×20 m样方,在每块标准样地内采用S型选取5个取样点,利用土钻分别采集0~10、>10~20、>20~40、>40~60、>60~80 cm土壤,所有土样均立即放入4℃冰冷藏箱并立即带回实验室处理。土样去除杂质、根系和凋落物等,用四分法分成两份,一份过2 mm土壤筛网后于4℃冰箱冷藏保存,用于土壤微生物生物量碳、可溶性有机碳、可溶性有机氮、微生物呼吸速率的测定[15]。另一份在1周内自然风干并过0.45 μm筛网后进行土壤总有机碳、总氮等理化性质的测定。试验开始前土壤基本理化性质如表2。
1.3 测定方法
土壤总有机碳、总氮含量使用碳氮元素分析仪(Elemental EL MAX CNS analyzer,德国)进行测定。
微生物呼吸速率采用室内培养法测定。称取相当于10 g干土质量的鲜土,平铺于500 mL培养瓶中,并用保鲜膜封口扎若干小孔,以便通风并防止土壤水分损失过快。在22℃的无照明培养瓶预培养10 h,使其适应培养环境后,将培养瓶取出进行洗气。采用注射器来回抽送数次使瓶内气体充分混匀后,抽气50 mL,使用Li840 CO2/H2O分析仪测定CO2浓度作为初始值,然后将培养瓶放回培养箱8 h后,使用Li840 CO2/H2O分析仪再次测定CO2浓度。
土壤微生物生物量碳采用底物诱导呼吸法(SIR)测定。底物诱导呼吸法的基本原理是添加葡萄糖形成系列葡萄糖浓度梯度,直到达到最高呼吸速率(一般不超过1 h),以此作为最大初始呼吸速率,用以估算土壤微生物生物量[16]。与微生物呼吸速率培养试验相似,本试验在22℃的无照明培养箱中培养1周,培养至第4 d时,对培养瓶进行洗气。培养期结束后,再次对培养瓶进行洗气,确保培养土壤已达到平衡稳定状态。随后,用注射器将800 μg·g-1的葡萄糖溶液快速均匀添加到培养瓶中的土壤表面,并保证培养瓶内土壤水量不超过田间持水量的60%,之后再将培养瓶放置于培养箱1 h,土壤呼吸稳定后用Li840 CO2/H2O分析仪测定CO2浓度作为初始值,随即将培养瓶放回培养箱2 h后用Li840 CO2/H2O分析仪再次测定CO2浓度。
1.4 计算和统计方法
土壤微生物呼吸速率[17]:
上述公式中,R表示土壤微生物CO2呼吸速率(mg·kg-1·h-1);K表示气体转化为标准系数的参数(g·m-3),V表示培养瓶内总气体体积(m3);m表示干土重(g);△c/△t为单位时间内CO2浓度的变化量(μmol·mol-1·h-1);T表示培养温度(℃);12/44为CO2中C的占比。
土壤微生物生物量碳(MBC)含量[18]:
上述公式中, 表示土壤培养前后CO2浓度变化量(μmol·mol-1·h-1);V表示培养瓶内总气体体积(m3);t表示培养时间(h);m表示干土重(g)。
土壤微生物熵(%)[19]:
土壤代谢熵(h-1)[19]:
式中CO2-C为培养8 h内微生物呼吸排放出的碳(μmol·mol-1·h-1)。
采用单因素方差分析和独立样本t检验两种分析方法分析各林分指标的差异(α=0.05)。所有统计分析均采用Excel和SPSS 20.0统计软件,相关图表在 Origin 9.0软件下完成,图表中数据为平均值±标准差。
2 结果与分析
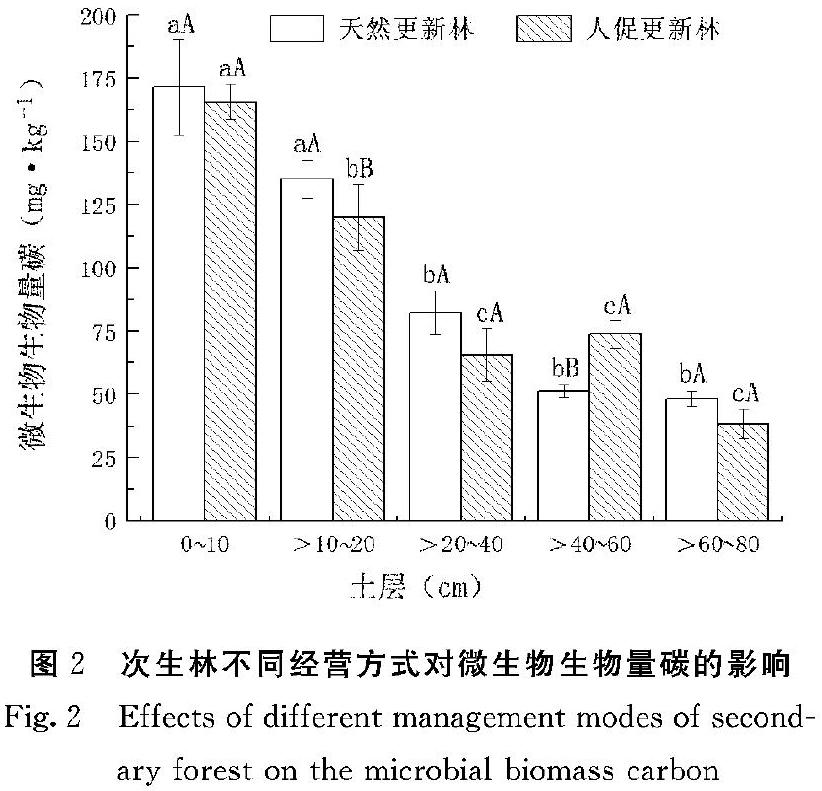
2.1 土壤微生物呼吸速率和微生物生物量碳的变化
次生林两种经营模式下,土壤微生物呼吸速率的变化趋势相近(图1),随土层深度的增加而下降,0~10 cm土壤微生物呼吸速率(0.44 mg·kg-1·h-1)均为最高。天然更新和人促更新林的微生物呼吸在0~10 cm 和>20~40 cm 土层无显著差异,但米槠人促更新林>10~20 cm和>40~80 cm土层的微生物呼吸速率显著高于天然更新(P<0.05),>10~20 cm差异最大,人促更新林是天然更新林的1.58倍。
土层深度对两种不同经营方式次生林MBC有显著影响(图2),表层0~10和>10~20 cm土层MBC含量显著高于深层土壤(>20~80 cm)(P<0.05)。除>40~60 cm天然更新林MBC含量高于人促更新林外,其余各土层MBC含量均高于人促更新林;但仅在>40~60 cm土层上差异显著(P<0.05),人促更新林是天然更新林的1.43倍。
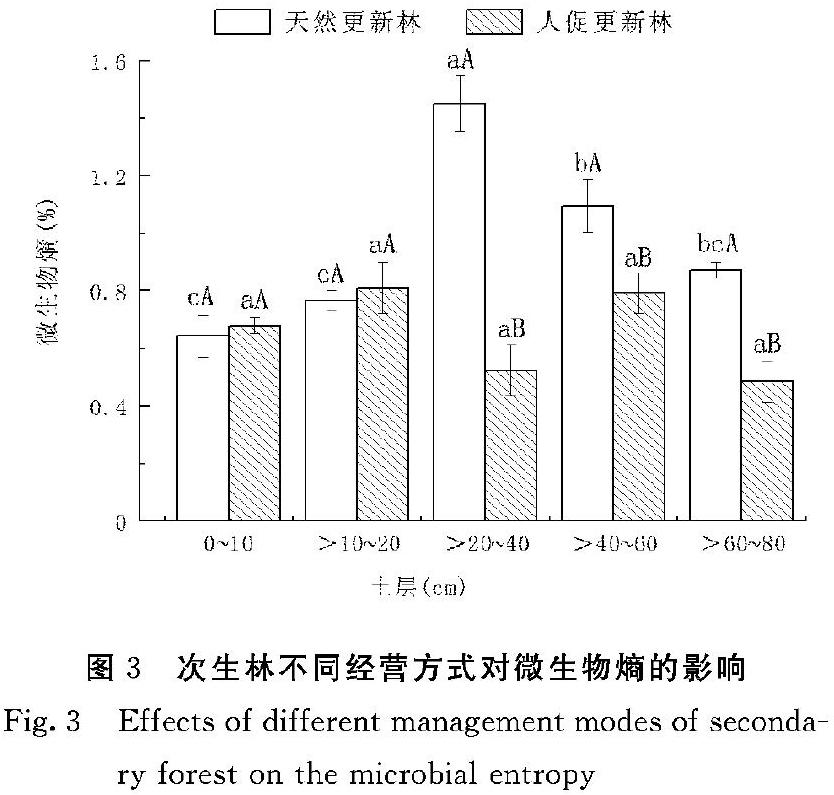
2.2 土壤微生物熵(qMB)和代谢熵(qCO2)的变化
次生林两种经营方式下表层0~10 cm和>10~20 cm土壤qMB无显著差异,但是>20~80 cm土壤中,天然更新林显著高于人促更新林;其中,天然更新林>20~40、>40~60和>60~80 cm土壤qMB分别是人促更新林的2.77倍、1.38倍和1.8倍(P<0.05)。天然更新林>20~80 cm土壤qMB显著高于0~20 cm,但人促更新林各土层间差异不显著(图3)。
从图4可知,人促更新林各土层微生物代谢熵均高于天然更新林。天然更新林0~10 cm土层qCO2显著高于其余土層,但人促更新林不同土壤层无明显变化规律。除>10~20 cm和>60~80 cm外,米槠人促更新林各土层qCO2与天然更新林无显著差异,其中人促更新林>10~20 cm和>60~80 cm分别是天然更新林的1.82倍(P<0.01)和1.71倍(P<0.05)。
3 讨论与结论
3.1 不同经营方式对常绿阔叶次生林土壤微生物呼吸和微生物生物量碳的影响
常绿阔叶次生林两种不同经营方式下,土壤微生物呼吸速率随土层深度的增加而减小,这与大多数的研究结果相似[18-19]。土壤微生物呼吸是微生物作用下的土壤有机质分解过程,受温度和降水等气候因子、土壤的养分供给、数量及活性等调控[20]。Holden等[3]研究表明,森林干扰改变土壤和植被特征,进而影响微生物的生化过程。本研究中,人促更新林除>10~20 cm和>40~80 cm两个土层的土壤微生物呼吸速率显著高于天然更新林外(P<0.05),其余土层两者并无显著差异,这可能是人促更新林林分密度小于天然更新林,林冠截留小,穿透雨量较大[21],地表凋落物在雨水淋溶和分解作用下分解腐烂速度较快,表层土壤养分易随雨水下渗至>10~20 cm土层,大量的易变化且有效性高的可溶性有机质输入到土壤[22],提高微生物呼吸速率。此外,人促更新林深层>40~80 cm土层微生物呼吸速率显著高于天然更新林,也可能是由于天然更新林>40~80 cm土层土壤含水率较低,土壤溶液中可溶性有机质的扩散受到阻碍,细菌等微生物可能处于饥饿状态,间接导致土壤微生物呼吸速率较低[23]。
有研究认为,MBC和土壤碳氮[24]、土壤有机碳矿化特征[25]等呈显著正相关关系。本研究中,天然更新林除>40~60 cm土层外的各土层MBC含量均高于人促更新林,原因可能是天然更新林有机碳、全氮含量和碳氮比均高于人促更新林,在碳氮比较高的土壤中,微生物有更多的碳素可转化为生物量[26-27],土壤养分含量是影响不同经营模式常绿阔叶次生林微生物生物量碳的主要因素。虽然本研究中天然更新林与人促更新林林龄一致,但2012-2016年5年的监测结果显示天然更新林年平均凋落物量(6.44±0.55)t·hm-2高于人促更新林(5.99±0.22)t·hm-2,同时人促更新林皆伐后不定期去除非目的树种等经营方式导致凋落物回归量降低,微生物的碳源和养分来源减少[28],不利于土壤微生物的生存。
3.2 不同经营方式对常绿阔叶次生林土壤微生物熵(qMB)和代谢熵(qCO2)的影响
土壤微生物熵是土壤有机碳向微生物生物量碳转化速度快慢的敏感指标[29],其比值大小是衡量有机碳积累或损失的一个重要指标。该比值高表示土壤碳呈累积趋势,土壤有机碳的活性程度越高[30]。有研究表明,土壤微生物熵主要由微生物同化和呼吸决定,受土壤资源和环境条件的影响[31]。Spohn等[32]在德国下萨克森州Solling山脉的研究指出,土壤碳氮是微生物代谢的基本能量和养分来源。Zhou等[33]对中国森林生态系统土壤微生物碳氮及相关系数的207项研究成果整合发现微生物熵随土壤C∶N的增加而显著减小。但Xu等[34]模型研究结果表明,基质质量(例如C∶N,木质素含量等)对土壤有机碳的微生物同化有积极影响,表现为基质质量越高,微生物熵越高。本研究中,天然更新林>20~80 cm土层qMB显著高于0~20 cm土层,这与Wen等[35]对不同林龄马尾松纯林和混交林土壤微生物熵的研究结果相似。人促更新林各土层的有机碳、微生物生物量碳含量和根系生物量均低于天然更新林,这可能是由于营林干扰中,根系残留死亡加速土壤内部微生物活动,加速了深层有机碳分解,土壤有机碳氮比较低,单位土壤有机碳的微生物生物量升高。但周义贵等[36]对川西亚高山不同土地利用类型的微生物碳动态特征的研究发现,植物根系通过向土壤中分泌易分解有机物质,增大根系周围微生物的活性从而提高微生物对碳源的利用效率。
微生物代谢熵是微生物可矿化碳与微生物生物量碳的比率[37],即单位生物量的微生物呼吸,是控制土壤异养呼吸的基本因素[38]。qCO2越低,表明土壤微生物群落对基质碳源的利用效率较高[39],单位微生物固定的碳越多[40],有利于提升土壤质量。本研究中,人促更新林各土层qCO2均高于天然更新林,原因可能是人促更新林土壤微生物生物量碳含量普遍低于天然更新林,同时在经营措施的影响下林地凋落物养分归还量及碳源减少,微生物生存条件较差,对碳源的利用效率低下[41],同时也表明人促更新林受环境胁迫或干扰的程度较高[42]。此外,天然更新林0~10 cm土层qCO2显著高于其余土层,这可能是由于米槠天然林进行强度择伐后经过天然更新等经营措施导致表层土壤细菌含量升高,与真菌相比,细菌产生较少的生物量,碳利用效率较低,呼吸损耗也较多[43],表现出更高的呼吸速率[44]。而天然更新林>10~80 cm土层qCO2普遍较低则可能是因为较深层土壤有机质来源较少,微生物需保持较高的碳利用效率维持自身生存[45]。
中亚热带常绿阔叶次生林人促更新模式对表层土壤微生物碳库积累起到积极作用。自然更新模式下土壤微生物对碳源的利用效率高于人工促进更新模式,更有利于土壤有机质固存,但在森林受到干扰的情况下,可能加大深层土壤碳排放。
参考文献:
[1]陈刚,涂利华,彭勇,等.瓦屋山扁刺栲-中华木荷常绿阔叶次生林土壤有机碳组分特征[J].生态学报,2015,35(18):6100-6109.
[2]王芸,欧阳志云,郑华,等.中国亚热带典型天然次生林土壤微生物碳源代谢功能影响因素[J].生态学报,2012,32(6):1839-1845.
[3]HOLDEN S R,TRESEDER K K.A meta-analysis of soil microbial biomass responses to forest disturbances[J].Frontiers in Microbiology,2013,4(163):1-17.
[4]SPEDDING T A,HAMEL C,MEHUYS GR,et al.Soil microbial dynamics in maize-growing soil under different tillage and residue management systems[J].Soil Biology and Biochemistry,2004,36(3):499-512.
[5]LIU Y,WEI X,GUO X,et al.The long-term effects of reforestation on soil microbial biomass carbon in subtropic severe red soil degradation areas[J].Forest Ecology and Management,2012,285:77-84.
[6]YAN T,YANG L,CAMPBELL CD.Microbial biomass and metabolic quotient of soils under different land use in the Three Gorges Reservoir area[J].Geoderma,2003,115(1-2):129-138.
[7]JIA GM,CAO J,WANG C,et al.Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin,northwest China [J].Forest Ecology and Management,2005,217(1):117-125.
[8]FANG C,MONCRIEFF J B.The variation of soil microbial respiration with depth in relation to soil carbon composition [J].Plant and Soil,2005,268:243-253.
[9]張伟东,汪思龙,颜绍馗,等.杉木根系和凋落物对土壤微生物学性质的影响[J].应用生态学报,2009,20(10):2345-2350.
[10]CONTI G,PREZ-HARGUINDEGUY N,QUTIER F,et al.Large changes in carbon storage under different land-use regimes in subtropical seasonally dry forests of southern South America[J].Agriculture Ecosystems and Environment,2014,197:68-76.
[11]赵华晨,高菲,李斯雯,等.长白山阔叶红松林和杨桦次生林土壤有机碳氮的协同积累特征[J].应用生态学报,2019,30(5):196-205.
[12]YANG Y S,WANG L X,YANG Z J,et al.Large ecosystem service benefits of assisted natural regeneration[J].Journal of Geophysical Research:Biogeosciences,2018,123:676-687.
[13]元晓春,林伟盛,蒲晓婷,等.更新方式对亚热带森林土壤溶液可溶性有机质数量及化学结构的影响[J].应用生态学报,2016,27(6):1845-1852.
[14]袁萍,周嘉聪,张秋芳,等.中亚热带不同森林更新方式生态酶化学计量特征[J].生态学报,2018,38(18):6741-6748.
[15]林杉,陈涛,赵劲松,等.不同培养温度下长期施肥水稻土的有机碳矿化特征[J].应用生态学报,2014,25(5):1340-1348.
[16]CREPIN J,JOHNSON L R.Soil sampling and methods of analysis[J].Journal of Environmental Quality,1993,38(1):15-24.
[17]吴东梅,郭剑芬,张政,等.DOM对米槠次生林不同土层土壤微生物呼吸及其熵值的影响[J].生态学报,2018,38(11):3806-3815.
[18]周焱,徐宪根,王丰,等.武夷山不同海拔梯度土壤微生物生物量微生物呼吸及其商值(qMB,qCO2)[J].生态学杂志,2009,28(2):265-269.
[19]纪娇娇,郑蔚,杨智杰,等.亚热带森林转换对土壤微生物呼吸及其熵值的影响[J].生态学报,2020,40(3):800-807.
[20]贾丙瑞,周广胜,王风玉,等.土壤微生物与根系呼吸作用影响因子分析[J].应用生态学报,2005,16(8):1547-1552.
[21]江淼华,吕茂奎,胥超,等.亚热带米槠次生林和杉木人工林林冠截留特征比较[J].水土保持学报,2017,31(1):116-121.
[22]NILSEN P,STRAND L T.Thinning intensity effects on carbon and nitrogen stores and fluxes in a Norway spruce(Picea abies(L.)Karst.)stand after 33 years[J].Forest Ecology and Management,2008,256(3):201-208.
[23]HICKS L C,MASUDUR R M,MONIQUE C,et al.The legacy of mixed planting and precipitation reduction treatments on soil microbial activity,biomass and community composition in a young tree plantation[J].Soil Biology and Biochemistry,2018,124:227-235.
[24]周正虎,王傳宽.帽儿山地区不同土地利用方式下土壤-微生物-矿化碳氮化学计量特征[J].生态学报,2017,37(7):2428-2436.
[25]何相宜,刘肖肖,戴伟.天山云杉林土壤有机碳矿化特征[J].西北林学院学报,2019,34(2):1-7.
[26]SHOEMAKER W R,LOCEY K J,LENNON J T.A macroecological theory of microbial biodiversity[J].Nature Ecology Evolution,2017,1(5):107.
[27]ZHOU Z,WANG C,JIANG L,et al.Trends in soil microbial communities during secondary succession[J].Soil Biology and Biochemistry,2017,115:92-99.
[28]XIONG Y,ZENG H,XIA H,et al.Interactions between leaf litter and soil organic matter on carbon and nitrogen mineralization in six forest litter-soil systems[J].Plant and Soil,2014,379(1-2):217-229.
[29]SPARLING G P,PANKHURST C,DOUBE B M,et al.Soil microbial biomass,activity and nutrient cycling as indicators of soil health[M].Biological indicators of soil health,1997.
[30]SINGH J S,RAGHUBANSHI AS,SINGH R S,et al.Microbial biomass acts as a source of plant nutrients in dry tropical forest and savanna[J].Nature,1989,338(6215):499-500.
[31]MANZONI S,TAYLOR P,RICHTER A,et al.Environmental and stoichiometric controls on microbial carbon-use efficiency in soils[J].New Phytologist,2012,196(1):79-91.
[32]SPOHN M,CHODAK M.Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils[J].Soil Biology and Biochemistry,2015,81:128-133.
[33]ZHOU Z H,WANG C K.Reviews and syntheses:Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China′s forest ecosystems[J].Biogeosciences,2015,12(22):6751-6760.
[34]XU X,SCHIMEL J P,THOMTON P E,et al.Substrate and environmental controls on microbial assimilation of soil organic carbon:a framework for Earth system models[J].Ecology Letters,2014,17(5):547-555.
[35]WEN L,LEI P,XIANG W,et al.Soil microbial biomass carbon and nitrogen in pure and mixed stands of Pinus massoniana and Cinnamomum camphora differing in stand age[J].Forest Ecology and Management,2014,328:150-158.
[36]周义贵,郝凯婕,李贤伟,等.川西亚高山不同土地利用类型对土壤微生物量碳动态特征的影响[J].自然资源学报,2014,29(11):1944-1956.
[37]HARTMAN W H,RICHARDSON C J.Differential nutrient limitation of soil microbial biomass and metabolic quotients(qCO2):is there a biological stoichiometry of soil microbes?[J].Plos One,2013,8(3):1-14.
[38]XU X,SCHIMEL J P,JANSSENS I A,et al.Global pattern and controls of soil microbial metabolic quotient [J].Ecological Monographs,2017,87(3):429-441.
[39]ZHAO F Z,REN CJ,HAN X H,et al.Changes of soil microbial and enzyme activities are linked to soil C,N and P stoichiometry in afforested ecosystems [J].Forest Ecology and Management,2018,427:289-295.
[40]邱曦,吕茂奎,黄锦学,等.不同培养温度下严重侵蚀红壤的有机碳矿化特征[J].植物生态学报,2016,40(3):236-245.
[41]ANDERSON TH,DOMSCH KH.Soil microbial biomass:The eco-physiological approach[J].Soil Biology Biochemistry,2010,42(12):2039-2043.
[42]ODUM E P.Trends Expected in Stressed Ecosystems[J].American Institute of Biological Sciences,1985,35(7):419-422.
[43]NIE M,PENDALL E,Bell C,et al.Positive climate feedbacks of soil microbial communities in a semi-arid grassland[J].Ecology Letters,2013,16(2):234-241.
[44]LAUGHLIN R J,RTTING T, MLLER C,et al.Effect of acetate on soil respiration,N2O emissions and gross N transformations related to fungi and bacteria in a grassland soil[J].Applied Soil Ecology,2009,42(1):25-30.
[45]SHIHAN A,HTTENSCHWILER S,MILCU A,et al.Changes in soil microbial substrate utilization in response to altered itter diversity and precipitation in a Mediterranean shrubland[J].Biology Fertility of Soils,2017,53(2):171-185.
(責任编辑:柯文辉)
