不同基因型烟草抗逆生理指标对低氮胁迫的响应
2020-03-08王俊梁喜欢王子璇尼金玉杨帅强郑云杨祥飞郑九洲朱肖文刘齐元
王俊 梁喜欢 王子璇 尼金玉 杨帅强 郑云 杨祥飞 郑九洲 朱肖文 刘齐元
摘要:抗逆生理指標可以衡量植物对逆境响应的程度,为了探究不同基因型烟草品种抗逆生理指标对低氮胁迫的响应,以耐低氮、氮高效型14P9,低氮敏感、氮低效型中烟100、K394和氮高效型G80为试验材料进行水培试验,测定烟草苗期低氮胁迫15、30 d的叶、茎、根中过氧化物歧化酶(SOD)活性、丙二醛含量和脯氨酸含量。结果表明,低氮胁迫 15 d 时,14P9叶、茎、根中具有更高的SOD活性,根中丙二醛含量低于低氮敏感型烟草品种;低氮胁迫30 d时,14P9叶、茎、根中丙二醛含量显著小于低氮敏感型品种,叶中脯氨酸含量显著大于低氮敏感型品种,茎中显著小于低氮敏感型品种。因此,低氮胁迫下耐低氮型烟草品种具有更高的SOD活性、更多的脯氨酸积累量,能减少低氮胁迫对植物的伤害。
关键词:烟草;低氮胁迫;超氧化物歧化酶;丙二醛;脯氨酸;抗逆生理
中图分类号:S572.01 文献标志码:A 文章编号:1002-1302(2020)21-0097-07
氮素的缺乏是一种胁迫作用,会引起植物体内抗氧化系统酶活性的变化,但变化趋势与植物的基因型和所处的环境有关[1]。过氧化物歧化酶(SOD)是植物抵抗低氮胁迫的重要保护酶,在响应逆境胁迫时,植物产生的自由基会诱发膜脂过氧反应,生成丙二醛等有害物质,植物可通过抗氧化酶系统来清除自由基,维持稳定[2]。脯氨酸含量的积累是许多植物对生物和非生物胁迫的一个应激反应,其积累发生在各种胁迫中[3-6]。脯氨酸是植物渗透调节的主要物质,植物在逆境中通过其含量的积累来减少逆境伤害[7]。大量研究表明,胁迫会提高SOD活性和增加丙二醛含量,也会增加脯氨酸的含量,以此来抵御植物受到的逆境伤害[8-11]。李春燕等研究表明,提高植物体内的抗氧化酶活性对减轻胁迫危害有一定的积极作用[12]。植物抗氧化酶活性和渗透调节物质含量的差异是植物品种在忍受胁迫时存在差异的生理基础[13]。低氮胁迫下,耐低氮品种和不耐低氮品种会显著增加丙二醛含量,不耐低氮品种增加更显著,同样,SOD活性也是相同的趋势,且不耐低氮品种的活性比耐低氮品种的活性低[14]。肖玉洁等研究表明,烟草低温耐受性品种和低温敏感型品种在胁迫下SOD 活性会明显降低,丙二醛含量则上升[15]。有研究表明,提高SOD活性会减少植物体内的丙二醛含量,脯氨酸含量会随着胁迫作用的进行而增加,并认为可将其含量高低作为衡量植物抗性的指标[6,16]。
1 材料与方法
1.1 试验材料
试验于2019年4月在江西农业大学农学院多功能实验室进行。材料为笔者所在课题组前期筛选出的耐低氮、氮高效烟草品种14P9,氮高效型品种G80,低氮敏感、氮低效品种中烟100(简称ZY100)和K394[17]。
1.2 试验方法
试验在室内进行,播种前将育苗器具用0.1%高锰酸钾溶液消毒,采用漂浮育苗,出苗前用烟草专用肥培养,到三叶一心期,以NH4NO3作为主要氮源,开始进行低氮(0.5 mmol/L,LN)处理和正常氮(5 mmol/L,NN)处理。
营养液配方主要参考前期筛选条件下的营养配方[17]。分别于处理后15 d和30 d各选取3株无损伤、无病害、生长状态一致烟苗的倒二叶或倒三叶、茎的中部和根的中部,在-80 ℃低温冰箱中保存,用于超氧化物歧化酶活性、丙二醛含量和脯氨酸含量的测定。
1.3 测定指标与方法
超氧化物歧化酶活性的测定采用氮蓝四唑(NBT)比色法;丙二醛含量的测定采用硫代巴比妥酸(TBA)比色法;脯氨酸含量的测定采用磺基水杨酸法。测定方法参考余前媛的方法[18]。
1.4 数据处理
采用Excel 2016和SPSS 20.0进行数据统计分析,采用Origin 8.5作图。
2 结果与分析
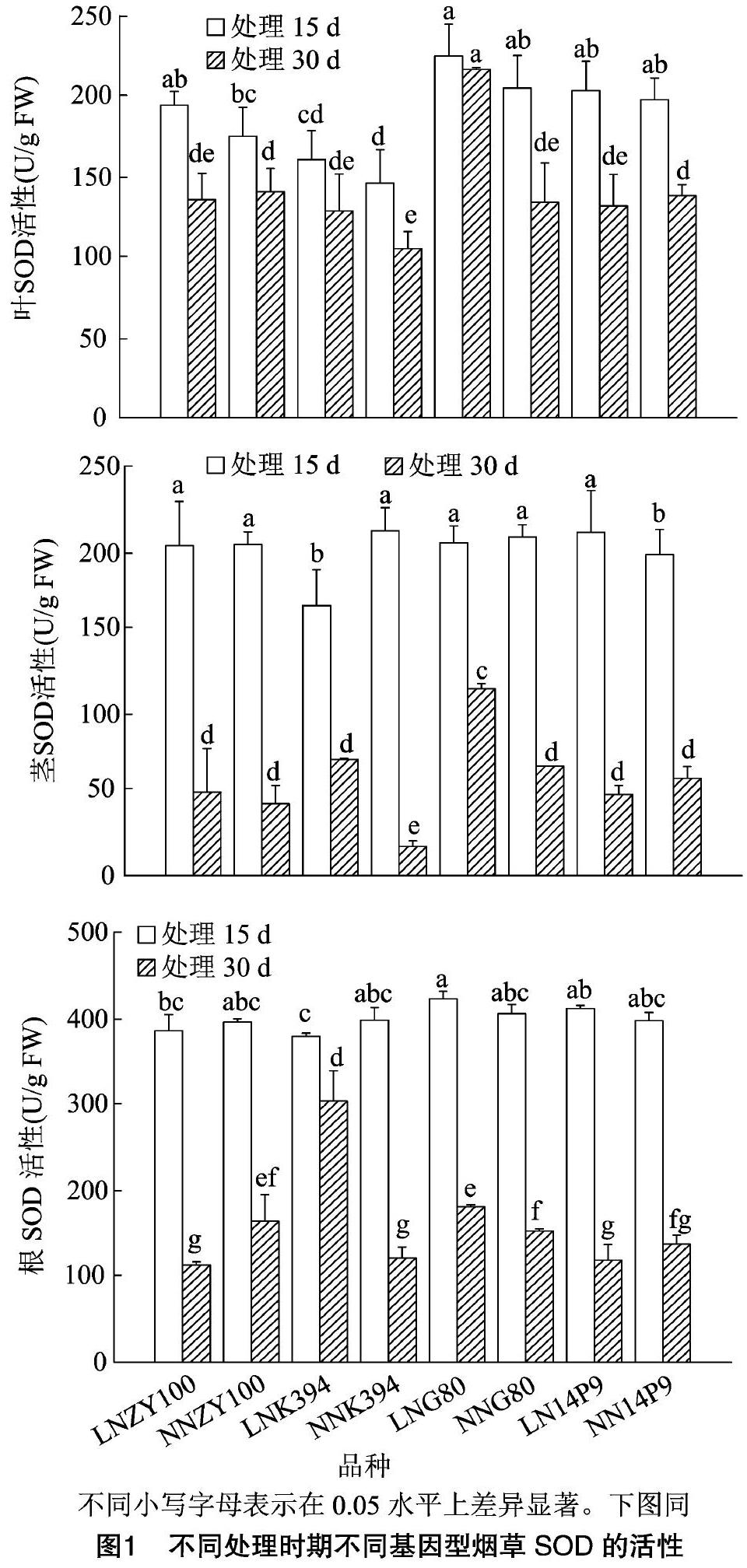
2.1 低氮胁迫时间对不同基因型烟草超氧化物歧化酶活性的影响
不同基因型烟草叶片(图1)中,中烟100正常氮处理与低氮胁迫下同一时期SOD活性均无显著差异,而低氮胁迫30 d下SOD活性则显著低于低氮胁迫15 d下活性;K394同一处理时期正常氮处理与低氮胁迫下SOD活性表现出无显著差异,低氮胁迫15 d与低氮胁迫30 d下SOD活性也无显著差异;G80低氮胁迫15 d和30 d下SOD活性无显著差异,处理15 d下低氮胁迫处理活性与正常氮处理活性无显著差异,处理30 d下LN处理>NN处理;14P9正常氮与低氮胁迫30 d SOD活性均显著低于正常氮与低氮胁迫下15 d活性,同一时期正常氮处理与低氮胁迫下活性表现为无差异。低氮胁迫15 d下,G80、14P9和中烟100的SOD活性无显著差异且均显著高于K394的活性;低氮胁迫30 d下,14P9、中烟100和K394的SOD活性也无显著差异但均显著低于G80的活性。
不同基因型烟草茎(图1)中,4个品种低氮胁迫30 d下SOD活性均显著低于低氮胁迫15 d下的活性;中烟100同一处理时期正常氮处理与低氮胁迫下SOD活性均无显著差异;K394处理15 d下LN处理NN处理;G80处理15 d下低氮胁迫与正常氮处理活性无差异,处理30 d下LN处理>NN处理;14P9处理15 d下LN处理>NN处理,处理30 d下低氮胁迫与正常氮处理下活性无显著差异。低氮胁迫15 d下,G80、14P9和中烟100的SOD活性无显著差异且均显著高于K394的活性;低氮胁迫30 d下,14P9、中烟100和K394的SOD活性也无显著差异但均显著低于G80的活性。
不同基因型烟草根(图1)中,中烟100低氮胁迫30 d下SOD活性显著低于低氮胁迫15 d活性,处理15 d低氮胁迫下与正常氮处理活性无显著差异,处理30 d下,LN处理NN处理;G80低氮胁迫30 d下SOD活性显著低于低氮胁迫15 d活性,低氮胁迫15 d下与正常氮处理活性无显著差异,处理30 d下LN处理>NN处理;14P9低氮胁迫30 d下SOD活性显著低于低氮胁迫15 d活性,同一处理时期正常氮处理与低氮胁迫下SOD活性均无差异。低氮胁迫15 d下,G80的SOD活性显著高于中烟100和K394,14P9的SOD活性与G80、中烟100无差异但显著高于K394;低氮胁迫30 d下,G80的SOD活性显著高于中烟100,却显著低于K394,14P9的SOD活性与中烟100无差异但显著低于G80、K394。
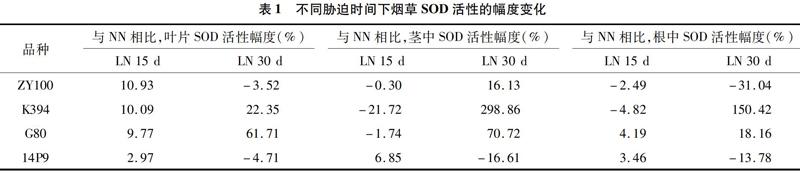
低氮胁迫15 d时,14P9叶片中的SOD活性升幅小于ZY100、K394,茎中仅14P9活性升高,其他3个品种活性均降低,其中G80降幅小于K394,根中14P9和G80活性升幅更大;低氮胁迫30 d时,14P9叶中活性降幅大于ZY100,G80升幅大于K394,茎中仅14P9活性降低,G80升幅大于ZY100,根中14P9降幅小于ZY100,G80升幅小于K394(表1)。
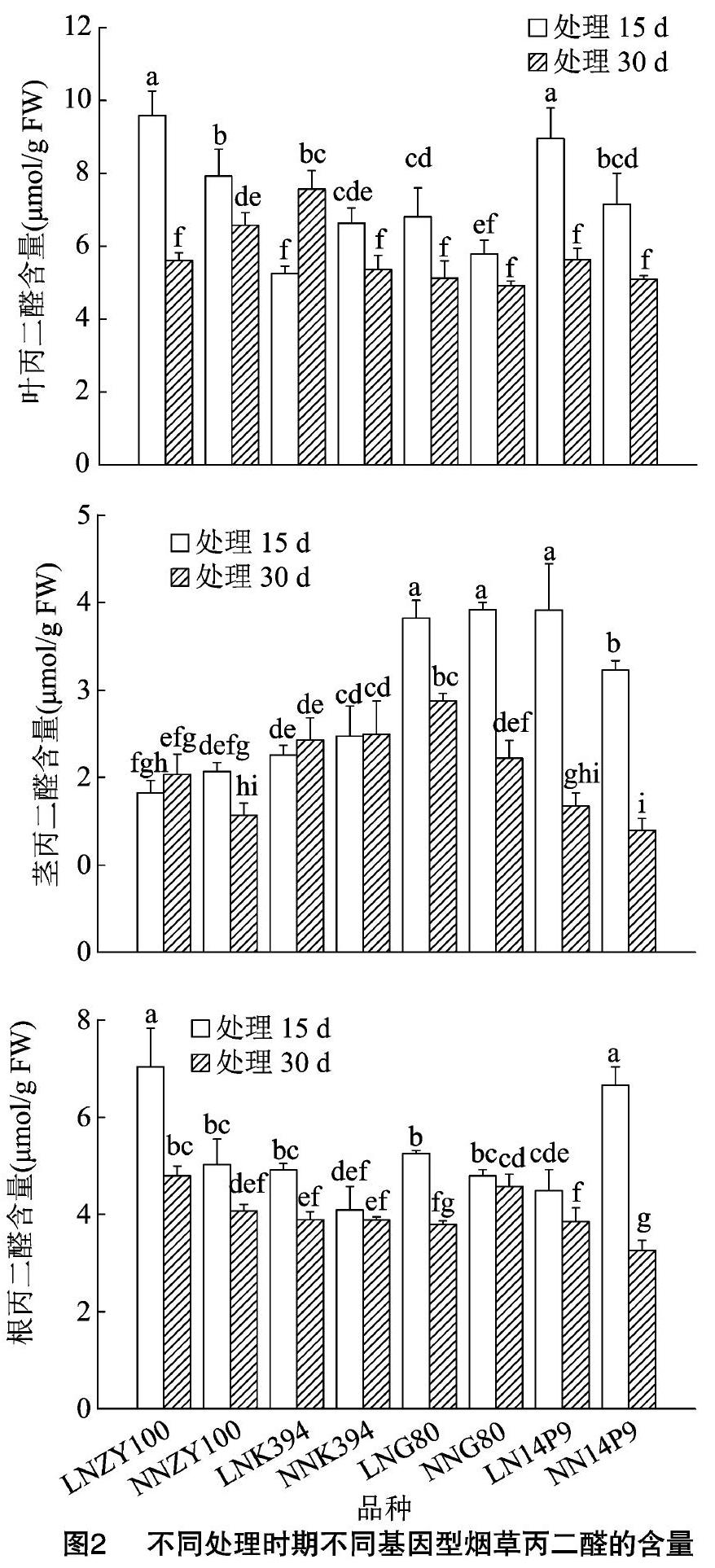
2.2 低氮胁迫时间对不同基因型烟草丙二醛含量的影响
不同基因型烟草叶片(图2)中,中烟100低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,处理30 d下LN处理NN处理;G80低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,低氮胁迫30 d下与正常氮处理30 d下丙二醛含量无差异;14P9低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,低氮胁迫30 d下与正常氮处理30 d下丙二醛含量无差异。低氮胁迫15 d下,G80的丙二醛含量显著小于中烟100却显著大于K394,14P9的丙二醛含量与中烟100无差异但显著大于K394、G80;低氮脅迫30 d下,G80、14P9和中烟100丙二醛含量无显著差异却显著小于K394的含量。
不同基因型烟草茎(图2)中,中烟100低氮胁迫30 d下丙二醛含量与低氮胁迫15 d下的含量无显著差异,低氮胁迫15 d下丙二醛含量与正常氮处理15 d的含量无显著差异,处理30 d下LN处理>NN处理;K394在不同氮处理和不同处理时期均表现为无显著差异;G80低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,低氮胁迫15 d下丙二醛含量与正常氮处理15 d下的含量无显著差异,处理30 d下LN处理>NN处理;14P9低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,低氮胁迫 30 d 下与正常氮处理30 d下丙二醛含量无差异。低氮胁迫15 d下,G80的丙二醛含量显著大于中烟100、K394,14P9的丙二醛含量显著大于中烟100、K394,与G80无显著差异;低氮胁迫30 d下,G80的丙二醛含量显著大于中烟100、K394,14P9丙二醛含量显著小于K394和G80的含量。
不同基因型烟草根(图2)中,中烟100低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,处理30 d下LN处理>NN处理;K394低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,低氮胁迫30 d下含量与正常氮处理30 d下的含量无显著差异;G80低氮胁迫30 d下丙二醛含量显著小于低氮胁迫15 d下的含量,低氮胁迫15 d下丙二醛含量与正常氮处理15 d下的含量无显著差异,处理30 d下LN处理NN处理。低氮胁迫15 d下,G80的丙二醛含量显著小于中烟100,与K394无显著差异,14P9的丙二醛含量显著小于中烟100、G80,与K394无显著差异;低氮胁迫30 d下,G80的丙二醛含量显著小于中烟100,与K394无显著差异,14P9丙二醛含量显著小于中烟100,与K394和G80的含量无显著差异。
低氮胁迫15 d时,14P9叶中丙二醛含量升幅大于ZY100、K394和品种G80,茎中仅14P9含量增加,其中G80降幅小于ZY100和K394,根中仅14P9含量降低,其中G80升幅小于ZY100和K394;低氮胁迫30 d,14P9升幅小于K394,大于G80,茎中14P9升幅小于ZY100和G80,根中14P9升幅大于ZY100和K394(表2)。
2.3 低氮胁迫时间对不同基因型烟草脯氨酸含量的影响
不同基因型烟草叶片(图3)中,中烟100低氮胁迫30 d下脯氨酸含量显著大于低氮胁迫15 d下的含量,处理15 d下LN处理>NN处理,处理30 d下LN处理>NN处理;K394低氮胁迫15 d下脯氨酸含量小于正常氮处理15 d下的含量,表现为无显著差异;G80低氮胁迫30 d下脯氨酸含量显著大于低氮胁迫15 d下的含量,低氮胁迫15 d下脯氨酸含量与正常氮处理15 d下的含量无显著差异,处理 30 d 下LN处理NN处理。低氮胁迫15 d下,G80的脯氨酸含量与中烟100、K394均无显著差异,14P9的脯氨酸含量显著小于K394,与中烟100、G80无差异;低氮胁迫30 d下,G80脯氨酸含量显著小于中烟100却显著大于K394,14P9与中烟100脯氨酸含量无显著差异,但显著大于K394、G80的含量。
不同基因型烟草茎(图3)中,中烟100低氮胁迫30 d下脯氨酸含量显著大于低氮胁迫15 d下的含量,处理15 d下LN处理NN处理;K394低氮胁迫30 d下脯氨酸含量显著大于低氮胁迫 15 d 下的含量,低氮胁迫15 d下脯氨酸含量与正常氮处理15 d的含量无显著差异,处理30 d下也表现出相同的趋势;G80低氮胁迫30 d下脯氨酸含量与低氮胁迫15 d下的含量无差异,低氮胁迫15 d下脯氨酸含量与正常氮处理15 d下的含量无显著差异,低氮胁迫30 d下LN处理NN处理。低氮胁迫15 d下,G80的脯氨酸含量显著大于中烟100、K394,14P9的脯氨酸含量显著小于G80,与中烟100、K394无显著差异;低氮胁迫30 d下,G80的脯氨酸含量显著小于中烟100,与K394无差异,14P9脯氨酸含量显著小于中烟100、K394,与G80的含量无显著差异。
不同基因型烟草根(图3)中,中烟100在不同氮处理和不同处理时期均表现为无显著差异;K394低氮胁迫30 d下脯氨酸含量显著小于低氮胁迫 15 d 下的含量,处理15 d下LN处理>NN处理,低氮胁迫30 d下含量与正常氮处理30 d下的含量无显著差异;G80低氮胁迫30 d下脯氨酸含量显著大于低氮胁迫15 d下的含量,低氮胁迫15 d下脯氨酸含量与正常氮处理15 d下的含量无显著差异,处理30 d下LN处理>NN处理;14P9同一处理条件下不同处理时期脯氨酸含量存在显著差异,同一处理时期不同处理条件也存在显著差异。低氮胁迫15 d下,G80的脯氨酸含量显著大于中烟100,与K394无差异,14P9的脯氨酸含量显著小于K394、G80,与中烟100无显著差异;低氮胁迫30 d下,G80的脯氨酸含量显著大于中烟100、K394,14P9脯氨酸含量显著小于G80,与K394和中烟100的含量不存在显著差异。
低氮胁迫15 d,耐低氮型烟草品种叶中丙二醛含量升幅大于低氮敏感型烟草品种,低氮胁迫30 d,耐低氮型烟草品种叶和茎中丙二醛含量升幅小于低氮敏感型烟草品种,根中升幅则大于低氮敏感型品种。丙二醛含量的变化与SOD活性的变化相对应,说明耐低氮型品种叶中低氮胁迫15 d时细胞膜质过氧化程度高,受到的伤害更大,随着胁迫时间的增加,则低氮敏感型品种的过氧化程度更大。
参考文献:
[1]王 勇,马 军,秦航道.氮对地枇杷幼苗生长及生理特征的影响[J]. 江苏农业科学,2019,47(23):181-185.
[2]裴 斌,张光灿,张淑勇,等. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J]. 生态学报,2013,33(5):1386-1396.
[3]朱世杨,刘 庆,张小玲,等. 盐胁迫对花椰菜苗期幼苗生物量及脯氨酸含量的影响[J]. 西南农业学报,2016,29(1):148-152.
[4]卢颖林,曾巧英,黄 莹,等. 低钾胁迫对斑茅抗氧化酶活性和脯氨酸含量的影响[J]. 甘蔗糖业,2018(3):9-14.
[5]刘龙元,贺鸿志,黎华寿. 水分胁迫对苦参生长生理及有效成分的影响[J]. 广东农业科学,2015,42(23):76-81.
[6]张 楚,张永清,路之娟,等. 低氮胁迫对不同苦荞品种苗期生长和根系生理特征的影响[J]. 西北植物学报,2017,37(7):1331-1339.
[7]Vendruscolo E C G,Schuster I,Pileggi M,et al. Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat[J]. Journal of Plant Physiology,2007,164(10):1367-1376.
[8]周艳虹,喻景权,钱琼秋,等. 低温弱光对黄瓜幼苗生长及抗氧化酶活性的影响[J]. 应用生态学报,2003,14(6):921-924.
[9]胡晓辉,郭世荣,李 璟,等. 低氧胁迫对黄瓜幼苗根系无氧呼吸酶和抗氧化酶活性的影响[J]. 武汉植物学研究,2005,23(4):337-341.
[10]张英华,杨佑明,曹 莲,等. 灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响[J]. 作物学报,2015,41(1):136-144.
[11]朴世领,刘 丹,安金花,等. 干旱胁迫及氮水平对烤烟生理特征的影响[J]. 河南农业科学,2006(11):43-47.
[12]李春燕,徐 雯,刘立伟,等. 低温条件下拔节期小麦叶片内源激素含量和抗氧化酶活性的变化[J]. 应用生态学报,2015,26(7):2015-2022.
[13]Trung D T,李 健,张风娟,等. 干旱胁迫下不同甘蔗品种叶片抗氧化酶活性和渗透调节物质含量的变化[J]. 热带作物学报,2018,39(5):858-866.
[14]林郑和,钟秋生,游小妹,等. 低氮对茶树生长及叶片抗氧化酶活性的影响[J]. 茶叶学报,2019,60(2):57-63.
[15]肖玉洁,李泽明,易鹏飞,等. 不同品种烟草响应低温胁迫生理生化差异分析[J]. 分子植物育种,2019,17(4):1346-1351.
[16]冯建灿,张玉洁,杨天柱.低温胁迫对喜树幼苗SOD活性、MDA和脯氨酸含量的影响[J]. 林业科学研究,2002,15(2):197-202.
[17]钟思荣,陈仁霄,陶 瑶,等. 耐低氮烟草基因型的筛选及其氮效率类型[J]. 作物学报,2017,43(7):993-1002.
[18]余前媛. 植物生理学实验教程[M]. 北京:北京理工大学出版社,2014.
[19]闫江艳,张永清,冯晓敏,等. 干旱胁迫及复水对不同黍稷品种根系生理特性的影响[J]. 西北植物学报,2012,32(2):348-354.
[20]李 强,罗延宏,龙文靖,等. 低氮胁迫对不同耐低氮性玉米品种苗期生长和生理特性的影响[J]. 草业学报,2014,23(4):204-212.
[21]謝孟林,李 强,查 丽,等. 低氮胁迫对不同耐低氮性玉米品种幼苗根系形态和生理特征的影响[J]. 中国生态农业学报,2015,23(8):946-953.
[22]杨中义,张发明,李永智,等. 不同烤烟品种对氮素胁迫响应差异的研究[J]. 云南农业大学学报,2011,26(2):240-245.
[23]肖国增,滕 珂,李林洁,等. 盐胁迫下匍匐翦股颖抗氧化酶活性及基因表达机制研究[J]. 草业学报,2016,25(9):74-82.
[24]姜慧芳,任小平. 干旱胁迫对花生叶片SOD活性和蛋白质的影响[J]. 作物学报,2004,30(2):169-174.
[25]赵福庚,刘友良. 胁迫条件下高等植物体内脯氨酸代谢及调节的研究进展[J]. 植物学通报,1999,16(5):540-546.
[26]赵婷婷,郑顺林,万年鑫,等. 早期施氮对马铃薯苗期抗旱能力的影响[J]. 干旱区资源与环境,2016,30(5):185-190.
[27]李 珣,刘中卓,纪薇薇. 施氮处理对不同株型水稻品种叶片渗透调节物质的影响[J]. 江苏农业科学,2019,47(15):117-121.闫军营,孙笑梅,程传凯,等. 腐殖酸与氮肥配施对豫北潮土冬小麦光合特性的影响[J]. 江苏农业科学,2020,48(21):104-110.
