氡对空气凤梨DNA 的辐射损伤效应
2020-03-03郑桂灵
郑桂灵 李 鹏 周 峰
1(青岛农业大学资源与环境学院 青岛266109)
2(南京晓庄学院食品科学学院 南京211171)
氡是铀(238U)衰变链中镭(226Ra)的衰变产物,半衰期为3.82 d。由于226Ra 广泛存在于岩石、土壤等各种地质环境中,所以,氡(222Rn)广泛存在于大气中,是自然界含量最丰富的天然放射性气体[1]。氡气本身在人体内驻留时间短,不会对人体健康造成很大危害。但是,氡衰变子体的性质与氡的性质完全不同,它们是重金属固体微粒,很容易和空气中的颗粒物质结合而变成结合态氡子体。氡子体被人体吸入后,衰变时产生的α粒子在体内照射,使得身体受害组织或细胞(主要是肺器官)发生电离化,破坏脱氧核糖核酸(DNA)的分子结构,影响细胞的再生过程,并引起细胞染色体的畸变,由此引发癌病变[2]。国际癌症研究机构(IARC)已将氡及其子体归为I 类致癌因素,世界卫生组织(WHO)也把氡作为19种人类致癌因子之一[3]。氡及其子体对于动物体的损伤已有很多研究。吸入氡子体后,大鼠上皮细胞的周期停滞,合成DNA 的细胞数明显减少[4]。Nie等[5]通过氡染毒大鼠尿液、外周淋巴细胞以及肺,结果发现,氡可以增加大鼠的氧化损伤,并抑制DNA修复酶8-羟基鸟嘌呤DNA 糖苷酶和8-羟基脱氧三磷酸鸟嘌呤核苷酶表达下降。Joster等[6]利用25.4 GBq 的流动氡气体照射源照射中国仓鼠卵巢细胞(Chinese hamsters ovary,CHO),在52 个氡诱发突变中,48%为基因丢失,23%为基因重组或失去1 个或几个外显子。之后的研究进一步表明,氡及其子体衰变产生的α粒子会造成一系列遗传上的改变,包括DNA 双链断裂、基因漂变、微核产生以及与之相关的蛋白表达等[7-11]。
氡对植物体是否像对动物体一样造成损伤尚罕有报道[12-13]。Villalobos-Pietrini等[14]通过实验表明,将Tradescantia花序经不同浓度的氡气处理后,在花粉母细胞减数分裂的四分体期观察到了微核的产生。我们的实验表明:松萝空凤(Tillandsiausneoides,T.usneoides)和贝可利空凤(Tillandsiabrachycaulos,T.brachycaulos)接受72 h的氡气薰蒸后遭受到了一定的生理损伤,如松萝空凤电导率值上升、贝可利空凤丙二醛(MDA)含量明显增加等;同时具有一定的抗性,如超氧化物歧化酶(SOD)含量在两种植物中均显著增加[15-16]。放射性物质对生物的伤害主要在于各种射线通过电离和激发对DNA 分子发生作用,使细胞受到损伤,从而导致各种危害[17],而氡气及其子体对植物DNA分子会产生什么影响一直未见报道。
我们继续以空气凤梨中的两种生态类型,即单株生长的贝可利空凤和丛生的松萝空凤做为实验材料,通过标准氡室对其进行不同氡浓度熏蒸处理后检测植物体的DNA 变化,以图阐明氡是否可对植物体产生显著影响,植物体受氡污染后哪些指标可反映这一影响的程度,并探讨了叶表鳞片在这一过程中所发生的作用,为研究氡与植物体的辐照损伤提供更多依据。
1 材料与方法
1.1 氡室设置
在北京大学的标准氡室中进行植物的氡气薰蒸处理。氡室由主腔室、氡源(226Ra)、测量系统(RAD7测氡仪,美国DURRIDGE公司)和其他配套设备组成;其容积约为1 m3,设有连接氡源(226Ra)的一条充氡气路,以及连接测氡仪的一条测量气路。氡室内配有小型风扇使腔室内氡浓度均匀。
1.2 植物氡气薰蒸
对单株生长的贝可利空凤,选取生长健康、大小相似的40株,将其随机分为4组。其中1组做为对照,置于正常室内环境中(氡平均浓度约为20 Bq/m3);1组人工去除其叶表鳞片后接受氡气的熏蒸,初始氡浓度为2 560 Bq/m3;另2组不去除叶表鳞片,直接接受氡气的熏蒸,初始氡浓度分别为2 879 Bq/m3和4 525 Bq/m3。
对于丛生的松萝空凤,称取3丛生长健康的松萝,每丛200 g,将其分为对照组和2 个实验组。对照组置于正常室内环境中,实验组初始暴露浓度分别为2530 Bq/m3和3 910 Bq/m3。
为减少植物的自身生长状态对实验结果的影响,实验前将植株浸泡于去离子水中20 min,使其吸水饱和,后在相同条件下统一晾30 min,以细线均匀悬挂于氡室中固定位置(图1)。封闭氡室后调节氡室起始氡浓度,对植物持续熏蒸72 h 后测其遗传指标。

图1 氡室中的空气凤梨:(a)贝可利空凤;(b)松萝空凤Fig.1 Tillandsia plants in the radon chamber:(a)T.brachycaulos;(b)T.usneoides
1.3 彗星实验
采用单细胞凝胶电泳技术(Single cell gel electrophoresis,SCGE),又称彗星实验(Comet assay)来检测氡气熏蒸后的植物DNA 损伤情况。由于该方法具有敏感、简便、快速、低耗、重复性好等独特优点,并可直接观察单个细胞内的DNA 损伤,从Ostling 等[18]采用SCGE 技术检测DNA 的双链断裂开始,该方法迅速成为近年常用的DNA损伤检测方法。
1.3.1 植物细胞核的提取
植物样品用双蒸水冲洗后剪碎,加入2%纤维素酶和果胶酶混合液,混匀后于4 ℃静置2 h,梯度离心收获,镜下观察细胞核形态完整,磷酸缓冲盐溶液(PBS)重复洗涤细胞2次,将细胞悬于PBS,调整细胞浓度至2×104mL−1,置于4 ℃冰箱备用。
1.3.2 碱性单细胞凝胶电泳实验
(1)铺胶:0.75%正常熔点琼脂糖凝胶煮沸后取100 μL迅速均匀铺于自制微电泳槽内,置于4 ℃冰箱1 min 使凝胶固化,取制好的细胞悬液25 μL 与75 μL 0.75%低熔点琼脂糖凝胶混匀后均匀铺于第一层凝胶上面,置于4 ℃冰箱固化1 min。(2)裂解:将微电泳槽置于新鲜配制的碱性裂解液中,于4 ℃冰箱中裂解2 h。(3)裂解后取出微电泳槽,用双蒸水漂洗去掉多余的盐分,置于提前加入4 ℃碱性电泳液的水平电泳仪中静置20 min,然后在20 V,200 mA 电泳条件下电泳20 min。(4)染色和观察:2 μg/mL溴化乙锭(EB)染色,双蒸水漂洗,去掉多余染液,先在荧光显微镜(Axioskop 2 mot plus,Carl Zeiss,德国)低倍镜下观察彗星,然后在高倍镜下,用图像采集系统随机抓取彗星图像,每份样品拍摄200个彗星细胞。
1.4 数据分析
每个彗星图像均采用波兰大学提供的CASP系统自动分析,测定彗星拖尾长度(Tail length)、彗星尾部DNA 相对含量(Tail DNA)、尾动量(TM)和Olive 尾动量(OTM)。对不同氡浓度处理后所测的遗传指标间的差异,采用SPSS 20.0 软件对数据进行单因素方差分析,p<0.05 视为差异有显著性。
2 结果与分析
2.1 氡对贝可利空凤叶片DNA 的辐射损伤
不同浓度氡处理后,贝可利空凤叶片细胞核电泳后获得彗星图像如图2 所示。由图2 可以看出,随着氡浓度增加,彗星尾部逐渐增长,表明DNA 迁移量增加。且尾部DNA 的含量随之增加。当氡浓度增加到4 525 Bq/m3时,从彗星图像上看仍然有明显的头部,但是DNA 迁移量明显增加(图2(d))。
以CASP 统计后,得到DNA 损伤程度的各种参数见表1。由表1 可以看出,与对照相比,被3种浓度的氡处理后,头部DNA 相对含量显著减少(p<0.05)。这一趋势在表征DNA 损伤程度的其他5个参数中更为明显,随着氡浓度增加,彗星长度、尾长、尾部DNA 相对含量、TM和OTM均显著增加(p<0.05),而且存在一定的剂量−效应关系,说明氡引起了贝可利空凤的DNA 损伤。
对去除叶表鳞片的植株而言(表1),虽然所施加的最初氡浓度较低(2 560 Bq/m3),但从彗星图像上可以看出,与对照相比,彗星尾部明显增长,DNA 迁移量明显增加(图2(c))。其DNA损伤指标值(表1)与不去除鳞片植物经高浓度(4 525 Bq/m3)氡处理后相近(p>0.05),远大于不去除鳞片植物经低浓度(2 879 Bq/m3)氡处理后的指标(p<0.05)。这说明没有叶表鳞片时,植株DNA 辐射损伤程度更高,而叶表鳞片的存在对氡的辐射起到了一定的防护作用。

图2 贝可利空凤叶片经氡处理后的彗星图像:(a)对照;(b)植物未去除鳞片,氡初始浓度为2 879 Bq∙m−3;(c)植物去除鳞片,氡初始浓度为2 560 Bq∙m−3;(d)植物未去除鳞片,氡初始浓度为4 525 Bq∙m−3Fig.2 Comet images for DNA damages in leaves of T.brachycaulos induced by radon:(a)control;(b)plants with foliar trichomes exposed to radon at initial concentration of 2 879 Bq∙m−3;(c)plants without foliar trichomes exposed to radon at initial concentration of 2 560 Bq∙m−3;(d)plants with foliar trichomes exposed to radon at initial concentration of 4 525 Bq∙m−3
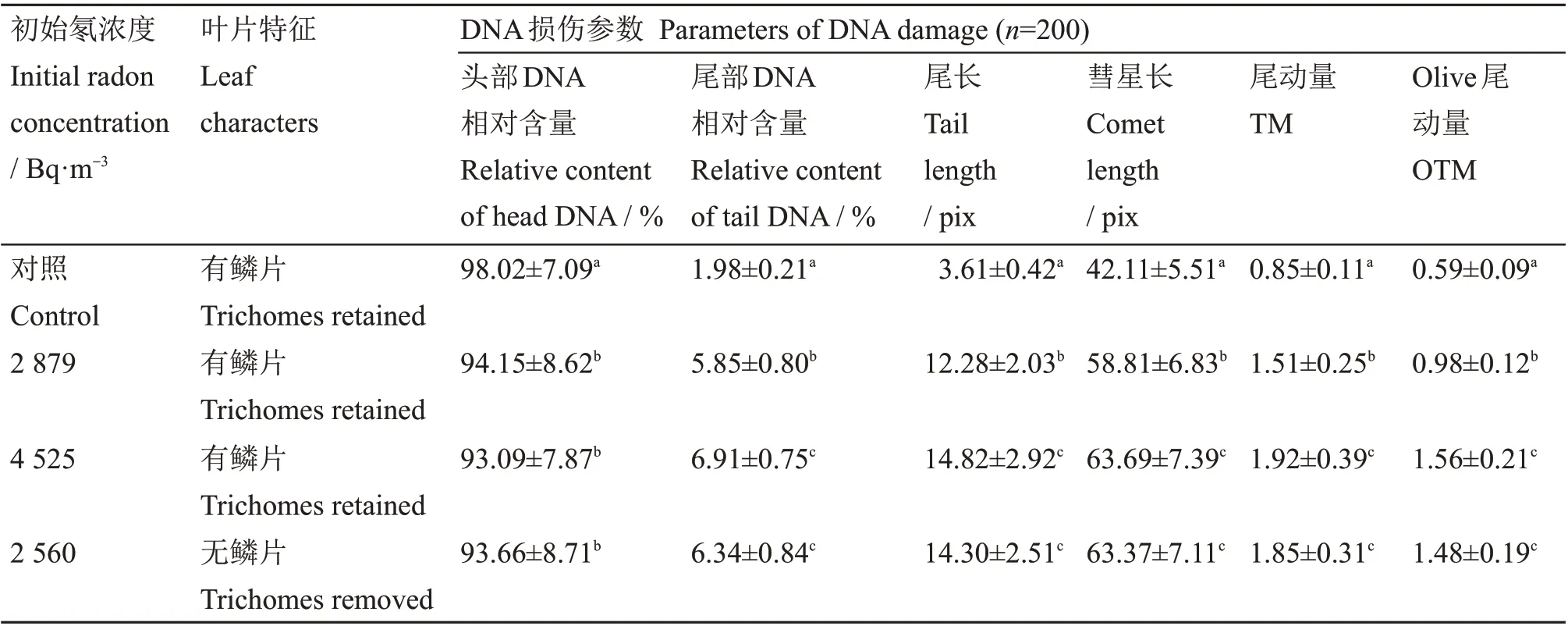
表1 贝可利空凤经氡处理后的SCGE分析Table 1 SCGE analysis of T.brachycaulos exposed to radon
2.2 氡对松萝空凤叶片DNA 的辐射损伤
经过不同浓度氡处理后,松萝空凤叶片DNA损伤状况与贝可利空凤类似。随着氡浓度增加,彗星尾部逐渐增长,表明DNA 迁移量逐渐增加(图3)。以CASP 统计表明,头部DNA相对含量明显减少,尾部DNA相对含量明显增加,彗星长度、尾长、TM 和OTM 的值也随之显著增加(p<0.05),这说明氡也引起了松萝空凤DNA 损伤并存在一定的剂量−效应关系(表2)。

图3 松萝空凤经氡处理后的彗星图像:(a)对照;(b)氡初始浓度为2 530 Bq∙m−3;(c)氡初始浓度为3 910 Bq∙m−3Fig.3 Comet images for DNA damages in T.usneoides induced by radon:(a)control;(b)treated at initial concentration of 2 530 Bq∙m−3of radon;(c)treated at initial concentration of 3 910 Bq∙m−3of radon

表2 松萝空凤经氡处理后的DNA损伤Table 2 DNA damage of T.usneoides exposed to radon
3 讨论
我们之前的研究已表明,空气凤梨可通过表面吸附、叶片吸收等作用有效吸收氡室中的氡,意味着氡可对植物进行体外辐照或内部辐照[19]。彗星实验所展示出来的DNA 指标(表1、2)表明,贝可利空凤和松萝空凤叶片DNA 确实不可避免地遭受了辐射损伤。在动物中,SCGE技术早已表明,随着大鼠暴露氡及其子体剂量的增加,外周血淋巴细胞、单个核细胞、肺细胞、支气管肺泡灌洗液细胞的DNA 迁移长度均逐渐增大,且有一定的剂量−效应关系[8-10]。植物在遭受辐照后也会产生类似情况,Saghirzadeh等[20]通过彗星实验发现高浓度的226Ra 对Alliumcepa的根细胞有巨大的影响。浓度1 000 μmol/L 的铀会导致Phaseolus根 细 胞DNA 几 乎 完 全 断 裂、 弥 散[21]。 当Tradescantia花粉母细胞经氡处理后,在其细胞分裂的四分体阶段出现了明显的微核现象[14]。因此,当空气凤梨叶片经氡处理后,其尾部DNA、尾长、TM 和OTM 的值随着氡浓度的增大而增加,且有明显的剂量−效应关系。这些指标也可以作为反映氡对植物损伤的敏感指标。
贝可利空凤去除鳞片实验也表明,空气凤梨对氡具有一定的抗性,而叶表鳞片在其中起着重要作用。因为在相近浓度氡胁迫下,人工去除鳞片的贝可利空凤DNA 损伤程度远远大于不去除鳞片的植株(表1)。实际上,做为常用的积累型指示植物,一个基本条件是必须具备的,即对污染物具有较强的抗性,不会受污染后很快致死。空气凤梨可以做为积累型指示植物的类群,已被广泛应用于Hg、Cu、Fe、Ni、Mn、Pb、V、Cs、Sr、Zn、PCB(多氯联苯)、PAH(多环芳烃)等多种大气重金属、核素和有机污染物的监测,其对各种污染物的较强抗性广为人知[22-26]。空气凤梨叶片表面覆盖有厚厚的鳞片,利用SEM 技术对松萝空凤吸收Hg、Pb、Cs、Sr 等重金属粒子的研究表明,绝大部分重金属颗粒被叶表鳞片所吸附[27]。也就是说,鳞片做为叶片最外层的保护结构,能有效防止污染物进入植物体内对植物产生伤害。虽然氡是一种辐射性气体,但其对生物体的影响除气态的222Rn 外,主要是其各种粒子态的子体,如钋218(218Po)、铅210(210Pb)、铋210(210Bi)、钋210(210Po)等,它们均主要以类似重金属的形式存在。因此,空气凤梨对氡也具有一定的抗性是不难理解的。
另外,即使有少量氡能进入植物内部,抗氧化酶系统的存在也能进一步减轻氡对植物的伤害。抗氧化酶是植物活性氧自由基清除剂,当植物受到逆境胁迫时,这些保护酶的活性会增强,从而可以有效清除体内的自由基,使膜系统减轻因自由基而引发的过氧化作用伤害,保持膜结构及功能的稳定[28-29]。之前的实验已表明,贝可利空凤和松萝空凤的SOD活性随着氡浓度的增加显著升高,说明在氡胁迫下,松萝体内活性氧清除体系开始发挥作用,这在一定程度上能减轻氡对植物的伤害,增强空气凤梨对氡的抗性[15-16]。
