日本梅花鹿与大丰麋鹿发情期反刍行为的差异
2020-02-27张宜贵UllahSanaTakasuFugo李忠秋
张宜贵, Ullah Sana, Takasu Fugo, 李忠秋*
(1. 南京大学生命科学学院动物行为与保护实验室,南京210023; 2. 奈良女子大学环境科学学部,日本奈良630-8506)
反刍是指食草动物进食一段时间后将半消化的食物从胃(一般是瘤胃)里返回嘴里再次咀嚼。反刍包含2个过程:1)食团从瘤胃中逆呕至口中;2)对逆呕出来的食团进行咀嚼再吞回瘤胃(Moquinetal.,2010;Li,2013;Wangetal.,2018)。食草动物的反刍行为对其充分消化有重要的作用,因为反刍动物在采食时都是匆忙进食,大部分食物未经充分咀嚼就吞咽进入瘤胃,而反刍可以通过反复咀嚼把食物分解成更小的颗粒以便食物在皱胃中被消化吸收(Wilson,1994;Pérez-Barbería & Gordon,2010)。
研究表明,一些内外因素如体型、觅食地、天气等均可影响食草动物的反刍行为(Blanchard & Fritz,2008;Moquinetal.,2010;Pérez-Barbería & Gordon,2010)。其中体型被认为是影响反刍行为的最重要因素(体型较大的个体会增加反刍频率或反刍次数、降低咀嚼速率等),而体型通常与个体的性别和年龄具有密切联系(Toit & Yetman,2005;Pérez-Barbería & Gordon,2010),因此,本文采用性别和年龄表征种间个体体型。Jarman-Bell原理表明,体型较大的动物由于新陈代谢需求低与肠道容量比例大可以以高纤维含量的低质量食物为食(Jarman,1974)。研究指出,新陈代谢需求、肠道容量和瘤胃体积与体型相关(Bell,1971;Jarman,1974;Pérez-Barberíaetal.,2008),即体型较大的动物能够使食物停留在瘤胃的时间更长,因此比体型较小的动物消化吸收更多的营养。而体型较小的动物从低质量的食物中(例如高纤维含量的食物)汲取营养的能力不占优势。同种动物可能会因性别、年龄不同而采取不同的反刍或咀嚼对策去最大化吸收能量和营养。
为了更好地研究反刍行为,研究者已经提出一系列特定的参数描述反刍行为,例如反刍时间/休息时间、食团咀嚼时间、单个食团咀嚼次数(咀嚼频率)以及单位时间咀嚼次数(咀嚼速率)(Pérez-Barbería & Gordon,2010;Li,2013)。麋鹿Elaphurusdavidianus为我国的特有珍稀鹿科Cervidae动物,Li(2013)较为系统地研究了体型因素即性别年龄对麋鹿反刍行为的影响,而体型因素是否在具有体型差异的物种种间也发挥作用尚不清楚。2018年 9月本研究组应邀到日本奈良进行学术交流。奈良的日本梅花鹿Cervusnipponnippon种群大且不惧人,方便行为学观察,其体型小于麋鹿,且体型具有明显的年龄性别差异。因此,利用相同的观察方法对日本梅花鹿进行反刍行为研究,试图比较麋鹿和日本梅花鹿反刍行为的种间及种内差异。
1 研究方法
1.1 研究地点和研究对象
日本梅花鹿的反刍数据在2018年9月收集于日本奈良市奈良公园(135°50′34.9″E,34°41′06.7″N);麋鹿的反刍数据在2012年7—8月收集于中国江苏大丰麋鹿国家级自然保护区(120°48′42.00″E,32°59′35.50″N)。
日本梅花鹿成年个体体长约150 cm,成年雄鹿体质量约150 kg,成年雌鹿100~120 kg,仅雄鹿有角,成年雄鹿角一般有4个分叉(鲜有5个分叉),全身有明显的白色斑点(宋延龄,刘志涛,2005)。麋鹿成年个体体长约200 cm,成年雄鹿体质量约180 kg,成年雌鹿约140 kg,亚成雄性约130 kg,亚成雌性约100 kg,幼体约13 kg(蒋志刚,丁玉华,2011),仅雄性有角。
研究期间,记录每只被观测样本的性别、年龄,并根据体型、体长、体高以及角的形状和大小分为5个类型:幼体(体小,无角,不能明显区分雌雄)、亚成雄性(有角但未分叉)、亚成雌性(体型约为成年的2/3,无角)、成年雄性(角粗大,分两叉或两叉以上,体长1.60~1.73 m)和成年雌性(无角,体长仅次于成年雄性)(戚文华等,2010;Li,2013)。
1.2 行为取样及数据分析
日本梅花鹿的行为采用焦点取样法在奈良公园草地上进行,研究者在10 m以外进行观测以降低对目标动物的扰动。虽无法进行个体识别,也未对目标动物标记,但公园内鹿群数量较大,且选取不同的观测地点,基本排除了同一个体被重复取样的可能性;麋鹿反刍数据收集(行为取样方法同日本梅花鹿)和整理由本研究组在2012年完成(Li,2013)。
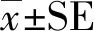
共收集发情期的77只日本梅花鹿个体和104只麋鹿个体的卧息反刍数据。其中,日本梅花鹿幼体24只,亚成雄性10只,亚成雌性17只,成年雄性13只,成年雌性13只;麋鹿幼体13只,亚成雄性24只,亚成雌性19只,成年雄性34只,成年雌性14只。
数据经验证满足正态分布,采用单因素方差分析(One-Way ANOVA)验证了性别-年龄(幼体、亚成雄性、亚成雌性、成年雄性、成年雌性)对日本梅花鹿和麋鹿反刍行为的影响,同时分析比较了两者反刍行为差异,探索种内和种间不同的反刍对策和咀嚼对策。数据分析使用SPSS 20.0,显著水平设置为0.05。
2 结果
对日本梅花鹿的研究表明,性别-年龄对咀嚼速率有极显著影响(F4,72=5.230,P=0.001),咀嚼速率为幼体>亚成雌性>亚成雄性>成年雌性>成年雄性;性别-年龄对食团咀嚼时间有显著影响(F4,72=3.313,P=0.015),食团咀嚼时间为成年雄性>亚成雄性>成年雌性>亚成雌性>幼体;未发现性别-年龄对咀嚼频率有显著影响(F4,72=1.810,P=0.136)(表1)。
对麋鹿的研究结果表明,性别-年龄对咀嚼频率(F4,99=4.770,P=0.001)有极显著影响,咀嚼频率为幼体>亚成雌性>亚成雄性>成年雌性>成年雄性;未发现性别-年龄对食团咀嚼时间(F4,99=1.661,P=0.165)和咀嚼速率(F4,99=2.121,P=0.084)有显著影响(表2)。

表1 性别-年龄对日本奈良日本梅花鹿反刍参数的影响Table 1 Effect of sex-age on the rumination parameters of Cervus nippon nippon in Nara Park, Japan
注: 同一列相同字母表示差异没有统计学意义; 表2同
Notes: the same letter in the same column indicates there is no significant difference; the same as table 2

表2 性别-年龄对大丰麋鹿反刍参数的影响Table 2 The effect of sex-age on rumination parameters of Elaphurus davidianus in Jiangsu province, China
日本梅花鹿和麋鹿的反刍行为在咀嚼频率上的差异有统计学意义(F1,179=4.336,P=0.039),在食团咀嚼时间(F1,179=26.391,P<0.001)和咀嚼速率(F1,179=56.414,P<0.001)上的差异有高度统计学意义(表3)。

表3 日本梅花鹿和大丰麋鹿反刍参数的对比Table 3 Comparison of the rumination parameters between Cervus nippon nippon in Nara Park, Japan and Elaphurus davidianus in Jiangsu province, China
3 讨论
本研究结果表明,性别和年龄显著影响奈良日本梅花鹿的咀嚼速率和食团咀嚼时间及大丰麋鹿的咀嚼频率,即性别-年龄在种内一定程度上影响了2种鹿科动物的反刍行为。以往研究表明,个体的性别和年龄可以影响反刍参数(Oakesetal.,1992;Dasetal.,1999;Melladoetal.,2005;Li,2013),同时指出性别-年龄的影响与体型的影响非常相似(Grossetal.,1995;Toitetal.,2005;Pérez-Barberíaetal.,2008;Moquinetal.,2010),即体型通常和性别-年龄有关。在对人类咀嚼效率的研究中发现,当控制体型时,性别和年龄对人类咀嚼效率的影响也随之消失(Grossetal.,1995)。这是因为体型与决定牙齿有效性的咬合面积存在一定的关系(Shipleyetal.,1994;Pérez-Barbería & Gordon,1998;Melladoetal.,2005)。因此,个体可以通过改变咀嚼行为补偿牙齿有效性的降低,进而提升咀嚼效率。
本研究中日本梅花鹿的咀嚼速率按幼体、亚成雌性、亚成雄性、成年雌性、成年雄性递减,且性别-年龄对咀嚼速率有极显著影响。行为取样时,观察到当在同一片草地采食时,体型较小的个体不停更换采食站以获得高质量的食物,而体型较大的个体则在同一采食站采食更长时间。这与Ruckstuhl(1998)的研究结果一致,即体型较小的雌性大角羊Oviscanadensis在采食时的平均移动距离是体型较大雄性的4倍。
日本梅花鹿随体型的增大,采取了不同的反刍和咀嚼对策,即在幼体时采食高质量的食物(低纤维),咀嚼速率高,食团咀嚼时间短;成年个体采食低质量的食物(高纤维),咀嚼速率低,食团咀嚼时间长。相同年龄的雌鹿比雄鹿的咀嚼速率更大,食团咀嚼时间更短。Pérez-Barbería等(2008)研究指出,Jarman-Bell原理可以解释有蹄类草食动物性别分离的出现,即两性个体在食物消化率上存在差异。本研究结果也为Jarman-Bell原理在解释有蹄类草食动物体型的性别差异上提供了有力的证据,体型更大的雄性通过一定的反刍对策能比雌性更好地消化高纤维的低质量食物。
事实上,体型对反刍行为的影响亦表现在种间水平上(Pérez-Barberíaetal.,2008)。本研究比较了大体型麋鹿和小体型日本梅花鹿的反刍行为差异。结果表明,两者在咀嚼频率、食团咀嚼时间和咀嚼速率上均有显著差异,体型更大的麋鹿咀嚼速率更小、食团咀嚼时间更长。由此可见,日本梅花鹿和麋鹿采取了不同的反刍和咀嚼策略最大化吸收营养。本研究结果与Jarman-Bell原理最初用来解释不同体型的非洲有蹄类动物采食不同纤维含量的食物相吻合,即体型更大的麋鹿会比体型较小的梅花鹿采食纤维含量较高的食物,如四川梅花鹿Cervusnipponsichuanicus多喜采食植物的芽、枝梢、嫩枝叶等(郭延蜀,2001),而麋鹿则可以采食干草或枯枝(王轶,王文,2011)。对同域分布的有蹄类食草动物而言,体型较大的物种由于低新陈代谢需求与大肠道容量比例可以以低质量的食物(高纤维含量)为食,而体型较小的物种则由于高新陈代谢需求与小肠道容量比例以高质量的食物(低纤维)为食,从而达到共存。这将为建立混合种群的珍稀有蹄类草食动物自然保护区提供思路。
种内种间的反刍行为差异,可能都是体型效应的一个缩影。反刍行为体型效应的产生,可能与其食性及食物选择有关(Pérez-Barbería & Gordon,2010)。受代谢率和进食能力的影响,体型较小的动物倾向于挑选少量的高质量(低纤维)的食物,而体型较大的动物则倾向于挑选大量的低质量(高纤维)的食物(Jarman,1974)。当较大的动物消化低质量的食物时,低咀嚼速率和长时间咀嚼可以提高咀嚼效率(Soestetal.,1994;Li,2013)。因此体型可能是影响反刍行为最重要的因子之一。
日本梅花鹿的反刍行为还可以作进一步的研究,例如觅食地、天气等对反刍的影响及其影响机制;探索繁殖期和非繁殖期反刍行为是否存在差异;不同性别-年龄的日本梅花鹿在休息时用于反刍的时间是否存在差异等。这将有助于进一步了解有蹄类动物的反刍行为,并为珍稀有蹄类动物保护、驯化及管理提供思路。
致谢:南京大学动物行为与保护实验室的王琳、罗云超、汪开宝、李雨航、万月、王鑫鑫、武亦乾、常浩等同学参与了野外调查,在此一并致谢。
