不同种子大小籽瓜品种叶片光合能力对源库调节的响应
2020-02-03陈丽婷陈年来
李 雨,陈丽婷,陈年来
(1.甘肃农业大学园艺学院,甘肃 兰州 730070;2.甘肃农业大学资源与环境学院,甘肃 兰州 730070)
源和库是作物经济产量形成的两个重要方面,通常用源和库的发展、协调、平衡等探讨作物的高产途径[1]。源库学说认为,作物产量的形成必须具备两个条件:一是要有足够强的源器官生产光合产物,二是要有充足的库器官接纳光合产物[2]。源、库单位的可塑性是园艺作物生产中实施摘叶整枝、疏花疏果等栽培技术的生理基础[3]。通过农艺措施改变作物固有的源库比例和协调性,可使植株在最佳光能利用与合理的营养分配状态下生长,既达到高产、高效的目的,又能提高产品品质[4-7]。
源、库器官之间具有功能互馈调节作用,源、库协调高效是作物高产的基本途径。库需求量过大,源器官负担重,易导致叶片衰老,此时减库或增源能够提高产量。疏花疏果可以提高叶片叶绿素含量和净光合速率,延缓叶片衰老,而减源则会进一步增加源器官负担,促进叶片衰老,导致光合能力降低[8]。源库比过大,大部分叶片因功能冗余或相互遮阴难以充分发挥光合潜力,此时减源或增库有利于产量提高。研究发现,在生长发育前期去叶处理使小麦、玉米、油茶叶绿素含量与净光合速率增加[9-11]。摘除叶片后,同化物供应源减少,所留叶片的光合产物不能满足库的需求,加重源器官负荷,致使叶绿素加速降解,叶片衰老加快[1,12]。甜瓜单蔓整枝和留瓜数较多的处理,果实发育后期植株早衰较为严重[7,13]。同时,源库比较小的群体叶面积指数一般较小,易造成漏光损失,不利于光能截获和产量形成;而源库比过高的群体叶片密度太大,易造成田间郁闭,冠层通风透光性能差,叶片光合效能不高,亦不利于高产[14]。
籽瓜是以种子为主要产品的作物,其库器官既包括果实也包括种子,即果实数目和大小、单个果实内的种子数目和大小均可能对叶片光合能力具有调节作用。目前关于籽瓜源库关系的研究较少,对于不同源库比条件下籽瓜源活力的研究未见报道。本试验以种子大小具有显著差异的3个籽瓜品种为材料,通过调节叶果比控制果实发育期间源库数目比,探讨源库比对籽瓜叶片光合能力的影响,为籽瓜优质高产生产中种植密度的确定提供理论依据。
1 材料与方法
1.1 试验材料
供试籽瓜材料为种子大小显著不同的3个品种,H3为小种子品种,千粒重120±5 g,单果重0.88 kg,全生育期115 d;H14为中等种子品种,千粒重240±20 g,单果重1.25 kg,生育期130 d;H26为大种子品种,千粒重400±50 g,单果重1.2 kg,生育期135 d。3个品种果皮色泽均为绿底色核桃纹,种子色泽为黑边白心。种子均为2018年采集,由甘肃农业大学瓜类研究所提供。
1.2 试验方法
试验于2019年5—9月在甘肃省民勤县进行,垄沟种植,行株距为1.1 m×0.3 m。当地土壤为砂质土,有机质8.25 g·kg-1,全氮、全磷、全钾含量分别为0.48、0.66、21.86 g·kg-1。生育期内平均降雨量19.38 mm,平均太阳辐射强度14.64 kJ·m-2·min-1,日均温度21.15℃,大气相对湿度46.61%。3个籽瓜品种均于开花坐果初期通过整枝疏果将单株叶果数目比调节为10(单蔓整枝,10片叶时摘心),20、30(双蔓整枝,在10或15片叶时摘心)和40、50(三蔓整枝,在13~17片叶时摘心),之后随时摘除其他新发生侧蔓,每株留1个果实(其他果实在坐定后摘除)。每个处理3次重复,每个重复种植25株,共45小区。试验田其他管理与生产田相同。
1.3 测定指标及方法
叶绿素的测定:于幼果期、果实膨大期、果实成熟期,每小区选择10株植株,用手持式叶绿素仪(SPAD-502型)测定其坐果节位叶片的叶绿素浓度,每个叶片记录3~5个稳定数值。
气体交换参数的测定:于幼果期、果实膨大期、果实成熟期,每小区选择1株植株,用CIRAS-2型便携式光合测定系统(PP Systems,UK)测定晴天上午8∶00—11∶00坐果节位及其前后各1片叶片的Pn、Gs、Ci、Tr,每个叶片记录3~5个稳定数值。
果实产量:采收后每个小区取果实15个,称量其重量。
1.4 数据分析
用Excel 2010软件对数据进行处理及作图,采用SPSS 23.0软件进行数据分析。
2 结果与分析
2.1 籽瓜叶片叶绿素含量对叶果比的响应
利用光谱吸收原理测定的SPAD值与叶绿素含量呈正相关,能够反映作物叶片的叶绿素相对浓度。3个籽瓜叶片SPAD值均随果实生育期演进逐渐降低,幼果期叶片SPAD均值略高于果实膨大期,膨大期SPAD值显著高于果实成熟期(图1)。幼果期至果实膨大期,SPAD均值降幅较小(3.7%),果实膨大期至成熟期降幅较大(6.9%),表明果实生育后期叶绿素含量下降更快。3个籽瓜品种间幼果期和果实膨大期叶片SPAD值差异显著,大种子品种H26叶片平均SPAD值(66.2)显著大于中等种子品种H14(65.2),H14的SPAD值显著大于小种子品种H3(64.6);果实成熟期品种间差异消失或改变。3个品种果实生育期叶片SPAD生育期平均值降幅略有差异,H26降幅(11.6%)大于H14(10.4%),H14大于H3(9.3%)。各时期3个籽瓜品种功能叶SPAD均值随叶果比增大而显著降低,但叶果比40与50间差异不显著,生育期平均SPAD值降幅也随叶果比增大而减小(叶果比50除外)。叶果比10(13.4%)>20(11.8%)>50(11.1%)>30(8.3%)>40(7.2%),说明一定范围内摘叶能增加叶片叶绿素含量,但摘叶处理或留叶数过多均易造成叶绿素加速降解。

注:不同大写字母表示相同品种不同叶果比处理间差异显著(P<0.05),不同小写字母表示相同叶果比处理不同品种间差异显著(P<0.05),下同。Note: Different uppercase letters indicate the significant difference between different leaf and fruit ratio treatments of the same variety (P<0.05), and different lowercase letters indicates the difference between different leaf and fruit ratio treatments of the same variety (P<0.05), the same below.
2.2 籽瓜叶片气体交换特性对叶果比的响应
2.2.1 净光合速率对叶果比的响应 3个籽瓜品种叶片净光合速率(Pn)随果实生育期演进逐渐降低,幼果期叶片Pn显著高于果实膨大期,膨大期Pn显著高于果实成熟期(图2)。幼果期至果实膨大期Pn平均降幅(28.8%)与果实膨大期至成熟期平均降幅(29.7%)相仿。3个品种在3个发育时期叶片Pn差异显著,小种子品种H3(21.3 μmol·m-2·s-1)叶片平均Pn显著大于中等种子品种H14(20.2 μmol·m-2·s-1),H14的平均Pn显著大于大种子品种H26(19.2 μmol·m-2·s-1)。3个品种果实生育期叶片Pn平均降幅H26(51.38%)>H14(49.6%)>H3(48.16%)。3个品种功能叶Pn均值随叶果比增大而显著降低,3个生育期的平均Pn降幅也随叶果比增大而显著降低(叶果比50除外)。叶果比10(58.27%)>20(52.13%)>30(49.67%)>50(46.23%)>40(42.27%),说明一定范围内摘叶能增加叶片净光合速率,但摘叶或留叶数过多均易造成叶片光合性能下降,净光合速率降低。
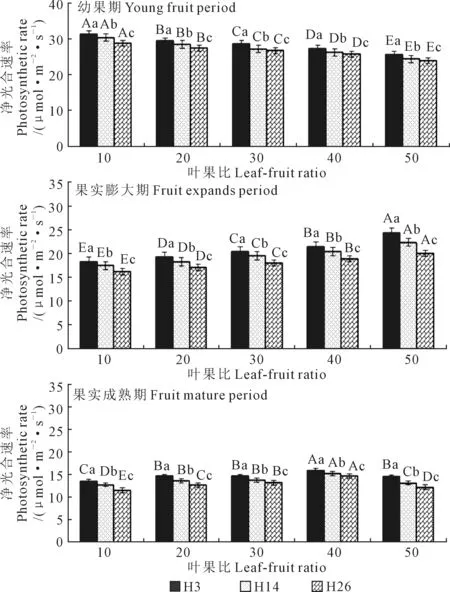
图2 不同叶果比处理的籽瓜叶片净光合速率
2.2.2 气孔导度对叶果比的响应 籽瓜叶片气孔导度(Gs)随生育期演进而降低,幼果期叶片Gs均值(0.71 mmol·m-2·s-1)显著高于果实膨大期(0.44 mmol·m-2·s-1),果实膨大期叶片Gs均值略高于果实成熟期(0.36 mmol·m-2·s-1)(图3)。幼果期至膨大期叶片气孔导度平均降幅为38.3%,大于膨大期至果实成熟期的气孔导度平均降幅(18.3%),说明在果实生育前期气孔导度下降速率更快。3个品种叶片生育期平均Gs大小为 H3(0.55 mmol·m-2·s-1)>H14(0.50 mmol·m-2·s-1)>H26(0.46 mmol·m-2·s-1),叶片Gs在果实各生育期的平均降幅为H3(48.9%)

图3 不同叶果比处理的籽瓜叶片气孔导度
2.2.3 蒸腾速率对叶果比的响应 随着生育期演进,3个籽瓜品种叶片的蒸腾速率(Tr)逐渐降低,幼果期叶片Tr均值显著高于果实膨大期,膨大期Tr均值显著高于果实成熟期(图4)。幼果期至膨大期叶片Tr的平均降幅(31.0%)略大于膨大期至成熟期(28.3%),表明相邻两个时期间蒸腾速率下降速率相似。3个品种间小种子品种H3的叶片平均Tr(10.4 mmol·m-2·s-1)略大于中等种子品种H14(9.8 mmol·m-2·s-1),H14的叶片平均Tr略大于大种子品种H26(9.1 mmol·m-2·s-1),叶片Tr在整个果实生育期的降幅H3(48.9%)
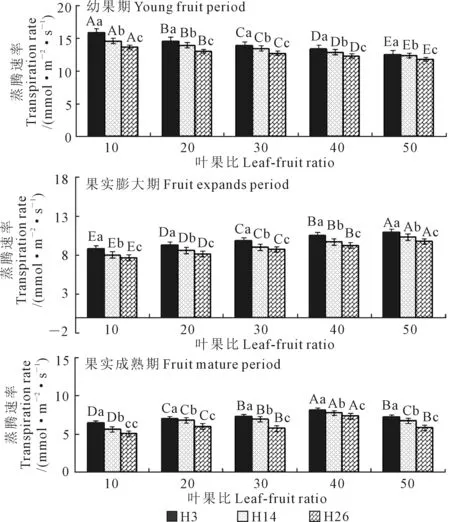
图4 不同叶果比处理的籽瓜叶片蒸腾速率
2.2.4 胞间CO2对叶果比的响应 从幼果期至果实膨大期,3个籽瓜品种叶片胞间CO2浓度(Ci)随着生育时期演进而显著升高,果实成熟期叶片平均Ci显著高于果实膨大期,膨大期叶片平均Ci显著高于幼果期(图5)。叶片Ci平均增幅幼果期至膨大期(10.3%)<膨大期至果实成熟期(14.1%),即在果实生育后期叶片Ci升高更快。3个品种间3个发育时期叶片平均Ci差异显著,H3(255.6 μmol·mol-1)
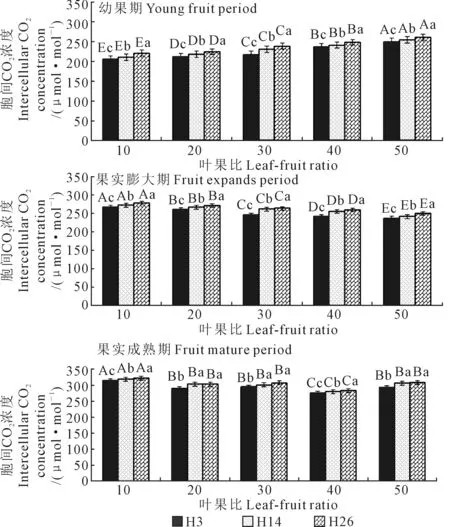
图5 不同叶果比处理的籽瓜叶片胞间CO2浓度
2.3 籽瓜果实产量对叶果比的响应
籽瓜果实产量随叶果比的增大依次增大(图6),不同叶果比的果实产量表现为:叶果比10(0.867 kg)<20(1.163 kg)<30(1.170 kg)<40(1.172 kg)<50(1.177 kg),方差分析表明,果实产量在叶果比10时最小,在20~50的叶果比间差异不大。3个品种的果实产量H3(0.88 kg)

图6 不同叶果比处理的籽瓜果实产量
3 讨 论
在瓜类作物生产中,对整个叶系统年龄结构调整主要是通过整枝来实现的,整枝就是通过减少无效或低效叶的发生,调节有机养分的流向与流量,从而间接影响整个叶系统的光合能力与寿命[15]。因此,在生产中采取修剪、摘叶、疏花疏果等措施改变光合同化产物在库与源间的均衡分配,从而对产量和果实品质产生重要影响[16]。
3.1 籽瓜叶片光合能力对源库调节的响应
植株叶片是光合作用的主要器官,光合作用是作物产量形成的基础,叶片光合速率与作物产量呈正相关[17]。库与源的关系改变影响果树光合作用及光合同化产物在库、源器官间的运输与分配,进而影响果树的经济产量[18]。在一定范围内,叶面积越大,叶片光合产物越多。叶面积取决于叶片数目与叶片大小,其中叶片大小对于同期植株来说差异较小,而叶片数的多少变化较大[19]。对于紧凑型作物植株来说,适当减少源叶片,避免叶片冗余,使得作物群体透光性能增强,有利于光合作用加强;但源的过度减少会使叶片群体光合势及光合生产率下降,反而降低产量[20]。另外也有研究证实,低库强下叶片中积累的光合同化物是造成净光合速率下降的原因[21-22]。唐瑞永[23]在甜瓜作物上的研究发现,在幼果期随着叶果比的增加,叶片的净光合速率和叶绿素浓度均降低。本研究结果与此一致,并且籽瓜叶片Gs、Tr在幼果期均随着源库比的增大而降低,而Ci随着源库比的增大而升高。在果实发育后期,叶果比50的处理叶绿素含量最低,主要是由于叶片较多,互相遮阴,造成叶绿素降低;叶片的净光合速率、气孔导度、蒸腾速率在叶果比10时最低,在叶果比40时最高,说明摘叶过多使叶片在果实发育后期光合能力下降最大;而胞间CO2浓度与此相反,在叶果比10时最高,叶果比40时最小。对果实产量的研究发现,叶果比10的果实产量最低,其余叶果比之间产量无明显差异,说明减源过度造成叶片光合能力下降,从而导致减产。本研究还发现小种子的果实产量显著小于中等种子大小和大种子的产量。
3.2 籽瓜叶片衰老对源库调节的响应
衰老是生物个体或器官生理机能随年龄增长而减弱的现象,研究发现源库比是影响作物叶片衰老的重要因素之一[24]。在作物生长过程中,不利因素往往能够引起叶片早衰,从而导致作物早熟、减产[25]。叶片衰老过程中积累的超氧阴离子能直接引发叶绿素的破坏及特异性地破坏叶绿素a,导致叶绿素分解破坏和叶绿素a/b值下降[26]。叶绿素分解被认为是叶片衰老的原发过程及衰老的真正标志,并且随着叶片衰老、叶绿素含量的下降,叶片光合能力也随之下降[27]。Pn、Gs、Tr、Ci、Chl均能反映叶片的光合能力。Pn是反映光合能力的直接指标,也是综合结果;Chl与光能截获与转化相关;Gs指示叶片获取CO2的水平;Ci则反映叶肉的光合活性,Ci较高说明叶肉将CO2转化为碳水化合物的能力下降。对作物的源、库研究发现,源库比例较低可加速植株的衰老,源库比例过大,叶片合成的光合产物原位积累对光合速率产生反馈抑制,同样会加速叶片衰老[28]。对花生和葡萄的研究均表明,摘除叶片后,光合作用源减少,源器官负荷加重,叶片衰老加快,且源库比越低,叶片衰老越快[1,29]。本研究结果与此一致,叶果比10~40的处理下,叶果比低的处理衰老速率大于叶果比高的处理,主要是由于叶片氮素转出、叶绿素降解,光合产物不足,叶肉细胞能量短缺,根系同化物供应不足等,造成水分、养分吸收能力和细胞分裂素等生长类激素合成能力下降,导致叶片衰老加速。另外,本研究中叶果比50的处理下光合产物在叶片积累导致了反馈抑制,叶面积指数过大导致冠层通风透光变差,引起中下层叶片同化物生产不足而长期处于“饥饿”状态,致使叶片加速衰老。植物光合能力的变化基本是由气孔因素引起的,可分为气孔限制型与非气孔限制型。气孔限制型是指叶片关闭气孔,使CO2通过气孔向叶肉细胞间隙扩散减慢,导致同化位点CO2供应不足、Ci下降,因而对光合能力产生限制,使Pn降低。非气孔限制型是指叶绿素含量及相关酶的活性降低,原初反应及碳同化作用受阻,叶肉细胞活性下降,叶肉利用CO2的能力不足,导致叶片Pn降低,同时叶片Ci增加[30-31]。本研究中随着叶片衰老,叶片Pn降低,叶片Ci升高,表明籽瓜叶片光合能力的降低主要属于非气孔限制型。
4 结 论
本研究发现,随着生育期的演进,籽瓜叶片叶绿素含量、净光合速率、气孔导度、蒸腾速率降低,胞间CO2浓度增加。随着籽瓜叶果比减少,果实生育前期叶片叶绿素含量、净光合速率、气孔导度、蒸腾速率增加,胞间CO2浓度降低。在叶果比10~40的处理下,叶果比越小,叶片衰老速率越快,叶果比50的处理由于叶片数量多、叶面积较大,叶片衰老加快。3个供试籽瓜品种在不同生育期的衰老速率无明显差异。叶果比10处理的果实产量最低,其余叶果比处理果实产量无明显差异,且小种子的果实产量显著小于中等种子和大种子的果实产量。因此,通过对籽瓜叶片进行叶果比处理,调整叶片密度,可以改善叶绿素含量及气体交换效率,延缓叶片衰老,为籽瓜高产栽培提供理论技术支撑。
