乳酸代谢与乳酸穿梭理论发展
2020-01-06谢东山
谢东山
(福建师范大学体育科学学院,福建福州 350117)
1 乳酸研究的经典时代
1.1 “乳酸前时代”到“乳酸时代”
探讨乳酸与运动的关系源于肌肉化学的研究。Von Muralt曾将肌肉化学(muscle chemistry)研究的发展分为4个时期:前乳酸时代(pre-lactic acid era)、乳酸时代(lactic acid era)、磷酸原时代(phosphorylation era)、肌球蛋白时代(myosin era)[1],可见乳酸的研究在人类探寻肌肉秘密的早期占据着重要地位。前乳酸时代始于1808年,由有机化学之父Berzelius首次发现了肌肉中游离乳酸的存在[2],但Berzelius的发现也导致这个领域的研究在刚开始后的近百年就陷入瓶颈,这期间不乏乳酸与糖原、乳酸与肌肉酸化、乳酸与肌肉工作量(the amount of work done)及乳酸与肌肉缺氧的报道[1,3],但方法及技术上的限制导致人们对于乳酸的研究始终困难重重。直到1907年,Fletche和Hopkins[4]的经典研究使得肌肉与乳酸的研究有了标志性的进展。他们不但发现肌肉疲劳时乳酸堆积、富氧环境下乳酸消失的现象,并认为收缩过程中释放的乳酸被用于再合成,而他们的研究方法还可以防止在提取和分析肌肉组织中的乳酸前,让肌肉组织中的乳酸在静息状态下增加,解决了长期困扰前人的问题,乳酸时代就此到来。
1.2 “乳酸时代”与肌肉热力学
1910年,Hill等[5]通过肌肉热力学研究将肌肉活动过程分为两部分:运动初始热和有氧恢复热,并测算出两者基本相等,这一结果支持了Fletche和Hopkins收缩过程中释放的乳酸被用于再合成的观点;但Hill也发现,有氧清除乳酸时产生的热量不足乳酸自身氧化热量的1/7,并隐约意识到恢复期乳酸可能并非仅氧化清除的问题。1920年,Meyerhof[6]对肌肉恢复过程中乳酸代谢和氧消耗、产热和氧消耗之间的关系以及安静期、工作期和恢复期肌肉中糖和乳酸的代谢进行了平行测定,这些测定最终为Hill在研究中提出的问题找到了确定性的答案。Meyerhof的研究结果表明,在恢复期与氧消耗量相对应的乳酸代谢量不超过乳酸消失总量的1/4~1/3,显然大部分乳酸是通过其他方式消失的;产热量与氧耗计算出的热量相比偏低,表明乳酸的燃烧与某个吸热过程相结合,并在此过程中利用了部分热量;当乳酸在肌肉中蓄积时,对应数量的糖原消失了,而当乳酸消失时,肌肉中的糖原又增多,且增多的量接近消失的乳酸总量与氧化量的差。这说明了在肌肉恢复过程中每3~4个乳酸分子中至多只有一个被氧化,而剩余的乳酸则重新合成为最初形成它的肌糖原,糖原就此被明确定义为乳酸的前体。Meyerholf和Hill的研究成果为他们赢得了1922年的诺贝尔生理学或医学奖,为之后氧债学说(O2debt hypotheis)的形成和乳酸循环(Cori circle)的发现奠定了基础。应当注意的是,他们的研究是以离体的青蛙肌肉为对象,忽视了生理条件下肌肉代谢的差异及人和哺乳动物与两栖动物在生理结构和机能上的差异,而人和哺乳动物的骨骼肌相较之下拥有更丰富的毛细血管床和更复杂的线粒体网状结构,其机体的代谢率、心输出量以及肌肉与肝脏的灌注率也更高。
1.3 乳酸与氧债学说
继提出有氧恢复热的理论之后,Hill和Lupton又于1923年正式提出氧债学说(O2debt hypothesis),认为运动后氧债的产生源于乳酸的氧化,并将氧债的快、慢时相分别对应于清除肌肉内和肌肉外乳酸的阶段[7]。对氧债的计算为人体在运动时无氧供能的定量提供了一种方法,然而这种通过计算运动后的氧耗评价运动时的无氧代谢能力被证明是很困难的,因为涉及运动后摄氧基线的建立;此外将离体青蛙肌肉等同于人体作为实验对象仍是Hill研究中的局限。
氧债学说于1933年得到了新的发展,Rodolfo Margaria[47]等在哈佛疲劳实验室通过人跑步的实验发现,运动后血乳酸立刻上升,而处于氧债快时相时血乳酸开始下降。Margaria将氧债快时相称作非乳酸成分(alactacid),慢时相称作乳酸成分,即氧债快时相与乳酸清除无关,而慢时相和乳酸清除有关。然而Ole Bang[48,49]在1934—1936年的研究得到了和Margaria不同的结果。他通过研究不同强度和持续时间的运动发现,运动一段时间后血乳酸达到峰值,即使此时氧耗已经回到运动前的水平。这表明Margaria的研究结果只是偶然产生的,而氧债学说在这之后又经历了许多质疑和挑战。如今氧债学说在生理学,特别是运动生理学中依然被普遍视为指导性准则。
2 乳酸和疲劳
长期以来,人们习惯性地认为缺氧引起乳酸堆积,乳酸堆积导致肌肉疲劳。这种认识的基础正是形成于乳酸研究的经典时代,其形成的过程大致是:1808年,Berzelius发现肌肉中存在游离乳酸,并认为这些游离乳酸的浓度和肌肉活动的程度相关,这是认识形成的源头;之后Fletcher、Hill、Meyerhof等研究人员钻研氧耗和乳酸的关系,氧债学说在他们的一系列研究下形成;20世纪60~70年代初,Wasserman等以氧债学说为理论依据提出无氧阈的概念,即肌肉收缩或运动导致的乳酸生成量激增是由于缺氧引起氧化磷酸化被抑制,至此乳酸和缺氧成为一对双生子;20世纪70~90年代,Sahlin等[8]和Bangsbo等[9]的研究指出乳酸堆积导致质子(H+)堆积进而降低了pH值,乳酸堆积引起肌肉酸化进而导致疲劳的认识成型。乳酸和疲劳的关系至今仍处于争议中,最早反对缺氧引起乳酸论断的是Jöbsis和Stainsby[10],他们使用表面荧光技术对狗股薄肌和腓肠肌中的氧化还原力(NAD/NADH)进行测试,发现肌肉收缩时NADH依然持续氧化成NAD+,电子呼吸链并未被抑制,由此认为乳酸并非缺氧的产物;1984年,Richard Connett[50]等发现收缩的狗骨骼肌中,氧分压在乳酸产生过程中保持在临界氧分压之上,这说明运动时骨骼肌中的乳酸是在氧充足的情况下产生的;1998年,Richardson等[11]对人的四头肌在静息和运动时的乳酸平衡进行测试,并结合NMR技术检测肌红蛋白的饱和度,结果证明运动时虽然肌肉氧分压明显下降至533 Pa,但依然保持在线粒体临界氧分压(1~267 Pa)之上许多,这在2017年Bendahan等[12]的研究中也得到了印证。可以肯定的是,运动时体内pH会下降,但导致pH下降的原因是复杂多样的。Peter Stewart[13]认为pH是水受到PCO2、弱酸浓度[Atot]和强离子差的影响而解离所产生的特性,并通过一系列推导得出强离子差的公式:
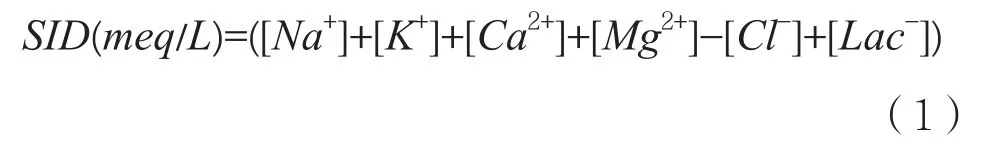
从中可以发现,乳酸并非唯一导致质子浓度变化的因素。事实上,近15年有研究发现乳酸有碱化血液pH的作用[14-15],还可以增强肌肉兴奋性并缓解疲劳[16]。
乳酸研究的经典时代将生理学引向一个新领域并达到新的高度,形成一套成体系的理论,将氧运输和糖代谢通过乳酸代谢有机地联系起来,有氧代谢和无氧代谢供能理论的建立和完善得益于此,从而深远地影响运动人体科学和运动训练学的发展,但也使乳酸逐渐背上代谢终产废物之名。然而科学发展总是在自我纠正的进程中,人们对乳酸的认识也向着更积极的方向发展,细胞间乳酸穿梭理论是改变人们对乳酸态度的转折点。
3 乳酸研究的革命
3.1 细胞间乳酸穿梭
细胞间乳酸穿梭理论的形成大致经历了3个阶段。第一,20世纪70~80年代初,随着同位素示踪法在糖与乳酸代谢研究中的应用[17-22],乳酸穿梭的概念在学界出现;同时,肌肉组织化学和肌肉生物化学特性的研究使人们对肌肉类型的划分和代谢特征有了更清晰的认识[23-24],进一步促进了这一概念的诞生。第二,Stanley等[25]利用同位素示踪法,发现肌肉运动产生的大量乳酸明显被摄取并转化成CO2。第三,基于上述研究,Brooks等[26]发展了乳酸穿梭的假设,并于1985年系统地提出细胞间乳酸穿梭的理论。
细胞间乳酸穿梭理论试图说明乳酸是机体有氧供能和糖异生的重要碳源,乳酸被产生它的细胞本身利用,或被输运到毗邻的细胞、组织和器官中利用。更为具体的理解如图1所示:运动伊始,乳酸在快肌纤维中产生并累积,之后一部分乳酸进入组织间质被毗邻慢肌纤维摄取并氧化,另一部分乳酸被心肌、肝脏和肾脏氧化或作为糖异生的底物。被慢肌纤维摄取的乳酸在氧化时受到两方面因素的影响:第一,如图2所示,脂肪作为能量来源,其氧化分解会抑制糖酵解,从而导致乳酸生成速率减慢;第二,线粒体的活性决定线粒体氧化底物的能力,从而影响丙酮酸和乳酸的清除。细胞间乳酸穿梭理论发表前后,Brooks及其所在团队做了大量关于葡萄糖和乳酸流量(glouse and lactate flux)的实验,验证和推动这一理论的发展,不仅确认了运动及运动后的恢复期有氧氧化是乳酸清除的主要途径(75%),强调了乳酸是有氧供能和糖异生的底物,还肇启了乳酸新的生物学意义,即乳酸代谢调节了氧化还原力进而影响细胞间的信号沟通[25-36]。
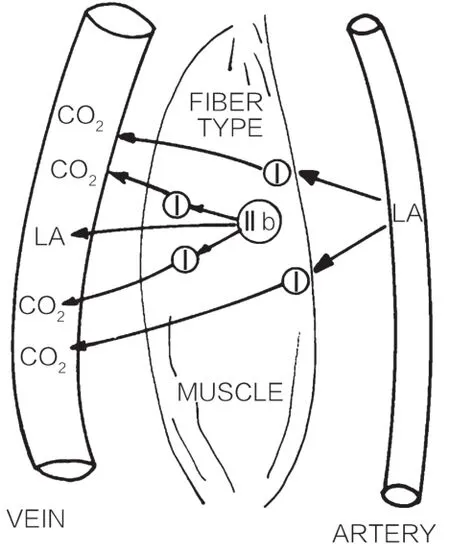
图1 细胞间乳酸穿梭[51]
运动伊始,乳酸在快肌纤维中产生并累积,之后一部分乳酸进入组织间质被毗邻慢肌纤维摄取并氧化,另一部分乳酸通过血液循环被心肌、肝脏和肾脏氧化或作为糖异生的底物
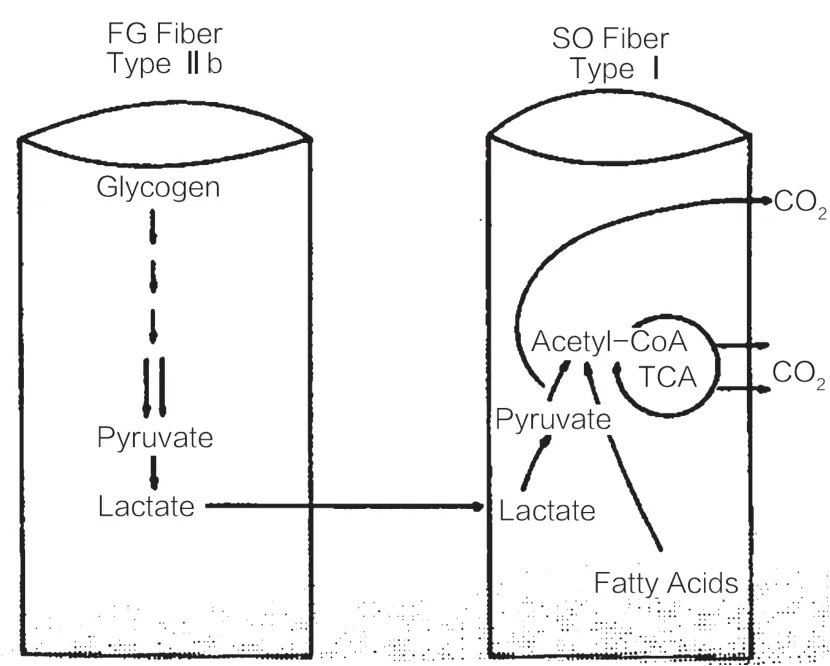
图2 Ⅱb型肌纤维与Ⅰ型肌纤维间乳酸穿梭
如今,细胞间乳酸穿梭理论中的大部分观点已经得到学界的广泛认可,该理论提示人们:第一,乳酸并不是代谢终产废物,而是代谢中间物,是重要的能源底物;第二,无论静息还是运动状态下,机体从未停止过代谢乳酸的步伐,早先人们视乳酸为肌肉组织因缺氧而生的观念也已作古。细胞间乳酸穿梭理论被长期应用于指导运动训练与恢复,运动训练后采用积极恢复方式促进乳酸的氧化代谢已成为常规。而乳酸在细胞信号沟通方面的意义也为“星形胶质细胞-神经元乳酸穿梭”这一研究热点进行了铺路并得到了反哺。
Brooks在细胞间乳酸穿梭理论的基础上,开始探索细胞内乳酸穿梭的奥秘。这一探索引来争议的同时,提出了“乳酸究竟在哪里被氧化”的疑问。
3.2 细胞内乳酸穿梭
3.2.1 细胞内乳酸穿梭的发展
1946年,Cori夫妇[37]认为处理乳酸的主要途径是通过肝脏和肾脏将乳酸作为糖异生的底物并转化为重碳酸盐。之后Mazzeo等[38]和Stanley等[25]经过研究发现,人和其它哺乳动物体内的乳酸大部分都被氧化为CO2,而重碳酸盐的形成只是因为CO2经碳酸酐酶的催化与水产生反应的结果。因为发现肌肉收缩产生的乳酸被大量氧化,人们开始对骨骼肌、心肌以及其它组织和细胞生成的乳酸的氧化场所产生好奇。细胞溶质多被认为是乳酸氧化为丙酮酸的起步场所,然而至今获得的实验数据还不能很好地支持这个观点。Henderson等[39]通过实验发现,人在静息状态下动脉血乳酸和丙酮酸的比值为10~20,但当肌肉进行中等强度的运动收缩时,静脉血中乳酸和丙酮酸的比值成倍上升,可以超过500。这说明即使丙酮酸有所升高,乳酸升高的程度也远远大于丙酮酸。由于红细胞和肺组织中乳酸脱氢酶(LDH)的存在,乳酸和丙酮酸间可逆反应导致比值上升的过程虽然显著,但也得到了一定的缓冲,使得这个剧烈上升过程在理论上变得迟钝,但依然存在极高的比值。细胞溶质作为乳酸生成的直接场所却没能在第一时间将其氧化为丙酮酸,使得研究者将目光投向线粒体,细胞内乳酸穿梭理论由此发轫。
细胞内乳酸穿梭理论由Brooks于1998年提出[28]。Brooks认为细胞溶质内由糖酵解或糖原分解生成的乳酸可以直接进入到该细胞的线粒体中被氧化,而不需要在细胞溶质中氧化为丙酮酸后再进入线粒体。理论以LDH酶动力学的研究为基础,认为线粒体内质网是丙酮酸井(pyruvate trap),可以使丙酮酸浓度极低,从而造成乳酸与丙酮酸间的氧化还原反应因乳酸的浓度优势向逆优势反应方向(乳酸氧化成丙酮酸)发展,产生的丙酮酸则继续在丙酮酸脱氢酶(PDH)催化下维持低浓度[29]。如图3所示,Brooks还提出线粒体膜上氧化复合体(mLOC)的模型来巩固自己的理论。mLOC由单羧酸转运蛋白1(MCT1)及其分子伴侣CD147、环氧化酶(COX)、乳酸脱氢酶(LDH)组成,能够将乳酸氧化为丙酮酸并转运至线粒体基质内。这一理论在学术界存在争议,其争论焦点在于线粒体上是否存在转运乳酸进入线粒体基质的载体以及线粒体中是否存在LDH及其活性如何。
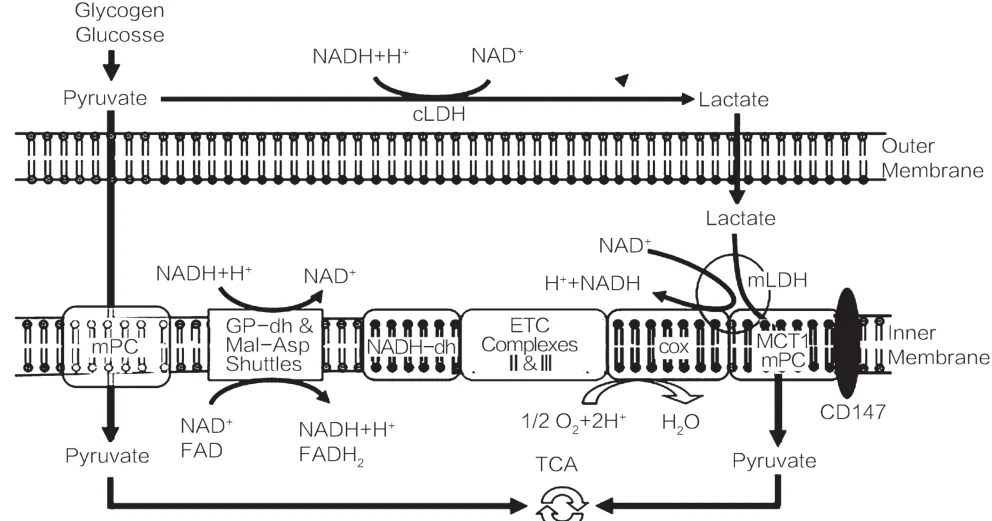
图3 细胞内乳酸穿梭[29]
线粒体膜上氧化复合体(mLOC):由单羧酸转运蛋白1(MCT1)及其分子伴侣CD147、环氧化酶(COX)、乳酸脱氢酶(LDH)组成,能够将乳酸氧化为丙酮酸并转运至线粒体基质内。
3.2.2 细胞内乳酸穿梭理论的争议
Hashimoto等[40]通过总结和归纳,得到了以下6点支持这一理论的证据:第一,通过电子显微镜和激光扫描共聚焦显微镜镜检均发现大鼠及人的骨骼肌、心肌、肝脏等组织细胞的线粒体中存在LDH;第二,显微镜镜检及蛋白质免疫印记实验表明大鼠及人的骨骼肌、心肌等组织细胞的线粒体与MCT1共定位;第三,从大鼠与人的骨骼肌、心肌、肝脏和肾脏等组织细胞分离出的线粒体均包含LDH、MCT1及其分子伴侣CD147;第四,使用大鼠骨骼肌、肝脏细胞进行试验得出的生理数据可知,与氧化丙酮酸相比,线粒体可以更快地氧化乳酸;第五,通过同位素示踪和核磁共振(MRS)方法证明了人和哺乳动物的心肌和骨骼肌细胞的线粒体能够氧化乳酸;第六,线粒体蛋白质组学研究结果支持这一假说。
Sahlin等[41]和Halestrap[42]提出了3点反对这一理论的证据。第一,实验证明,经过精确的密度梯度离心排除了线粒体断片存在MCT1和LDH的可能。第二,由线粒体断片测得的LDH活性极低,仅有0.7%,这与线粒体中NADH/NAD+氧化还原对还原力更强的事实相符;而如果乳酸能够在线粒体内被氧化成丙酮酸,则NADH/NAD+氧化还原对中氧化力应该占主导。所以在热力学定理下,乳酸在线粒体内转化成为丙酮酸的理论是不可行的。第三,细胞质膜和线粒体分别拥有不同的单羧酸转运载体,后者为丙酮酸转运体(MPC),是线粒体载体家族的一员,对乳酸的亲和力极低。
双方争论的终点落在线粒体样品的制备上,即线粒体内膜小囊泡的分离及如何排除线粒体内质网的干扰。线粒体在样品制备过程中因外在污染(细胞质)而使实验得出其拥有氧化乳酸的功能或样品在制备过程中损失了这一功能成为了这一争论中的疑点。
细胞内乳酸穿梭理论具有一定的颠覆性,如果承认这一理论,则需要重新思考如何在运动中及运动后消除乳酸,而考虑到快肌本身氧化乳酸的能力,也应重新审视传统的以乳酸为运动训练监控和评价指标的体系,有氧运动和无氧运动的分类界限也将因此被模糊。笔者对细胞内乳酸穿梭理论持保守态度,原因有以下3点。第一,尽管Brooks的理论以核磁共振、高精度显微镜镜检等先进实验手段的结果为佐证,却还无法合理地解释一个基本问题:丙酮酸井假设的合理性依赖一定浓度的丙酮酸,因为只有丙酮酸达到足够的浓度,才能适用丙酮酸脱氢酶(PDH)的催化反应,并且能够维持足够的NADH来推动电子传递链的反应[43]。促使这一理论体系运转所要求的极低丙酮酸浓度的界限仍需更进一步和更精准的实验来验证。第二,静息状态下细胞内丙酮酸本身浓度极低,远低于乳酸,过高的丙酮酸对人体的危害大于乳酸堆积,生理规律本身可能会限制丙酮酸浓度的升高,运动前后乳酸和丙酮酸比值的剧烈上升似乎并不能成为否认乳酸在细胞溶质中氧化成丙酮酸的有力证据;第三,如图4所示,Halestrap[42]在研究中提到,单羧酸转运蛋白在将乳酸由胞外转运至胞内的过程中,存在限速步骤,即单羧酸转运蛋白在完成乳酸转运的循环恢复构象时,需要更长的时间(k2阶段)。根据细胞间乳酸穿梭理论,乳酸转运至对其氧化能力更强的组织或细胞中时,时间可能会延搁,加上转运时程,不会在短时间内充分地被氧化为丙酮酸。
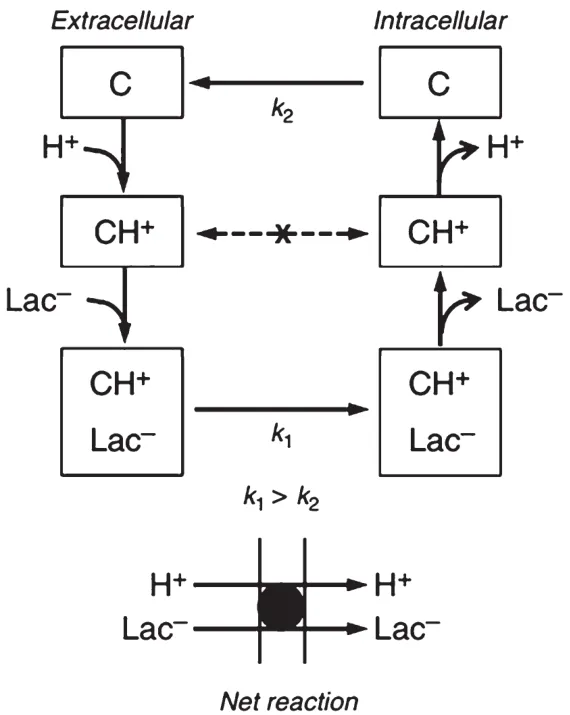
图4 单羧酸转运蛋白动力学机制[42]
单羧酸转运蛋白在将乳酸由胞外转运至胞内的过程中,存在限速步骤,在完成乳酸转运的循环恢复构象时,需要更长的时间(k2阶段),而从结合到分离乳酸根时的构象变化(k1阶段)则更快。
研究者们对于乳酸氧化是否发生在线粒体的争论还在继续,对于乳酸本身及其代谢的认识也在加深。人们应该更加积极的看待乳酸,并重新审视其代谢终产废物的身份,阐明乳酸氧化产所的问题也将更好地理解细胞间乳酸穿梭理论。
3.3 国内乳酸研究进展
由于对学术权威的迷信和思维惯性,国内学术界对于乳酸的认识和研究略显滞后,旧有观念依然普遍指导着体育科学的教学和研究工作。事实上,在1991年,国内已经有学者提出废除氧债和无氧阈、建立运动后过量氧耗和个体乳酸阈的意见[44-45],但影响甚微。近些年,有学者开始关注到乳酸直接参与调节细胞线粒体功能的现象[46],这对体育科学科研工作者来说,是个良好的信号和示范。
4 结 语
乳酸的早期研究建立在生理学、化学、热力学等传统学科上,进行直接观察的条件有限,且实验对象从两栖动物过渡到哺乳动物,造成实验数据并不稳定,因此形成了一些分歧。随着同位素示踪法和核磁技术等直接观察手段的发展,研究者们获得了更精确的数据,并意识到了两栖动物和哺乳动物在生理机能上的差异,这些对修正前人的经典研究都是有利的,同时颠覆了乳酸是代谢废物的身份。如今更进一步的研究探讨乳酸的信号作用,并将其视为伪激素,这也为运动对机体的调控提供了新的思路。
