赤霉素对植物种子萌发及幼苗生长影响的研究进展
2019-12-23杜晨曦王金丽周华坤殷恒霞
杜晨曦 王金丽 周华坤 殷恒霞
摘要:赤霉素(Gibberellins,GAs)作为植物体内广泛存在的一类生长调节剂,参与调控植物种子萌发,下胚轴伸长,叶片伸展,花、果实及种子发育等诸多生理过程。大量研究表明,赤霉素在植物抵抗非生物胁迫中也发挥着重要作用,主要通过调节GAs的生物合成、信号转导及其生物活性,以提高植物对非生物胁迫的耐受性。系统综述了近年来外源赤霉素对植物种子萌发以及幼苗生长发育影响的相关研究,从种子萌发、生理生化特性、相关基因调控等方面进行了系统阐述,旨在为赤霉素在农作物生产中的利用提供参考依据。
关键词:赤霉素;种子萌发;幼苗生理特性;基因调控
中图分类号:Q945 文献标识码:A
文章编号:0439-8114(2019)22-0009-06
DOI:10.14088/j.cnki.issn0439-8114.2019.22.002 开放科学(资源服务)标识码(OSID):
Research progress on the effects of gibberellin on plant
seed germination and seedling growth
DU Chen-xi1a,1b,WANG Jin-li1a,1b,ZHOU Hua-kun2,YIN Heng-xia1a
(1a.State Key Laboratory of Plateau Ecology and Agriculture Qinghai University;1b.College of Eco-Environmental Engineering,Qinghai University,Xining 810016,China;2.Key Laboratory of Restoration Ecology of Cold Area in Qinghai Province,Northwest Institute of Plateau Biology, Chinese Academy of Sciences,Xining 810008,China)
Abstract: Gibberellin (Gibberellins, gas) as a kind of growth regulator widely existing in plants, are involved in the regulation of plant seed germination, hypocotyl extension, leaf extension, the development of the flower, fruit and seed and many other physiological processes. A large number of studies have shown that GAs also play an important role in plant resistance to abiotic stress by regulating the biosynthesis, signal transduction and biological activity of GAs, in order to improve the tolerance of plants to abiotic stress. The recent studies on the effect of exogenous gibberellins on seed germination and the growth and development of seedlings were systematically reviewed, mainly focusing on seed germination, physiological and biochemistry characteristics, and related gene regulation, and an important reference basis for the utilization of the gibberellin in the production of the crops was provided.
Key words: gibberellin; seed germination; seedling physiological characteristics; gene regulation
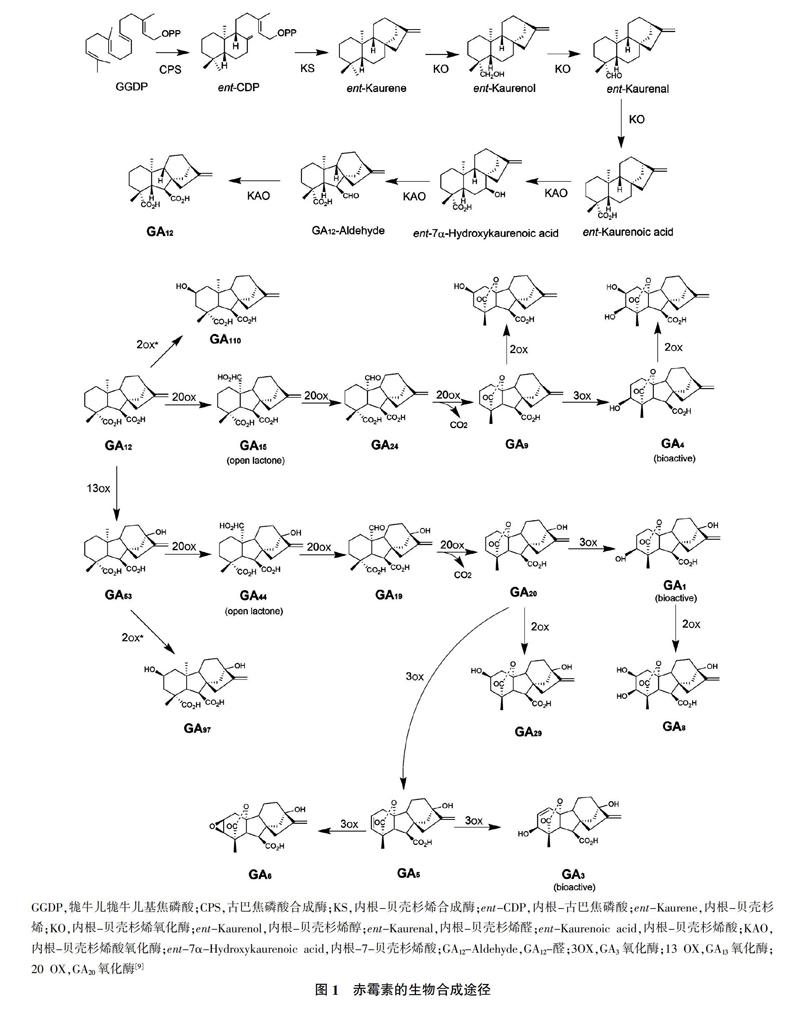
赤霉素(Gibberellin,GAs)是廣泛分布于植物的一类重要生长调节剂,参与种子萌发,幼苗生长发育、果实成熟发育等重要的生理过程[1,2]。赤霉素的种类很多,自1926年日本学者黑泽英一在研究水稻恶苗病中发现赤霉素以来,已陆续发现136种天然赤霉素,但仅有部分赤霉素具有生理功能,如GA1、GA3、GA7、GA30等。赤霉素是由4个异戊二烯组成的二萜类物质,主要在高等植物细胞的质体、内质网和细胞质基质中合成[3],其生物合成途径如图1所示[4],其中参与赤霉素合成的主要酶中,CPS和KS属于TPSs酶,K0和KAO属于P450类,GA20ox和GA3ox等属于2ODDs类[5-8]。
1 外源赤霉素对植物种子萌发的影响
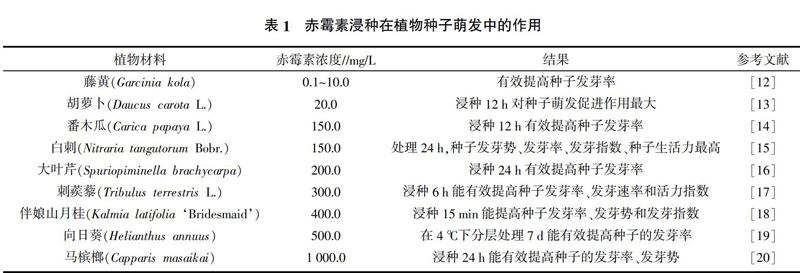
赤霉素能促进生长素的合成和细胞分裂膨大,提高种子胚内酶的活性并促进代谢活动,尤其对休眠种胚的生长有显著的促进作用(表1)[10]。研究发现,赤霉素通过诱导植物相关基因的表达控制酶蛋白的合成和各种内源生长调节剂的分泌,促进植物体内多种生理生化代谢反应,从而提高种子的活力[11]。
不同植物种子对于外源赤霉素的敏感程度也不尽相同,因此,摸索适宜浓度的赤霉素对于种子萌发具有重要意义(表1)。赤霉素促进植物种子萌发的机理主要有两个方面:首先,一定浓度的赤霉素对种皮的蜡质层具有不同程度的腐蚀作用,浸种后能提高种皮的透水、透气性,增强种子内生理生化过程与呼吸作用,促进胚生长,从而能促进种子萌发[21];其次,赤霉素能促进生长素的合成和细胞分裂膨大,提高种子胚内酶(如淀粉酶)的活性和代谢活动,从而提高其发芽势和发芽率[22,23]。另外,研究发现种子浸出液电导率的高低能够反映种子细胞膜的完整性[24],通过对羊草、洋葱种子浸出液的电导率进行检测发现,赤霉素浸种浓度过高时,与对照相比电导率较高,可能高浓度的赤霉素对种子产生了胁迫,反而抑制了种子的萌发[25,26]。
2 外源赤霉素对植物幼苗生理生化特性的影响
赤霉素通过促进细胞分裂伸长进而调控植物株高、器官大小、叶面积以及根系发育[27-32]。江雪[27]研究发现,外源赤霉素显著促进了毛竹实生苗新分蘖植株的株高和节间伸长;李品荣等[28]研究发现,外源赤霉素能促进李子开花结实。大量的研究表明,外源赤霉素对株高、单株叶片数、嫩枝鲜重等都具有十分重要的作用[29-32]。
2.1 外源赤霉素对植物幼苗抗氧化系统酶活性的影响
自然环境条件下,植物为了维持正常的生长发育,通过自身抗氧化酶系统和抗氧化剂(即非酶系统) 进行不利生长条件下活性氧的清除[33]。其抗氧化酶系统中超氧化物歧化酶(SOD)是植物抗氧化的第一道防线,能清除细胞中多余的超氧阴离子;过氧化氢酶(CAT)和过氧化物酶(POD)可以使H2O2歧化成无毒害的水和氧分子[34]。丙二醛(MDA)是植物器官衰老或在逆境条件下受伤害时组织或器官膜脂质发生过氧化反应而产生的,它的含量与植物衰老及逆境伤害有密切关系。研究发现,外源赤霉素通过提高活性氧清除剂SOD和CAT的活性调节脂质过氧化,从而抑制植株衰老[35-38]。同时,研究表明,外源喷施赤霉素后,植株体内MDA含量显著降低,有效地防止了保护酶和膜系统的破坏,延缓了膜系统的衰老[13,37,39]。
2.2 外源赤霉素对植物幼苗渗透调节物质含量的影响
可溶性蛋白质、可溶性糖和脯氨酸是植物体内主要的渗透调节物质,通过保持植物细胞的含水量和膨压势来维持正常的生长和代谢[40]。Matsukura等[41]研究发现,外源赤霉素能够增加水稻植株中包括糖在内的渗透溶质的总量。邹竣竹[42]、刘春冬等[43]研究发现,低浓度的赤霉素促进了野牛草和杨梅幼苗中可溶性糖、可溶性蛋白含量的增加。何丽等[39]对番茄的研究发现,外源赤霉素处理的番茄幼苗体内脯氨酸的含量显著低于对照。因此,外源赤霉素处理能够促进植物体内渗透调节物质的显著增加,并使植物细胞保持较高的含水量,从而保证了植物的正常生长和代谢活动。
2.3 外源赤霉素对植物幼苗叶片叶绿素含量的影响
Aftab等[44]研究发现,外源赤霉素处理使得黄花蒿叶片的气孔导度、内部CO2含量和净光合速率显著提高。同时,大量研究表明,外源赤霉素处理使叶片的叶绿素含量、细胞间CO2浓度、光合速率、气孔导度和蒸腾速率都有不同程度的增加,延缓衰老过程中叶绿素的降解[45-48]。
2.4 外源赤霉素对植物内源生长调节剂含量的影响
内源生长调节剂作为植物体内重要的调控因子,参与了植物生长发育过程中一系列的生理生化过程,对植物营养生长时期各器官的形成和生长发挥着重要作用[49]。内源生长素可以通过调节赤霉素的合成和代谢,共同调控植物主根、侧根和根毛的形成延伸,叶片中营养元素的运输以及促进叶原基的发生等生理活动。内源脱落酸、生长素和赤霉素在植物生长发育的多个过程中存在相互作用,共同调控植物的生长发育过程。研究表明,植物通过调节生长素和脱落酸的平衡来控制侧根的发育,在幼苗生长时期脱落酸与赤霉素具有相互拮抗作用[50],而且外源赤霉素能够降低内源脱落酸水平[51]。通过外源赤霉素浸种后植株内赤霉素和生长素含量均呈逐渐升高的趋势,反之,脱落酸和乙烯的含量呈逐渐下降的趋势,由此可以看出,外源赤霉素通过降低脱落酸含量来打破植物种子休眠而促进种子萌发[52-56]。Liu等[57]研究发现,外源赤霉素提高了水稻分蘖节中的ABA和IAA含量,而降低了玉米素+玉米素核糖苷(Z+ZR)的含量。總而言之,外源赤霉素能够使植株内IAA含量升高,而IAA能够直接调节细胞的伸长,从而促进了植物的生长发育。
3 外源赤霉素对植物幼苗生长相关基因表达的影响
赤霉素信号传导路径中的主要元件由活性GA受体GID1蛋白、核心调控DELLA蛋白、泛素E3连接酶复合体(SCFSLY1/GID2)和降解作用的26S蛋白酶等组成[58-61]。近年来,随着研究的不断深入,赤霉素信号传导路径中主要元件之间的相互作用及功能已比较明确,赤霉素信号传导路径主要过程为活性GA、GID1蛋白受体和DELLA蛋白结合后形成三联体,并且在F-box型蛋白SLY1和GID2的标靶下与26S蛋白相结合后被降解[61-63]。相关研究结果表明,在植物体内内源活性赤霉素水平较低的情况下,DELLA蛋白通过抑制赤霉素效应来减少植物生长,而此过程中调控植物生长的转录因子,如SPY和EL1均可增强其对植物生长的抑制作用[64];而当植物体内内源活性赤霉素水平较高时,GID1蛋白受体通过构象变化结合活性赤霉素,之后与DELLA蛋白形成三联体,最后DELLA蛋白被降解,解除了对植物生长的抑制作用,开启活性赤霉素对植物生长的促进作用[65,66]。
Rinne等[67]研究发现,外源赤霉素处理后使与其合成相关的基因GA20ox和GA3ox的表达量下调。李军等[68]以一定浓度的赤霉素水溶液灌注太子参根部,结果发现赤霉素生物合成关键酶基因GA20ox1、GA20ox2和GA3ox1表达量显著降低,并且2个赤霉素受体蛋白基因GID1a和GID1b的表达量也显著降低。同时,Zhuang等[69]用赤霉素处理果梅枝条后发现,GA20ox、GA2ox等与赤霉素代谢相关的基因也发生了显著差异表达。Ueno等[70]通过转录组测序,发现有2个与赤霉素信号相关的基因GASA和ERD14,以及赤霉素的受体GID1在橡树季节性休眠诱导和解除中起关键作用。Barros等[71]研究扁桃花芽萌发的机制时发现,与赤霉素代谢相关的基因PdGA20ox和PdGA2ox差异表达导致赤霉素代谢发生变化,从而影响扁桃的花芽萌发。Gocal等[72]研究模式植物拟南芥时发现,具有GAMYB样活性的atmy33基因在外源赤霉素处理后表达量增加,且促进植株在短时间内开花。魏志刚等[73]用不同浓度等体积赤霉素(GA3)溶液进行叶面喷洒处理白桦,研究发现与白桦成花相关的基因LFY、ELF3、GI和LD表达量都明显降低,表明外源赤霉素抑制了促进开花相关基因的表达。
4 展望
赤霉素在植物的生长发育过程中发挥着重要的调控作用。目前,虽然已经对赤霉素的作用机制有了初步了解,但在农业生产中为了更好地利用赤霉素,仍需继续进行深入研究。①外源赤霉素与内源赤霉素之间的相互作用机制。研究表明,通过外源赤霉素浸种或者叶片喷施,会使内源赤霉素的含量增加;但是目前外源赤霉素如何调控相应的基因,进而调控内源赤霉素的整个生物合成过程及随后的信号感知及信号传导过程仍不清楚。②赤霉素与其他生长调节剂在休眠进程中的相互作用机理。许多研究表明,赤霉素与脱落酸的含量比值决定着木本植物的休眠状态;同时其他生长调节剂在休眠进程中同样起着重要的作用;研究赤霉素与其他生长调节剂对休眠进程的调控机制,对深入理解赤霉素的破眠机制具有十分重要的意义。③赤霉素代谢相关基因与其他基因之间的调控网络。尽管赤霉素生物合成、信号感知及传导过程已经研究得比较透彻,但赤霉素如何通过调控赤霉素代谢相关基因进而调控其他生命活动相关基因仍不清楚。④赤霉素使用方法的改进。各种传统栽培措施基本上是侧重运用外部条件来影响植物生理状况,而赤霉素的使用,则是外部条件加内源生长调节剂水平的双重调控,从而为农作物栽培获得更高的产量提供了可能。
参考文献:
[1] OGAWA M,HANADA A,YAMAUCHI Y,et al. Gibberellin biosynthesis and response during Arabidopsis seed germination[J].Plant cell,2003,15(7):1591-1604.
[2] CHENG H,QIN L,LEE S,et al. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function[J].Development,2004,131(5):1055-1064.
[3] 潘瑞熾,王小菁,李娘辉.植物生理学[M].北京:高等教育出版社,2012.
[4] YAMAGUCHI S. Gibberellin metabolism and its regulation[J].Annu Rev Plant Biol,2008,59:225-251.
[5] HELLIWELL C A,CHANDLER P M,POOLE A,et al. The CYP88A cytochrome P450,ent-kaurenoic acid oxidase,catalyzes three steps of the gibberellin biosynthesis pathway[J].Proceedings of the national academy of sciences,2001,98(4):2065-2070.
[6] HELLIWELL C A,SULLIVAN J A,MOULD R M,et al. A plastid envelope location of Arabidopsis ent-kaurene oxidase links the plastid and endoplasmic reticulum steps of the gibberellin biosynthesis pathway[J].The plant journal,2001,28(2):201-208.
[7] SUN T P,KAMIYA Y. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase a of gibberellin biosynthesis[J].The plant cell,1994,6(10):1509-1518.
[8] APPLEFORD N E J,EVANS D J,LENTON J R,et al. Function and transcript analysis of gibberellin-biosynthetic enzymes in wheat[J].Planta,2006,223(3):568-582.
[9] 蔡斌华,庄维兵,高志红,等.赤霉素解除木本植物季节性休眠机制的研究进展[J].西北植物学报,2014,34(10):2145-2152.
[10] 李保珠,赵 翔,安国勇.赤霉素的研究进展[J].中国农学通报,2011,27(1):1-5.
[11] 李冬玲,段红霞,刘鸿晨,等.赤霉素及其功能类似物与赤霉素受体的研究进展[J].农药学学报,2013,15(6):601-608.
[12] KOUAKOU K L,KOUAKOU C,KOFFI K K,et al. Effect of mechanical scarification and gibberellins (GA3) on seed germination and growth of Garcinia kola (Heckel)[J].Journal of applied biosciences,2016,103(1):9811-9818.
[13] 张会灵,余义和,郭大龙,等.赤霉素对萝卜种子萌发及幼苗生理特性的影响[J].北方园艺,2016(15):31-33.
[14] DESAI A,PANCHAL B,TRIVEDI A,et al. Studies on seed germination and seedling growth of papaya (Carica papaya L.) CV. madhubindu as influenced by media,GA3 and cow urine under net house condition[J].Journal of pharmacognosy and phytochemistry,2017,6(4):1448-1451.
[15] 郭曄红,蔺海明.赤霉素和细胞激动素对白刺种子萌发的调控研究[J].中国生态农业学报,2009,17(6):1196-1199.
[16] 王永红,苏 李,张艳红.层积处理及赤霉素浸种对大叶芹种子萌发的影响[J].湖北农业科学,2018,57(1):71-73.
[17] 韦 妍,李 萍,田 婵,等.不同浓度赤霉素浸种对刺蒺藜种子萌发特性的影响[J].延安大学学报(自然科学版),2018,37(1):84-87,91.
[18] 付远洪,钱沉鱼,李朝婵,等.不同浓度赤霉素对伴娘山月桂种子萌发的影响[J].种子,2017,36(2):5-8.
[19] ADAMS R P,TEBEEST A K. The effects of gibberellic acid (GA3),ethrel, seed soaking and pre-treatment storage temperatures on seed germination of Helianthus annuus and H. petiolaris[J].Phytologia,2016,98:213-218.
[20] 赵东兴,李 春,李 涛,等.4种植物生长调节剂对马槟榔种子萌发和幼苗生长的影响[J].南方农业学报,2015,46(10):1834-1838.
[21] 申惠翡,赵 冰,黄文梅.温度和赤霉素对金背杜鹃种子萌发和幼苗生长的影响[J].江苏农业科学,2016,44(6):281-283.
[22] INADA S,SHIMMEN T. Regulation of elongation growth by gibberellin in root segments of Lemna minor[J].Plant and cell physiology,2000,41(8):932-939.
[23] UBEDA-TOM?魣S S,FEDERICI F,CASIMIRO I,et al. Gibberellin signaling in the endodermis controls Arabidopsis root meristem size[J].Current biology,2009,19(14):1194-1199.
[24] 钟希琼,王惠珍.高等植物赤霉素生物合成及其调节研究进展[J].植物学通报,2001,18(3):303-307.
[25] 崔 爽.外源赤霉素(GA3)对羊草生长发育及生物量的影响[D].长春:东北师范大学,2004.
[26] ALI M A,HOSSAIN M M,ZAKARIA M,et al. Effect of GA3 on quality seed production of onion in bangladesh[J].Ecofriendly agricultural journal,2015,8(3):47-50.
[27] 江 雪.外源赤霉素对毛竹实生苗生长的影响[D].南京:南京林业大学,2015.
[28] 李品荣,陈 强,常恩福,等.李子丰产栽培技术研究[J].中国生态农业学报,2003(1):130-132.
[29] KUCHLAN P,KUCHLAN M K,HUSAIN S M. Effect of foliar application of growth activator,promoter and antioxidant on seed quality of soybean[J].Legume Res,2017,40(2):313-318.
[30] DILIP W S,SINGH D,MOHARANA D,et al. Influence of gibberellic acid(GA3) on seed germination and seedling growth of Kagzi Lime[J].Journal of scientific agriculture,2018,1:62-68.
[31] 徐小玉,张凤银,曹 阳.赤霉素和乙烯利对美女樱种子萌发及幼苗生长的影响[J].种子,2014,33(6):72-74.
[32] RAHEMI M,BANINASAB A. Effect of gibberellic acid on seedling growth in two wild species of pistachio[J].The Journal of horticultural science and biotechnology,2000,75(3):336-339.
[33] FOYER C H,DESCOURVIERES P,KUNERT K J.Protection against oxygen radicals:An important defense mechanism studied in transgenic plants[J].Plant, cell & environment,1994,17(5):507-523.
[34] 李 璇,岳 红,王 升,等.影响植物抗氧化酶活性的因素及其研究热点和现状[J].中国中药杂志,2013,38(7):973-978.
[35] DHINDSA R S,PLUMB-DHINDSA P L,REID D M. Leaf senescence and lipid peroxidation: Effects of some phytohormones,and scavengers of free radicals and singlet oxygen[J].Physiologia plantarum,1982,56(4):453-457.
[36] 施 园.外源赤霉素对甜瓜幼苗建成与产量形成的影响[D].黑龙江大庆:黑龙江八一农垦大学,2017.
[37] 张秀丽.赤霉素和矮壮素对绿豆生育性状和生理指标及产量的影响研究[D].长春:吉林农业大学,2007.
[38] 赵 鑫,张继澍,王 敏.CaCl2和GA3处理对枣果采后衰老和膜脂过氧化的影响[J].西北农林科技大学学报(自然科学版),2003(2):118-120,124.
[39] 何 丽,刘 杰,何彩燕,等.不同浓度赤霉素对番茄种子萌发及幼苗生长的影响[J].农业科技与信息,2018(4):43-45.
[40] DAI H P, WEI S H,TWARDOWSKA I,et al.Hyperaccumulating potential of Bidens pilosa L. for Cd and elucidation of its translocation behaviorbased on cell membrane permeability[J].Environ Sci Pollut Res,2017,24:23161-23167.
[41] MATSUKURA C,ITOH S,NEMOTO K,et al. Promotion of leaf sheath growth by gibberellic acid in a dwarf mutant of rice[J].Planta,1998,205(2):145-152.
[42] 邹竣竹.激素浸种对野牛草种子发芽与幼苗生长生理的影响[D].北京:中国林业科学研究院,2017.
[43] 刘春冬,傅建卿,陈汝顶.赤霉素对东魁杨梅花芽分化期生理特性的影响[J].南方园艺,2013,24(1):13-14,18.
[44] AFTAB T,KHAN M M A,IDREES M,et al. Stimulation of crop productivity, photosynthesis and artemisinin production in Artemisia annua L. by triacontanol and gibberellic acid application[J].Journal of plant interactions,2010,5(4):273-281.
[45] RAMTEKE V,PAITHANKAR D H,BAGHEL M M,et al. Impact of GA3 and propagation media on growth rate and leaf chlorophyll content of papaya seedlings[J].Research journal of agricultural sciences,2016,7(1):169-171.
[46] 曹柳青.赤霉素对冬枣光合作用和内源激素的影响[D].河北保定:河北农业大学,2006.
[47] SKUTNIK E,RABIZA J,WACHOWICZ M,et al. Senescence of cut leaves of Zantedeschia aethiopica and Z. elliottiana. part I. chlorophyll degradation[J].Acta Sci Pol,2004,3(2):57-65.
[48] KHADER S,SINGH B P,KHAN S A. Effect of GA3 as a post-harvest treatment of mango fruit on ripening,amylase and peroxidase activity and quality during storage[J].Scientia horticulturae,1988,36(3-4):261-266.
[49] RUBIO V,BUSTOS R,IRIGOYEN M L,et al. Plant hormones and nutrient signaling[J].Plant molecular biology,2009,69(4):361-373.
[50] BRADFORD K J,NONOGAKI H. Seed development,dormancy and germination[M].Oxford:Blackwell publishing,2007.
[51] TOYOMASU T,YAMANE H,MUROFUSHI N,et al. Effects of exogenously applied gibberellin and red light on the endogenous levels of abscisic acid in photoblastic lettuce seeds[J].Plant and cell physiology,1994,35(1):127-129.
[52] 方志荣,苏智先,胡进耀. 脱落酸、赤霉素和乙烯对种子休眠的萌发和调控[J].西華师范大学学报(自然科学版),2007,28(2):127-132.
[53] 王 宁,梅海军,袁美丽,等.赤霉素浸种和变温层积过程对冬青种子激素含量的影响[J].河南农业大学学报,2010,44(5):524-527.
[54] 吴建明,李杨瑞,杨 柳,等.赤霉素诱导甘蔗节间伸长与内源激素变化的关系[J].热带作物学报,2009,30(10):1452-1457.
[55] 曹柳青.外源赤霉素对冬枣叶片内源生长调节剂含量的影响[J].南方农业,2015,9(21):193-194.
[56] 郑顺林,袁继超,马 均,等.外源赤霉素对小整薯休眠解除中的激素变化及物质转化的影响[J].西南农业学报,2008,21(2):323-327.
[57] LIU Y,DING Y F,WANG Q S,et al. Effect of plant growth regulators on growth of rice tiller bud and changes of endogenous[J].Acta agronomica sinica,2011,37(4):670-676.
[58] UEGUCHI-TANAKA M,ASHIKARI M,NAKAJIMA M,et al. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin[J].Nature,2005,437(7059):693-698.
[59] PENG J,CAROL P,RICHARDS D E,et al. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses[J].Genes & development,1997,11(23): 3194-3205.
[60] MCGINNIS K M,THOMAS S G,SOULE J D,et al. The Arabidopsis SLEEPY1 gene encodes a putative F-box subunit of an SCF E3 ubiquitin ligase[J].Plant cell,2003,15(5):1120-1130.
[61] MILLER J,GORDON C. The regulation of proteasome degradation by multi-ubiquitin chain binding proteins[J].FEBS letters,2005,579(15):3224-3230.
[62] HARBERD N P,BELFIELD E,YASUMURA Y.The angiosperm gibberellin-GID1-DELLA growth regulatory mechanism:How an “inhibitor of an inhibitor” enables flexible response to fluctuating environments[J].Plant cell,2009,21(5):1328-1339.
[63] SUN T P. The molecular mechanism and evolution of the GA-GID1-DELLA signaling module in plants[J].Current biology,2011,21(9):338-345.
[64] SUN T. Gibberellin-GID1-DELLA:A pivotal regulatory module for plant growth and development[J].Plant physiology,2010, 154(2):567-570.
[65] DILL A,THOMAS S G,HU J,et al. The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation[J].Plant cell,2004,16(6):1392-1405.
[66] GRIFFITHS J,MURASE K,RIEU I,et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis[J].Plant cell,2006,18(12):3399-3414.
[67] RINNE P L H,WELLING A,VAHALA J,et al. Chilling of dormant buds hyperinduces FLOWERING LOCUS T and recruits GA-inducible 1,3-β-glucanases to reopen signal conduits and release dormancy in Populus[J].Plant cell,2011,23(1):130-146.
[68] 李 軍,周 涛,郑 伟,等.外源GA3对太子参块根发育及赤霉素生物合成的调控[J].分子植物育种,2018,16(20):6867-6874.
[69] ZHUANG W,GAO Z,WANG L,et al. Comparative proteomic and transcriptomic approaches to address the active role of GA4 in Japanese apricot flower bud dormancy release[J].Journal of experimental botany,2013,64(16):4953-4966.
[70] UENO S,KLOPP C,LEPL?魪 J C,et al. Transcriptional profiling of bud dormancy induction and release in oak by next-generation sequencing[J].BMC genomics,2013,14(1):236.
[71] BARROS P M,GON?覶ALVES N,SAIBO N J M, et al. Cold acclimation and floral development in almond bud break:Insights into the regulatory pathways[J].Journal of experimental botany,2012,63(12):4585-4596.
[72] GOCAL G F W,SHELDON C C,GUBLER F,et al. GAMYB-like genes,flowering,and gibberellin signaling in Arabidopsis[J].Plant physiology,2001,127(4):1682-1693.
[73] 魏志刚,钱婷婷,张凯旋,等.外源GA3对白桦成花基因影响的定量PCR分析[J].林业科学,2011,47(7):187-192.
