小麦几丁质酶基因家族的全基因组鉴定及禾谷镰刀菌胁迫下的表达分析
2019-11-21刘建丰袁祖丽
徐 武,刘建丰,张 戈,徐 文,袁祖丽
(1.河南农业大学 生命科学学院,河南 郑州 450002; 2.河南省农业科学院 农业经济与信息研究所,河南 郑州 450002; 3.河南省农业科学院 作物设计中心,河南 郑州 450002)
几丁质酶(EC 3.2.1.14)是广泛存在于植物中的一种糖苷水解酶,能水解N-乙酰氨基葡萄糖聚合物[1]。几丁质酶是一类与疾病相关的蛋白质,可以破坏病原菌的细胞壁和昆虫的外骨骼,在植物防御生物和非生物胁迫过程中发挥着重要作用[2]。根据氨基酸序列相似性,可将植物几丁质酶分为糖苷水解酶亚家族18(GH-18)和糖苷水解酶亚家族19(GH-19)[3]。根据CAZy数据库(http://www.cazy.org/Glycoside-Hydrolases.html),植物几丁质酶被分为5个不同的类(Ⅰ—Ⅴ)[4]。其中,Ⅰ类、Ⅱ类和Ⅳ类属于GH-19亚家族,Ⅲ类和Ⅴ类属于GH-18亚家族[3]。Ⅰ类几丁质酶成员具有N-末端甲壳素结合区和GH-19催化结构域;Ⅱ类几丁质酶成员只有1个催化结构域,与Ⅰ类几丁质酶成员具有较高的序列和结构相似性,但缺乏甲壳素结合区;Ⅳ类几丁质酶成员与Ⅰ类几丁质酶成员具有较高的同源性,但由于甲壳素结合区和催化结构域的缺失,其同源性较小[5];Ⅲ类和Ⅴ类几丁质酶成员均具有GH-18催化结构域和保守序列DXDXE,但与其他类成员无氨基酸序列同源性[6]。
在高等植物生长发育过程中,几丁质酶基因在植物抗病过程中起到重要作用[7-16]。1998年,TABEI等[7]通过农杆菌介导的方法将水稻的几丁质酶基因RCC2导入黄瓜,与非转基因植株相比,转基因黄瓜对灰霉病的抗性显著提高。2000年,YAMAMOTO等[8]将1个水稻Ⅰ类几丁质酶基因导入葡萄,显著提高了转基因葡萄对白粉病的抗性。近年来,已将几丁质酶基因转入水稻[9-10]、棉花[11]、大豆[12-13]、花生[14]、烟草[15-16]等高等植物中,也明显提高了转基因植株对各类病菌的抗性,这些研究说明几丁质酶基因在植物抗病应答过程中具有重要的作用。
小麦作为世界上重要的粮食作物之一,其生长发育过程中受到许多病菌的危害,尤其是禾谷镰刀菌侵染引起的赤霉病。近年来,赤霉病成为造成小麦严重减产的病害之一,不仅降低小麦产量,而且还产生赤霉毒素影响品质,甚至危害人类健康。自小麦参考基因组序列公布以来,小麦基因功能的研究报道越来越多,但是小麦中几丁质酶家族成员全基因组鉴定的研究尚未见报道。研究几丁质酶基因对提高小麦赤霉病抗性具有重要意义。为此,对小麦几丁质酶基因家族成员进行鉴定,并对其基因结构、保守基序(Motif)、染色体分布、系统进化及禾谷镰刀菌胁迫下的表达情况进行分析,为进一步研究几丁质酶基因在小麦生物胁迫应答方面的作用及小麦抗病分子育种中的应用奠定基础。
1 材料和方法
1.1 数据来源
小麦基因组参考序列及其注释文件均下载自Ensembl Plants数据库(ftp://ftp.ensemblgenomes.org/pub/plants/release-41)。
1.2 小麦几丁质酶基因家族成员的鉴定
小麦几丁质酶基因家族成员结构域信息隐马尔科夫模型(PF00704和PF00182)均下载自Pfam数据库(http://pfam.xfam.org/)。利用HMMER 3.1软件结合几丁质酶基因家族成员结构信息隐马尔科夫模型搜索小麦基因组注释的蛋白质序列文件,E值设置为 1E-10。如果同一个基因对应的多个注释蛋白质序列被鉴定到,选取长度最长的蛋白质序列。采用Compute pI/Mw tool(https://web.expasy.org/compute_pi/)预测蛋白质的等电点和分子质量。
1.3 小麦几丁质酶基因家族基因结构和保守基序分析
根据鉴定到的小麦几丁质酶家族成员蛋白质序列编号和Ensembl Plants中小麦基因组注释信息文件,提取小麦几丁质酶基因家族基因的注释信息,然后将其提交GSDS网站(http://gsds.cbi.pku.edu.cn)来绘制基因结构图。用MEME软件(http://meme-suite.org/tools/meme)来预测几丁质酶家族成员的保守基序。
1.4 小麦几丁质酶家族成员的系统进化分析
将鉴定到的小麦几丁质酶家族成员蛋白质序列利用MEGA 7.0软件包中的MUSCLE软件进行多重序列比对,然后进行系统进化树构建。系统进化树采用NJ(Neighbor-joining)法构建,抽样次数为1 000[17]。
1.5 小麦几丁质酶基因家族基因的表达分析
小麦RNA-Seq测序数据来自SRA数据库[18],登录号为SRR7000820—SRR7000822、SRR7000826—SRR7000828、SRR7000832—SRR7000834、SRR7000838—SRR7000840、SRR7000844—SRR7000846、SRR7000850—SRR7000852、SRR7000856—SRR7000858、SRR7000862—SRR7000864。采用TopHat 2[19]软件将其与参考基因组序列比对,运用HTSeq软件[20]统计每个转录本的reads数,然后用DESeq2 R包[21]进行差异分析,保留log2(变化倍数)大于1的转录本进行热图制作。
2 结果与分析
2.1 小麦几丁质酶基因家族成员的鉴定及序列特征分析
在小麦基因组中共鉴定到159个几丁质酶基因家族成员,如表1所示,这些几丁质酶基因主要分成2个亚家族,即GH-18和GH-19家族,分别有82个和77个基因。小麦几丁质酶基因编码蛋白质序列长度为157~739个氨基酸,分子质量为16 648.96~79 774.66 u,理论等电点介于4.10~10.25。对小麦几丁质酶基因家族成员进行染色体定位分析发现,本研究鉴定到的159个小麦几丁质酶基因家族中的154个基因不均匀分布在21条染色体上,5个定位在尚未组装完整的scaffold上。3号染色体上几丁质酶基因数目最多,共49个,其中3A、3B、3D染色体上分别有18、16、15个;2号染色体次之,含有31个,2A、2B、2D染色体上分别有12、8、11个;7A、7B、7D染色体上分别有8、12、7个; 1A、1B、1D染色体上分别有9、8、6个;5A、5B、5D染色体上分别有7、4、4个;4A、4B、4D染色体上分别有1、1、3个;6号染色体上数目最少,共4个,6A、6B、6D染色体上分别有2、1、1个。从染色体组的角度来看,同一编号染色体A、B、D 3套染色体组含有的几丁质酶基因数目差别不大,表明几丁质酶基因在A、B、D 3套染色体组进化过程中没有发生明显的基因丢失现象。

表1 小麦几丁质酶基因家族成员信息
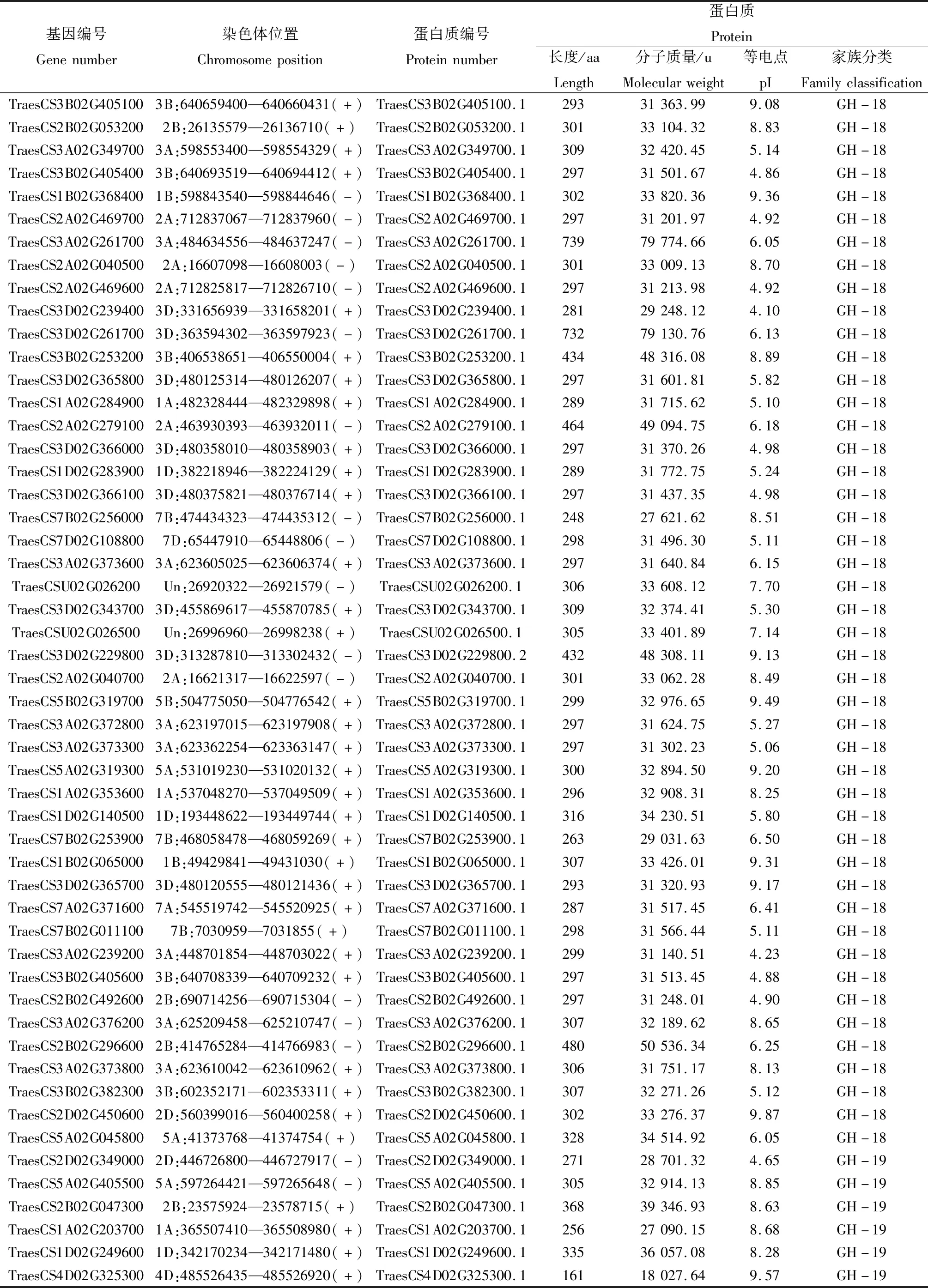
续表1 小麦几丁质酶基因家族成员信息

续表1 小麦几丁质酶基因家族成员信息

续表1 小麦几丁质酶基因家族成员信息
2.2 小麦几丁质酶家族成员的分类
利用本研究鉴定到的小麦几丁质酶家族成员蛋白质序列构建系统进化树,并与已知拟南芥几丁质酶家族成员蛋白质序列进行比对,然后根据已知拟南芥几丁质酶家族成员分类信息对小麦几丁质酶家族基因成员进行分类,结果(图1)表明,小麦几丁质酶家族成员分为5类,GH-19和GH-18亚家族分别聚在2个不同的分支。GH-18亚家族包括Ⅲ类和Ⅴ类,其中,Ⅱ类含有34个成员,Ⅴ类含有48个成员。GH-19亚家族中包括Ⅰ、Ⅱ、Ⅳ类,其中,Ⅰ类含有52个成员,Ⅱ类含有13个成员,Ⅳ类含有12个成员。

图1 小麦几丁质酶家族成员的系统进化树Fig.1 The phylogenetic tree of chitinase family members in wheat
2.3 小麦几丁质酶基因家族成员的基因结构和保守基序分析
为了进一步研究小麦几丁质酶基因家族成员的进化关系,根据小麦基因组参考序列的功能注释信息并绘制其外显子-内含子结构图(图2—3),结果表明,GH-18亚家族基因含有1~8个外显子,其中,大部分只有1个外显子,不含有内含子;少量含有1~2个内含子;TraesCS3D02G229800.2、TraesCS3A02G223700.1和TraesCS3B02G253200.1三个基因含有7~8个内含子。GH-19亚家族基因含有1~5个外显子,大部分成员含有1~2个内含子,基因长度差别不大,其中,TraesCS3A02G415700.1基因内含子长度比较长;TraesCS3D02G229800.2、TraesCS3A02G223700.1、TraesCS3B02G253200 .1和TraesCS3A02G415700.1四个基因长度比其他几丁质酶基因长很多,而且内含子长度比外显子长度长,说明在进化过程中这4个基因内部可能发生了大片段的插入。对GH-18和GH-19亚家族成员中的保守基序进行分析(图4—5)发现,GH-18亚家族82个成员中有79个包含保守基序5,67个成员中包含保守基序1、保守基序2、保守基序3、保守基序4;GH-19亚家族77个成员中有74个包含保守基序1、保守基序4,其中,GH-19亚家族的第Ⅳ亚类不含有保守基序2,剩下65个成员中62个含有此基序。综上所述,相同亚家族的几丁质酶基因几乎具有相似的外显子-内含子结构和保守基序。

图2 小麦GH-18亚家族成员基因结构分析

图3 小麦GH-19亚家族成员基因结构分析

图4 小麦GH-18亚家族成员保守基序分析
2.4 小麦几丁质酶基因家族成员的表达分析
通过分析4个小麦品系的RNA-Seq数据,得到几丁质酶基因家族成员在小麦接种禾谷镰刀菌前后的表达谱。由图6可知,虽然几丁质酶基因在不同小麦品系中被诱导表达的程度存在差异,但是在4个小麦品系被禾谷镰刀菌侵染后,一些基因(TraesCS1A02G203600.1、TraesCS1D02G249600.1、TraesCS3A02G260100.1、TraesCS6D02G199500.1、TraesCS6A02G216100.1、TraesCS2D02G349400.1、TraesCS2A02G350900.1、TraesCS2A02G350800.1、TraesCSU02G026200.1、TraesCS7B02G256000.1、TraesCS1B02G158000.1、TraesCS1D02G140500.1、TraesCS1A02G141500.1、TraesCS3A02G373800.1、TraesCS3B02G406000.1)的表达均明显上调,说明几丁质酶基因在小麦应答禾谷镰刀菌过程中起重要的作用。

图5 小麦GH-19亚家族成员保守基序分析
3 结论与讨论
本研究从小麦基因组中共鉴定到159个几丁质酶基因,分为GH-18和GH-19两个亚家族,相同亚家族成员几乎具有相似的外显子-内含子基因结构和保守基序,并且聚类在同一分支上。此外,在A、B、D 3套染色体组中鉴定到的几丁质酶基因家族基因数目差别不大,说明几丁质酶基因在A、B、D 3套染色体组长期进化中没有发生明显的基因丢失现象。
近年来,植物几丁质酶基因在很多农作物抗逆抗病过程中具有重要的作用[9-16]。本研究分析了几丁质酶基因家族成员在3个小麦抗病品系和1个小麦感病品系被禾谷镰刀菌侵染前后的表达变化,结果表明,一些几丁质酶基因家族成员的表达明显被禾谷镰刀菌诱导,而且同一家族的不同基因成员的表达量也存在明显的差异。这种现象可能由于基因家族在扩张的过程中同一家族不同基因成员负责不同生物或者非生物胁迫应答功能不同所致。
