性信息素诱捕斜纹夜蛾雄蛾发育日龄结构的研究
2019-11-19陈庆华曾娟杨玲贾勇李庆封传红陈晓娟胡容平
陈庆华,曾娟,杨玲,贾勇,李庆,封传红,陈晓娟,胡容平
性信息素诱捕斜纹夜蛾雄蛾发育日龄结构的研究
陈庆华1,曾娟2,杨玲3,贾勇4,李庆5,封传红6,陈晓娟1,胡容平1
(1四川省农业科学院植物保护研究所/农业农村部西南作物有害生物综合治理重点实验室,成都 610066;2全国农业技术推广服务中心,北京 100125;3农业农村部农产品质量安全中心,北京 100120;4资阳市植保植检站,四川资阳 641300;5四川农业大学农学院,成都 611134;6四川省农业厅植物保护站,成都 610041)
【】判断性信息素诱捕雄蛾发育状况对于客观评价群集诱杀技术的效果具有重要意义。本文通过斜纹夜蛾()内生殖系统形态与发育日龄的研究,判断田间诱捕雄蛾的发育情况和虫源性质,为应用性信息素诱杀技术防控斜纹夜蛾提供理论依据。2014—2015年,利用昆虫解剖镜对室内羽化1—10日龄斜纹夜蛾雄蛾进行解剖测量,形成依据精巢长半轴判断雄蛾日龄的发育标准;2015—2017年,对四川省眉山市东坡区、四川省成都市金堂县田间利用性信息素诱捕到的斜纹夜蛾进行逐日收集并解剖测定,进而依据该标准划分田间逐日性诱的雄蛾日龄结构,分析诱捕雄蛾对自然交配的影响。斜纹夜蛾雄蛾内生殖系统由精巢、输精管、贮精囊、射精管、附腺组成,精巢结构稳定,具有一定弹性,易测量。随着日龄增长,精巢长半轴数值呈减小趋势,1日龄精巢长半轴均值为1 103.54 μm,10日龄雄蛾精巢长半轴比1日龄长半轴减少44.71%,精巢长半轴()与日龄()关系式为:=-48.52+1084(2=0.9472,RMSE=36.8)。1—5日龄组、6—8日龄组、9—10日龄组之间精巢长半轴长度差异显著。根据实验室饲养羽化斜纹夜蛾1—10日龄的精巢长半轴长度与羽化日龄间的数据关系,制定出精巢长半轴判别雄蛾日龄标准表,用于检测田间信息素诱捕雄蛾日龄组成。2015—2017年金堂县和2017年东坡区田间诱捕的雄蛾日龄结构趋势一致,日龄越大,该日龄雄蛾占总诱捕数的百分数越低。金堂县田间诱捕斜纹夜蛾雄蛾1日龄平均百分比为39.89%,2日龄为21.42%,3日龄为15.75%,1—3日龄雄蛾共计占总诱捕数72.22%—79.02%,5—10日龄占比均在10%以下;2017年东坡区田间诱捕斜纹夜蛾雄蛾中1日龄百分比为39.93%,2日龄为20.79%,3日龄为19.54%,1—3日龄雄蛾共计占总诱捕雄蛾数的80.26%,5日龄以后占比较小。各日龄雄蛾平均百分数与日龄数呈指数函数关系下降。根据逐日诱捕雄蛾数量动态,4月末至8月末,金堂县和东坡区斜纹夜蛾均发生3代,第1代、第2代诱捕的雄蛾数以1—3日龄青年蛾为主,金堂青年蛾与老年蛾比为3.32、1.54,东坡区青年蛾与老年蛾比为3.34、1.58;随着虫量增加,第3代东坡区以老龄蛾数量占优势,两者比为0.76,低于金堂的2.34。斜纹夜蛾雄蛾精巢大小能够反映雄蛾日龄,通过检测田间性诱雄蛾日龄的结果表明,利用性信息素捕获的雄蛾以1—3日龄青年雄蛾为主,有利于降低田间成虫交配率。
斜纹夜蛾;雄蛾;内生殖系统;日龄;性信息素诱捕
0 引言
【研究意义】斜纹夜蛾()属鳞翅目(Lepidoptera)夜蛾科(Noctuidae),是一种世界性分布的农业害虫,该虫食性杂、发生范围广、世代重叠,在我国暴发危害严重[1-2]。斜纹夜蛾成虫趋光性较弱,上灯数量少,加之由于杀虫灯容易受周围光源干扰,传统灯光诱杀效果不明显[3]。斜纹夜蛾性信息素由于具有专一性强、高效、无污染等特点而受到广泛关注,诸多研究明确了其性信息素活性成分组成[4-5]、作用机理[6]、使用方法和田间效果[7-10]、诱捕器设计[11]等,为使用性信息素控制斜纹夜蛾发生动态提供了支撑。利用性信息素诱捕雄蛾(下文简称“性诱”),减少雄、雌交配次数,以达到降低雌蛾产卵量和有效卵量的目的。但如果诱捕的雄蛾为交配后的雄蛾或者是老弱雄蛾,利用性信息素控制害虫就失去了意义。通过观察雄蛾内生殖系统形态变化来分析发育日龄,判断诱捕斜纹夜蛾雄蛾发育状况[12],可为科学评价利用性信息素控制害虫的防治效果提供依据。【前人研究进展】根据对昆虫内生殖系统特征的解剖观察可以判别昆虫的发育阶段。鳞翅目成虫的发育级别可以通过解剖雌成虫卵巢来判断,通常根据雌虫卵粒的成熟情况、颜色及脂肪的多寡等内部结构特征,判断其不同的发育级别[13-14]。在科研工作者大量的研究中,形成了卵巢发育的分级标准。一般鳞翅目害虫卵巢分为5级,如黏虫()[15]、甜菜夜蛾()[16]、稻纵卷叶螟()[17]、棉铃虫()[18]、斜纹夜蛾[19]等,1、2级为未交配状态,3—5级为老熟蛾[19]。对鳞翅目雄蛾内生殖系统的研究多是以观察形态学和组织学特征为主,如茶尺蠖()[20]、墨西哥水稻螟()[21]、沙棘木蠹蛾()[22]、二化螟()[23]、斜纹夜蛾[24]、黏虫[15]等。通过对二化螟雄蛾内生殖系统的比较研究,发现其内生殖器内含物特征可以区别二化螟雄蛾交配与否[25]。斜纹夜蛾成虫日龄对交配行为影响较大,雌雄交配主要发生在羽化后的1、2 d[26]。棉褐环野螟()成虫不同日龄与交配状态组合对繁殖力具有一定的影响[27];稻纵卷叶螟雄蛾发育日龄与精巢大小密切相关[12]。【本研究切入点】不同于幼虫随着发育而脱皮,昆虫成虫的体形基本保持稳定,很难从雄蛾外部形态的变化得出发育日龄。本文从内生殖系统解剖角度出发,利用斜纹夜蛾雄蛾个体精巢长半轴变化,推测雄蛾日龄,再根据斜纹夜蛾日龄与交配节律的研究评价对群体交配的影响。【拟解决的关键问题】对斜纹夜蛾雄蛾内生殖系统进行解剖,得到精巢与日龄的发育标准,进而依据该标准划分田间逐日性诱的雄蛾日龄,获得青年雄蛾在诱捕中的比例,评价性诱控害作用。
1 材料与方法
1.1 供试虫源
实验室饲养虫源:2014—2015年由四川农业大学提供斜纹夜蛾蛹,用于研究精巢大小与日龄关系;2016年由四川省农业科学院提供斜纹夜蛾蛹,用于研究精巢大小与体长关系。将供试蛹放入铺有棉花和纱布的饲养盒内待羽化,羽化后即转移至饲养笼内,用消毒的脱脂棉浸10%蜂蜜水悬挂于笼中,为成虫提供营养。
田间昆虫:2015—2017年在四川省成都市金堂县蔬菜基地利用性诱采集,2017年在四川省眉山市东坡区蔬菜基地采集。每日观察并记录收集虫量。斜纹夜蛾性诱剂诱芯和诱捕器均由宁波纽康生物技术有限公司生产。
1.2 测量及精巢解剖工具、方法
用游标卡尺测量斜纹夜蛾体长,用昆虫解剖镜(Motic:Digital Microscope SMZ171)解剖并测量精巢长/短半轴,方法参照陈庆华等[12]。田间日诱捕雄蛾数量超过50头,选择50头进行解剖,再按照诱捕数折算各日龄比例。
1.3 数据统计与分析
日龄划分方法:某日龄精巢长半轴下限=本龄精巢长半轴的均值-标准差;某日龄精巢长半轴上限=上一龄精巢长半轴下限。如1日龄下限=1日龄精巢长半轴均值-标准差;2日龄精巢长半轴下限=2日龄精巢长半轴均值-标准差;2日龄精巢长半轴上限=1日龄精巢长半轴下限。以此类推,直到第10日龄,10日龄精巢长半轴上限=9日龄精巢长半轴下限。
试验中各日龄精巢长半轴均值、方差分析、多重比较、体长与精巢的相关系数计算、田间不同日龄雄蛾占比与日龄数拟合均采用MathWorks公司Matlab R2014a软件进行。
2 结果
2.1 斜纹夜蛾雄蛾内生殖系统器官组成
斜纹夜蛾雄蛾内生殖系统由精巢、输精管、贮精囊、射精管、附腺组成(图1)。其形态特征为精巢1个,球形或椭圆形,颜色橘色,精巢下端有2根输精管,连接贮精囊。精巢结构稳定,具有一定弹性,易测量。贮精囊,1对,自输精管中间膨大,交叉形成x形,囊内呈乳白色。输精管1对,分别连接于贮精囊的上下两端。射精管1条,折叠于腹腔内的极长的透明管道,上端与输精管连接,末端接于阳茎。附腺1对,两管紧贴并折叠在一起,一端与输精管连接,另一端角质化并封闭,管壁为乳白色。
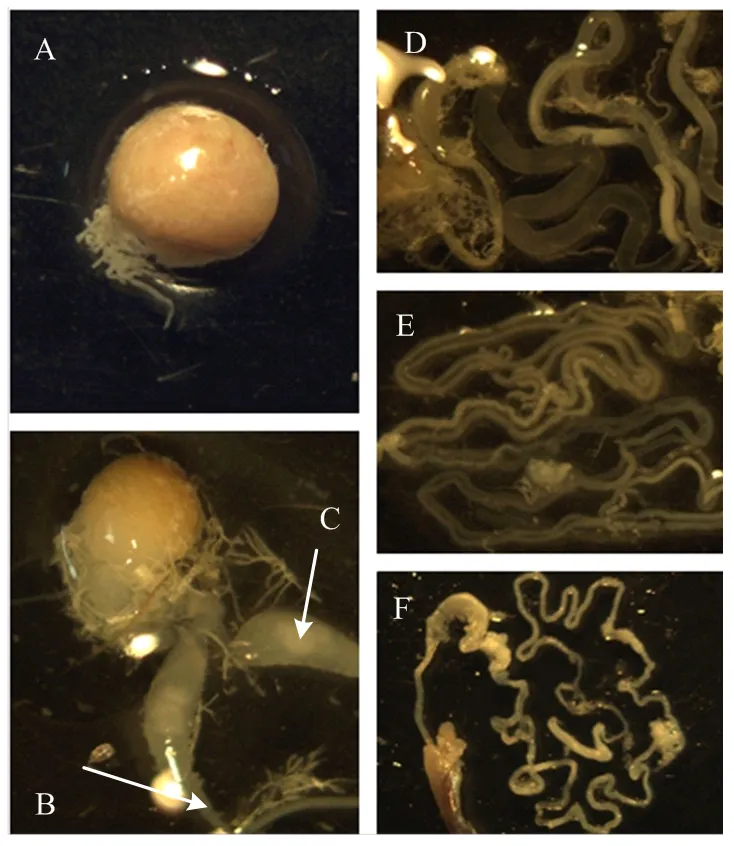
A:精巢Testis;B:输精管Vas deferens;C:贮精囊Vesicles;D、F:射精管Ejaculatory duct;E:附腺Accessory gland
2.2 斜纹夜蛾雄蛾精巢标准的制定
解剖羽化1—10日龄斜纹夜蛾雄蛾共180头,测量分析各日龄精巢长半轴数值表明,随着日龄增长,精巢长半轴数值呈减小趋势(图2),1日龄精巢长半轴均值为1 103.54 μm,2日龄精巢长半轴为均值987.35 μm,直至10日龄精巢长半轴均值为610.29 μm,比1日龄减小44.71%。1—5日龄组、6—8日龄组、9—10日龄组之间精巢长半轴长度差异显著。精巢长半轴()与日龄()关系式为:=-48.52+1084(2=0.9472,RMSE=36.8)。斜纹夜蛾1—10日龄雄蛾精巢长半轴长度区间划分见表1。
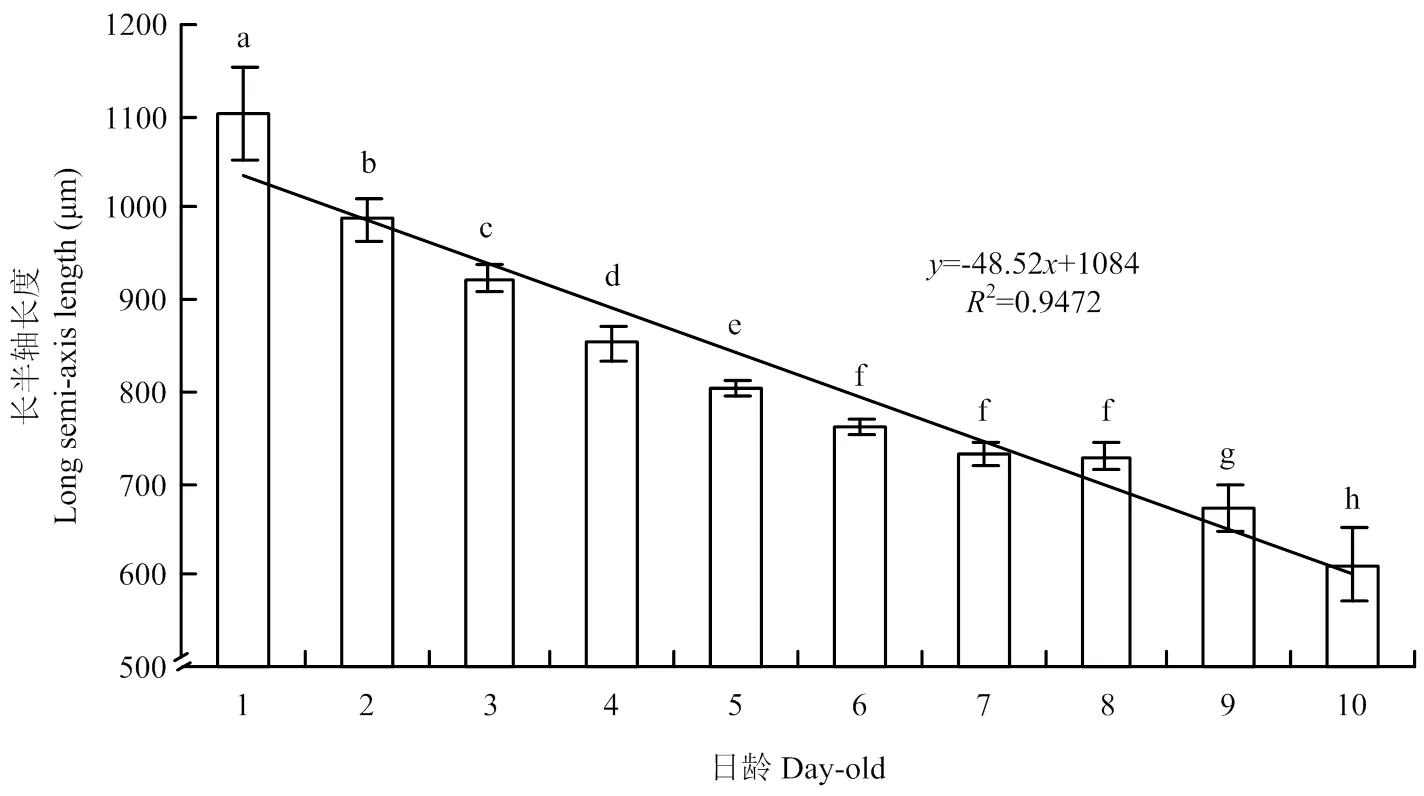
图中条形柱上短线为标准差,条形柱上不同小写字母表示各日龄组之间差异显著(P<0.01,One-way ANOVA)
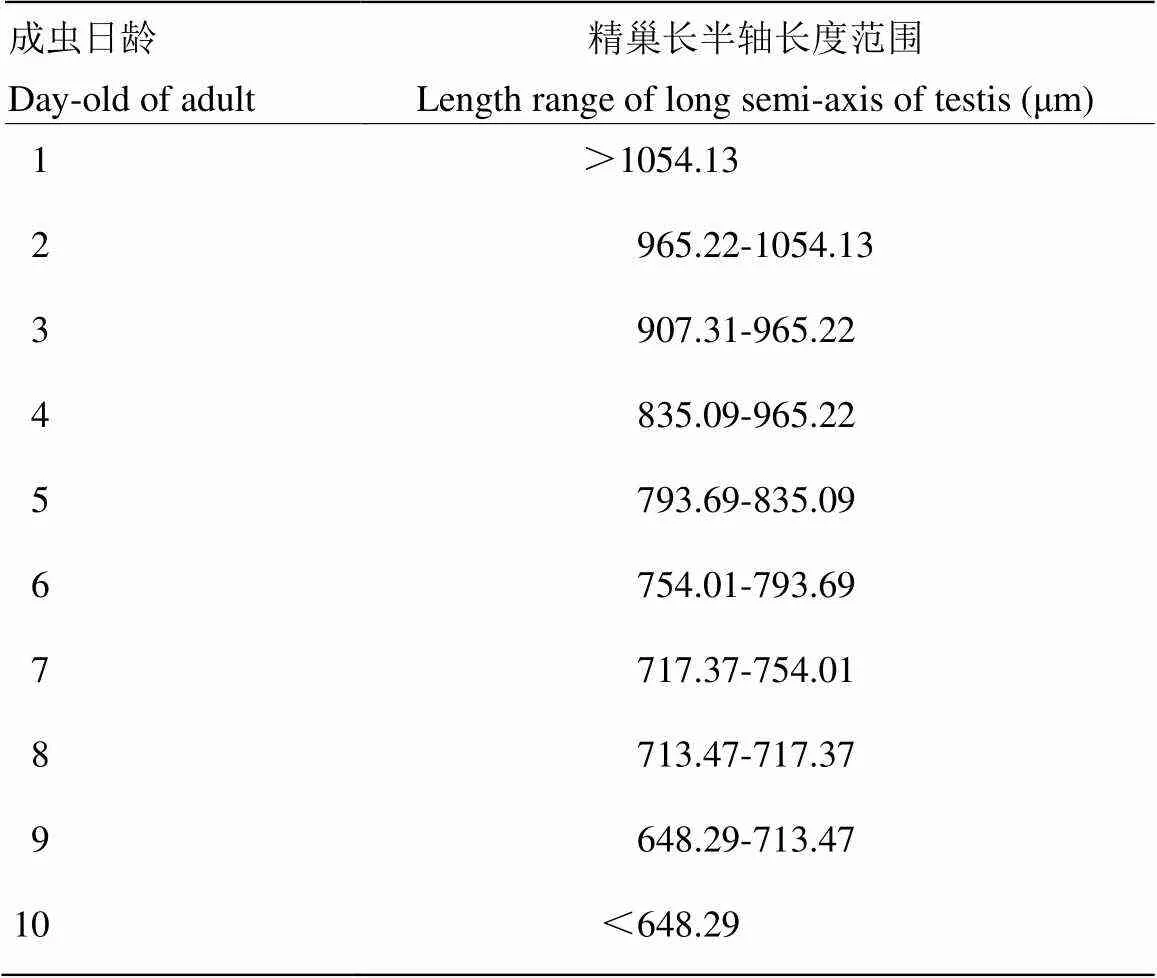
表1 斜纹夜蛾1—10日龄雄蛾精巢长半轴长度划分指标
2.3 田间性信息素诱捕雄蛾日龄的比例
2015—2017年金堂县和东坡区田间诱捕的雄蛾日龄组成趋势一致,日龄越大,占比越低。2015年在金堂县用性信息素诱捕斜纹夜蛾为180头,1日龄占整个诱虫量的40.00%,1—3日龄虫量占总虫量的72.22%,5日龄以后虫量占总虫量的14.44%。2016年在金堂县用性信息素诱捕斜纹夜蛾为205头,1日龄占总虫量的40.49%,1—3日龄虫量占总虫量的79.02%,5日龄以后虫量占总虫量的8.78%。2017年分别在东坡区和金堂县逐日诱捕斜纹夜蛾,东坡区共诱捕1 520头雄蛾,1日龄占39.93%,1—3日龄占80.26%。金堂县共诱捕963头雄蛾,1日龄占39.15%,1—3日龄占76.74%,与2015、2016年趋势吻合。利用非线性最小二乘法,算法为trust-region,对各日龄雄蛾所占平均百分数()与日龄()进行指数函数关系拟合:=63.64×e-0.488x(2=0.9838,RMSE=1.718)(图3)。
2.4 田间诱捕雄蛾日龄组成逐日动态
图4为1—3日龄与4—10日龄雄蛾数与总雄蛾数的逐日动态图,两地均有3个发蛾期,金堂县第1代(5月8日至7月4日)1—3日龄青年蛾与4—10日龄老年蛾之比为3.32,第2代(7月5日至8月5日)比为1.54,第3代(8月6日至8月31日)比为2.34。东坡区3个发蛾期的青年蛾与老年蛾之比分别为3.34、1.58、0.76。

图3 2015—2017金堂县、2017年东坡区田间性诱斜纹夜蛾不同日龄雄蛾比例
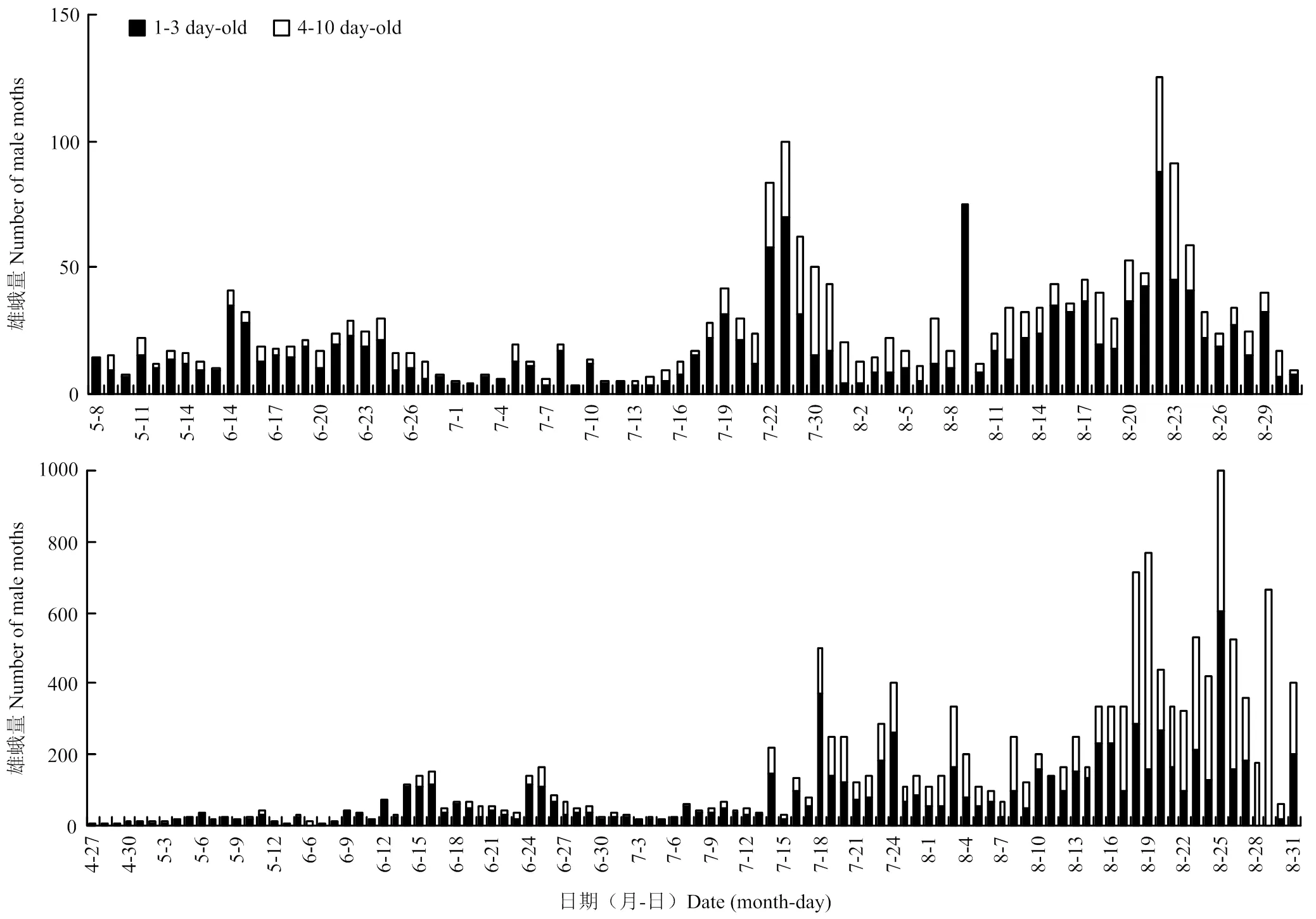
图4 2017年金堂县(上)、东坡区(下)田间逐日性诱斜纹夜蛾1—3日龄和4—10日龄动态
3 讨论
成虫日龄、体长、精巢大小、进攻器官与雌虫的交配选择有一定关系[28-30],然而成虫体长与精巢大小之间并没有明显关系。试验测试1日龄雄蛾精巢长半轴(,μm)与体长(,mm)的关系式为:=18.825+990.8(2=0.0373,=0.0464),两者呈弱相关。试验用虫体长标准差为1.15 mm,折算成精巢长半轴为21.80 μm,而精巢长半轴从1日龄到2日龄减少116.19 μm,远高于体长的影响值。从日龄分别与体长和精巢大小变化来看,斜纹夜蛾1日龄雄蛾体长为18.08 mm,雄蛾1日龄体长比10日龄长0.92 mm,10 d缩短5.1%,而精巢长半轴随日龄的变化44.71%,因此体长不能判断雄蛾发育日龄。
昆虫的交配行为受到多种外部因素的影响,如环境温度和相对湿度、光强度、光周期等[31-34],但自然条件下,斜纹夜蛾雄蛾日龄是主要影响因素。斜纹夜蛾成虫生命只有10 d左右[35],老年雄蛾精子数量和活力会下降,产生有害突变[36],交配日龄越早,种群成功繁衍风险越小。在实验室配对的情况下,斜纹夜蛾交配行为主要发生在1—2日龄,3龄之后极少发生[26]。本文通过对3年两地的田间诱捕的雄蛾发育日龄进行检测表明,1—2日龄共占总虫量的61.31%,1—4日龄的雄蛾占总虫量的90%左右。更多的富有活力的年青雄蛾被诱捕,可以直接减少斜纹夜蛾雌蛾的交配率,加之没有被诱捕的雄蛾也会受到干扰,造成了一定程度的交配阻隔,实现了控制田间斜纹夜蛾子代种群数量的目的。田间试验表明,性信息素诱杀可明显地降低大豆田斜纹夜蛾交配率,连续2个月内雌蛾交配率仅为40%左右,而对照为80%以上[37]。
当存在精子竞争时,精巢的大小与物种进化关系密切。如果持续地诱杀具有大精巢个体,田间具有小精巢个体将占优势。对棘突动物、索伊羊的观察表明,更大的精巢能够增加产生精子的受精成功率[38-39]。本试验中具有较大精巢的低日龄雄蛾被诱捕,田间剩余的是具有较小精巢雄蛾,雌蛾的受精成功率也相应降低,下一代种群数量和质量均下降。在实际应用中发现,连续几年应用性诱控害后,一些害虫自然发生程度有下降的趋势[10],除了避免杀伤了天敌的因素外,上述因素也可能是原因之一。
本研究发现当田间斜纹夜蛾1代、2代发生时,田间密度较低,金堂县、东坡区青年蛾与老年蛾比例接近,1代分别为3.32和3.34,2代分别为1.54和1.58;3代发生时,东坡区田间斜纹夜蛾虫量明显高于1、2代及金堂虫量,诱捕器中老年蛾占优势。这种现象与二化螟类似,利用性信息素诱杀二化螟越冬代的效果要好于其他代次[40],随着雌虫密度的增加,田间雌蛾会与性信息素相互竞争,从而导致防治效果下降[41]。因此,利用性信息素诱杀技术适合于低密度时的害虫防控。
4 结论
雄蛾精巢大小与发育日龄显著相关,根据精巢大小与雄蛾日龄关系检测性诱雄蛾日龄的结果表明,利用田间性信息素捕获的雄蛾以青年雄蛾为主,1—3日龄雄蛾占70%以上,有利于降低田间成虫交配次数。
致谢:本项研究得到国家公益性行业(农业)科研专项“昆虫性诱剂合成与缓释技术研究”的大力资助。参加项目的研究人员还有郑仕军、唐裕智、胡远华、陈霞等。在项目实施过程中,有关市县植保技术推广部门给予了积极配合和大力协助,在此一并致谢!
[1] 周晓梅, 黄炳球. 斜纹夜蛾抗药性及其防治对策的研究进展. 昆虫知识, 2002, 39(2): 98-102.
ZHOU X M, HUANG B Q. Insecticide resistance of the common cutworm() and its control strategies., 2002, 39(2): 98-102. (in Chinese)
[2] 杨新军, 林华峰, 鲍晓莉. 6种寄主植物对斜纹夜蛾生长发育的影响. 中国植保导刊, 2006, 26(9): 30-31.
YANG X J, LIN H F, BAO X L.Effects of six host plants on the growth and development of, 2006, 26(9): 30-31. (in Chinese)
[3] 曾娟, 杜永均, 姜玉英, 刘万才. 我国农业害虫性诱监测技术的开发和应用. 植物保护, 2015, 41(4): 9-15.
ZENG J, DU Y J, JIANG Y Y, LIU W C. Development and application of sex pheromone in population monitoring of crop insect pests in China., 2015, 41(4): 9-15. (in Chinese)
[4] TAMAKI Y, YUSHIMA T. Biological activity of the synthesized sex pheromone and its geometrical isomers of(F.) (Lepidoptera: Noctuidae)., 1974, 9(2): 73-79.
[5] YUSHIMA T, NOGUCHI H, TAMAKI Y, FUKAZAWA N, SUGINO T. Mating and sex pheromone ofF. (Lepidoptera: Noctuidae): An introductory report., 1973, 8(1): 18-26.
[6] 孙凡, 胡隐月, 杜家纬. 斜纹夜蛾性信息素通讯系统. 昆虫学报, 2002, 45(3): 404-407.
SUN F, HU Y Y, DU J W. The sex pheromone communication system of(Fabricius)., 2002, 45(3): 404-407. (in Chinese)
[7] KAWASAKI K, MIYASHITA K. Mating suppression by individual components of the sex pheromone ofF. (Lepidoptera: Noctuidae) under field conditions., 1976, 11(4): 320-326.
[8] NEMOTO H, TAKAHASHI K, KUBOTA A. Reduction of the population density of(F.) (Lepidoptera: Noctuidae) using a synthetic sex pheromone. I. experiment in taro field., 1980, 24(4): 211-216. (in Japanese)
[9] HIRANO C. Evaluation of survey trapping locations for(Lepidoptera: Noctuidae) with sex pheromone traps., 1981, 25(4): 272-275. (in Japanese)
[10] SHOBU S, MIKURIYA H, KONDO T. Relationship between adult pheromone trap catches and injury in soybean leaves by young larvae ofF. (Lepidoptera: Noctuidae)., 2003, 47(4): 137-141.
[11] RANGA RAO G V, WIGHTMAN J A, RANGA RAO D V. The development of a standard pheromone trapping procedure for(F.) (Lepidoptera: Noctuidae) population in groundnut (L.) crops., 1991, 37(1): 37-40.
[12] 陈庆华, 曾娟, 曾伟, 李庆, 陈晓娟, 邹禹. 稻纵卷叶螟雄蛾内生殖系统形态指标在性诱测报中的应用. 昆虫学报, 2017, 60(8): 927-935.
CHEN Q H, ZENG J, ZENG W, LI Q, CHEN X J, ZOU Y. Application of the morphological indicators of the male internal reproductive system in forecasting the population dynamics of the rice leaf roller,(Lepidoptera: Pyralidae) by sex pheromone trapping., 2017, 60(8): 927-935. (in Chinese)
[13] 齐国君, 芦芳, 胡高, 王凤英, 高燕, 吕利华. 卵巢解剖在我国迁飞昆虫研究中的应用. 中国植保导刊, 2011, 31(7): 18-22.
QI G J, LU F, HU G, WANG F Y, GAO Y, Lü L H. The application of ovarian dissection in the research on migratory insects in China.,2011, 31(7): 18-22. (in Chinese)
[14] 关瑞峰, 姚文辉, 王茂明, 孔丽萍, 潘初昕, 徐金汉. 水稻迁飞性害虫卵巢解剖及虫源性质分析. 福建农业科技, 2008(3): 30-32.
GUAN R F, YAO W H, WANG M M, KONG L P, PAN C X, XU J H. Analysis of ovary anatomy and insect origin of rice migratory pests., 2008(3): 30-32. (in Chinese)
[15] 石毓亮, 张之光, 李玉椿. 粘虫生殖系统解剖与自然交配及其在实践中的应用. 山东农学院学报(自然科学版), 1964, 9: 45-56.
SHI Y L, ZHANG Z G, LI Y C. Anatomy and natural mating of reproductive system of(Walker) and its application in practice., 1964, 9: 45-56. (in Chinese)
[16] 王宪辉, 徐洪富, 许永玉, 刘勇, 周真. 甜菜夜蛾雌性生殖系统结构、发育分级及在测报上的应用. 植物保护学报, 2003, 30(3): 261-266.
WANG X H, XU H F, XU Y Y, LIU Y, ZHOU Z. The structures and developmental progress of reproductive system of beet armyworm,(Hübner), and their use in forecast., 2003, 30(3): 261-266. (in Chinese)
[17] 张孝羲, 陆自强, 耿济国. 稻纵卷叶螟雌蛾解剖在测报上的应用. 昆虫知识, 1979,16(3): 97-99.
ZHANG X X, LU Z Q, GENG J G. Application of anatomy of female moths ofin forecasting., 1979, 16(3): 97-99. (in Chinese)
[18] 张韵梅, 牟吉元. 棉铃虫卵巢发育的组织化学及测报分级研究. 山东农业科学, 1994(3): 7-9.
ZHANG Y M, MU J Y.Histochemical and predictive classification of ovarian development of(Hübner)., 1994(3): 7-9. (in Chinese)
[19] 武怀恒, 万鹏, 黄民松. 斜纹夜蛾雌蛾生殖系统发育的形态变化. 湖北农业科学, 2012, 51(24): 5654-5657.
WU H H, WAN P, HUANG M S. Morphological change on the development of female reproductive system in(Fabricius)., 2012, 51(24): 5654-5657. (in Chinese)
[20] 张家侠, 孙钦玉, 赵强, 夏先江, 丁勇. 茶尺蠖雄成虫生殖系统形态学与组织学观察. 茶叶科学, 2015, 35(6): 527-533.
ZHANG J X, SUN Q Y, ZHAO Q, XIA X J, DING Y. Morphological and histological observations on the reproductive system in male adults ofProut., 2015, 35(6): 527-533. (in Chinese)
[21] SPURGEON D W, RAULSTON J R, LINGREN P D, SHAVER T N, PROSHOLD F I, GILLESPIE J M. Temporal aspects of sperm transfer and spermatophore condition in Mexican rice borers (Lepidoptera: Pyralidae)., 1994, 87(2): 371-376.
[22] 周娇, 李娟, 翁强, 骆有庆. 沙棘木蠹蛾生殖系统形态学和组织学观察. 应用昆虫学报, 2012, 49(6): 1629-1635.
ZHOU J, LI J, WENG Q, LUO Y Q. Morphological and histological observations on the reproductive system of, 2012, 49(6): 1629-1635. (in Chinese)
[23] 宋俊元, 胡阳, 傅强, 郑永利. 二化螟雄虫内生殖系统结构的解剖. 浙江农业科学, 2012(5): 686-688.
SONG J Y, HU Y, FU Q, ZHENG Y L. Anatomy of male internal reproductive system ofWalker., 2012(5): 686-688. (in Chinese)
[24] 徐博. 斜纹夜蛾前胸腺与性腺的形态结构和发育研究[D]. 广州: 中山大学, 2012.
XU B. Research on the morphological structure and development ofprothoracic glands and gonads[D]. Guangzhou: Sun Yat-sen University, 2012. (in Chinese)
[25] 郭前爽, 王春荣, 宋显东, 冯波, 陈庆华, 杜永均. 日龄和交配状态对二化螟雄蛾内生殖器特征的影响. 昆虫学报, 2019, 62(7): 838-848.
GUO Q S, WANG C R, SONG X D, FENG B, CHEN Q H, DU Y J. Effects of age and mating status on the characteristics of internal genital organs of male adults of the striped rice stem borer,(Lepidoptera: Crambidae)., 2019, 62(7) 838-848. (in Chinese)
[26] 武承旭, 杨茂发, 姚苗苗, 曾昭华. 斜纹夜蛾雌雄配对后成虫活动行为节律及时间分配. 植物保护学报, 2016, 43(6): 979-985.
WU C X, YANG M F, YAO M M, ZENG Z H. Diurnal activity rhythm and time budgets of adult behavior of(Fabricius) (Lepidoptera: Noctuidae) after paired., 2016, 43(6): 979-985. (in Chinese)
[27] 张清泉, 张雪丽, 陆温. 成虫日龄与交配状态对棉褐环野螟繁殖力的影响. 植物保护, 2012, 38(2): 71-74.
ZHANG Q Q, ZHANG X L, LU W. Effects of adult age and mating status on fecundity of., 2012, 38(2): 71-74. (in Chinese)
[28] KELLY C D. Sperm investment in relation to weapon size in a male trimorphic insect?, 2008, 19(5): 1018-1024.
[29] Moore P J, Moore A J. Reproductive aging and mating: the ticking of the biological clock in female cockroaches., 2001, 98(16): 9171-9176.
[30] Biedermann R. Mating success in the spittlebug(Scopoli, 1763) (Homoptera, Cercopidae): the role of body size and mobility., 2002, 20(1): 13-18.
[31] Kanno H. Mating behavior of the rice stem borer moth,Walker (Lepidoptera: Pyralidae). IV. Threshold-light- intensity for mating initiation under various temperature., 1980, 15(4): 372-377.
[32] Kanno H, Sato A. Mating behavior of the rice borer moth,Walker (Lepidoptera: Pyralidae ). Ⅲ. Joint action of temperature and relative humidity on mating activity., 1980, 15(1): 111-112.
[33] 余棋, 闫祺, 董双林. 温度对二化螟性信息素通讯的影响. 应用生态学报, 2017, 28(11): 3784-3788.
YU Q,YAN Q, DONG S L. Effect of temperature on the sex pheromone communication ofWalker (Lepidoptera: Pyralidae)., 2017, 28(11): 3784-3788. (in Chinese)
[34] 邱小芳, 谢建坤, 吴丽萍, 喻琴, 张萌萌, 王光耀, 徐韶晖, 涂小云. 光周期变化对毛健夜蛾交配和产卵的影响. 昆虫学报, 2016, 59(10): 1103-1114.
QIU X F, XIE J K, WU L P, YU Q, ZHANG M M, WANG G Y, XU S H, TU X Y. Effects of varied photoperiods on the mating and oviposition of(Lepidoptera: Noctuidae) adults., 2016, 59(10): 1103-1114. (in Chinese)
[35] 武承旭, 陈俊, 黄康, 廖启荣. 斜纹夜蛾雌虫寿命及产卵量的影响因子研究. 广东农业科学, 2012(17): 68-70.
WU C X, CHEN J, HUANG K, LIAO Q R. Affecting factors of longevity and fecundity offemale., 2012(17): 68-70. (in Chinese)
[36] Hansen T F, Price D K. Good genes and old age: do old mates provide superior genes?, 1995, 8(6): 759-778.
[37] Kitamura C, Kobayashi M. A comparison between communication disruption and mass trapping methods in mating suppression effect of a synthetic sex pheromone toF. (Lepidoptera: Noctuidae)., 1985, 20(2): 222-224.
[38] Poulin R, Morand S. Testes size, body size and male-male competition in acanthocephalan parasites., 2000, 250: 551-558.
[39] Preston B T, Stevenson I R, Pemberton J M, Coltman D W, Wilson K. Overt and covert competition in a promiscuous mammal: the importance of weaponry and testes size to male reproductive success., 2003, 270: 633-640.
[40] 焦晓国, 宣维健, 王红托, 苏建伟, 盛承发. 水稻二化螟性信息素防治研究进展. 昆虫知识, 2003, 40(3): 193-199.
JIAO X G, XUAN W J, WANG H T, SU J W, SHENG C F. Advances in the use of sex pheromone to control., 2003, 40(3): 193-199. (in Chinese)
[41] KONDO A, TANAKA F. Effect of wild virgin females on pheromone trap efficiency in two annual generations of the rice stem borer moth,(Walker) (Lepidoptera: Pyralidae)., 1994, 29(2): 279-281.
The Structure of Day-Age of MaleMoths by Sex Pheromone Trapping in the Field
CHEN QingHua1, ZENG Juan2, YANG Ling3, JIA Yong4, LI Qing5, Feng ChuanHong6, CHEN XiaoJuan1, HU RongPing1
(1Institute of Plant Protection, Sichuan Academy of Agricultural Sciences/Key Laboratory of Integrated Pest Management on Crops in Southwest, Ministry of Agriculture and Rural Affairs, Chengdu 610066;2National Agro-Technical Extension and Service Centre, Beijing 100125;3Center for Agro-Food Quality and Safety, Ministry of Agriculture and Rural Affairs, Beijing 100120;4Plant Protectionand Plant Quarantine Station of Ziyang City, Ziyang 641300, Sichuan;5College of Agronomy, Sichuan Agricultural University, Chengdu 611134;6Plant Protection Station of Sichuan province, chengdu 610041)
【】It is of great significance to determine the developmental status of male moths trapped by sex pheromone for objective evaluation of the effect of mass trapping. This study aims to research the internal reproductive system and developmental age of, determine the development situation of male moths captured in the field and the nature of insect source, and to provide a theoretical basis for the application of sex pheromone trapping of【】In 2014 and 2015, after eclosion from the laboratory-reared pupae, 1-10 day-old male moths were dissected under the microscope. The criterion of the developmental age of malemoths was determined by the long semi-axis of testis. Using this criterion, the day-old of male moths captured daily with sex pheromone trapping in the fields of Dongpo and Jintang counties of Sichuan Province from 2015 to 2017 was identified. The effect of trapping male moths on mating behaviors and patterns was then analyzed.【】The internal reproductive system of malemoths was composed of testis, vas deferens, seminal vesicle, ejaculatory duct and accessory gland. The structure of testis was stable, elastic and easily-measured. With the increase of age, the length of long semi-axis of testis gradually decreased. The mean value of long semi-axis of testis of 10-day-old males decreased by 44.71%, compared with that of 1-day-old male testis (1 103.54 μm). The relationship equation between the long semi-axis () and day-old () was=-48.52+1084 (2=0.9472, RMSE=36.8).There were significant differences in the length of long semi-axis of testis among the age groups of 1-5, 6-8 and 9-10 day-old. The criterion table displaying the relationship between the day-old of male moths and the length of long semi-axis of testis was built to identify the age of male moths captured by pheromone trapping in the field.Similar day-age structures of male moths were found in Jintang from 2015 to 2017 and in Dongpo in 2017, with lower percentages of older male moths. Specifically, the mean percentage of 1-day-old male moths captured by sex pheromone trapping in Jintang was 39.89%, and that of 2- and 3 day-old was 21.42% and 15.75%, respectively. In other words, 1-3 day-old male moths accounted for 72.22%-79.02% in total, and the percentage from 5- to 10-day-old male moths was below 10%.In Dongpo, the percentage of 1-, 2- and 3-day-old male moths was 39.93%, 20.79% and 19.54%, respectively, as a total of 80.26%, after 5-day-old of age, the percentage was relatively small. The mean percentage of male moths of each day-old decreased exponentially with age. The daily trapping dynamic showed the 3 generations of adults in the fields of Jintang and Dongpo from the end of April to the end of August. The male moths of the 1st and 2nd generations were mainly between ages of 1-3 day-old, and the ratio of younger moths (1-3 day-old) of Jintang to older moths (4-10 day-old) was 3.32 and 1.54, and that of younger moth to older moths in Dongpo was 3.34 and 1.58, respectively. With increased numbers of adults in Dongpo, the majority of 3rd generation was older moths, with the ratio of younger moths to older moths of 0.76, which was lower than that of Jintang (2.34).【】The size of the testis of malemoth can reflect its day-age. Male moths captured through sex pheromone trapping are mainly composed of 1-3 day-old, which is beneficial to reduce mating rate in the current generation and the whole population in the next ones.
; male moth; internal reproductive system; day-old; sex pheromone trapping
10.3864/j.issn.0578-1752.2019.21.010
2019-05-20;
2019-06-25
国家公益性行业(农业)科研专项(201203036)、中央引导地方科技发展专项(2019ZYD099)
陈庆华,E-mail:710478145@qq.com。曾娟,E-mail:zengjuan@agri.gov.cn。陈庆华和曾娟为同等贡献作者。通信作者胡容平,E-mail:44573780@qq.com
(责任编辑 岳梅)
