甘薯抗病虫相关基因SGT1的TRAP分析
2019-11-05王崇杨新笋雷剑苏文瑾柴沙沙张文英王连军
王崇 杨新笋 雷剑 苏文瑾 柴沙沙 张文英 王连军
摘要:以抗病虫相关基因SGT1作为靶标基因,设计出1条固定引物,与11条随机引物组合。利用这11对引物对试验材料鄂薯11和鄂紫薯13进行TRAP-PCR扩增,发现11对引物组合扩增出清晰条带,且表现出良好的多态性。研究获得与SGT1基因相关联的TRAP标记,该标记可进行甘薯[Ipomoea batatas(L.) Lam.]群体的遗传多样性分析,为甘薯的抗病虫育种提供理论依据。
关键词:甘薯[Ipomoea batatas(L.) Lam.];抗病虫;SGT1基因;TRAP标记
中图分类号:Q789;S531 文献标识码:A
文章编号:0439-8114(2019)17-0119-04
DOI:10.14088/j.cnki.issn0439-8114.2019.17.032 开放科学(资源服务)标识码(OSID):
Abstract: Disease and pest resistance related gene SGT1 was used as a target gene to design a fixed primer,then for paring 11 pairs primers with 11 arbitrary primers. The 11 pairs of primers were used to amplify Eshu 11 and Ezishu 13 by TRAP-PCR. It was found that 11 pairs of primer combinations amplified clear bands and showed good polymorphism. In this study, TRAP markers associated with SGT1 gene were obtained, which can be used to analyze the genetic diversity of sweetpotato[Ipomoea batatas(L.) Lam.] population, and provide theoretical basis for disease and insect resistance breeding of sweetpotato.
Key words: sweetpotato[Ipomoea batatas(L.) Lam.]; disease and insect resistance; SGT1 gene; TRAP marker
甘薯[Ipomoea batatas(L.) Lam.]是世界上重要的粮食、饲料、工业原料及新型能源用块根作物,中国是世界上最大的甘薯生产国,常年种植面积550万hm2,鲜薯产量约1.2亿t,分别占世界甘薯种植总面积和总产量的60%和85%[1]。甘薯在遗传上高度杂合,种内、种间杂交不亲和以及遗传资源匮乏,遗传基础狭窄,病虫害、病毒病危害严重,严重制约了甘薯品种的遗传改良[2]。在目前的甘薯育种中,亟须开发出能够与目标性状或相关基因紧密连锁的分子标记。
SGT1(Suppressor of the G2 allele of skp1)基因主要功能是调控着丝粒的装配,并调节泛素对目标靶蛋白的修饰[3]。研究发现,SGT1基因发生沉默或基因突变,植物对R基因引起的病害抗性变弱,如果SGT1基因在植物体内过表达,则植物对病原微生物的抗性有所增强[4]。在植物体内,SGT1与HSP90(Heat shock protein,热激蛋白90)和RAR1(Required for Mla12 resistance)形成复合体,在抗病信号转导中扮演重要角色[5,6]。虽然抗病调控相关基因SGT1不是起着直接作用,但是它在植物抗病中占有不可或缺的地位[7]。SGT1对细菌、真菌、病毒和线虫等病原均具有抗病反应,起到重要的调控作用[8-10]。
TRAP技术[11]是由SRAP技术[12]发展而来,具有简单、高效、重复性好、效率高等优点,已在菜豆[13]、小麦[14]、烟草[15]、棉花[16]等多种作物中得到了广泛应用。本研究以抗病基因作为靶标基因,开发设计TRAP标记引物,旨在评价分析TRAP标记技术在甘薯研究中的利用价值,发掘出有效的与SGT1紧密连锁的TRAP标记,为筛选甘薯抗病品种提供一定的理论依据,为甘薯抗病虫遗传改良奠定基础。
1 材料与方法
1.1 材料
供试材料为高抗蔓割病、抗根腐病和茎线虫病、感黑斑病的甘薯品种鄂薯11;感蔓割病、高感根腐病、中抗茎线虫病、高感黑斑病的甘薯品种鄂紫薯13。通过海南和武汉两地田间调查小象甲危害指数,鄂薯11受危害程度比鄂紫薯13輕。鄂薯11和鄂紫薯13种植于湖北省农业科学院粮食作物研究所试验田。
1.2 引物设计
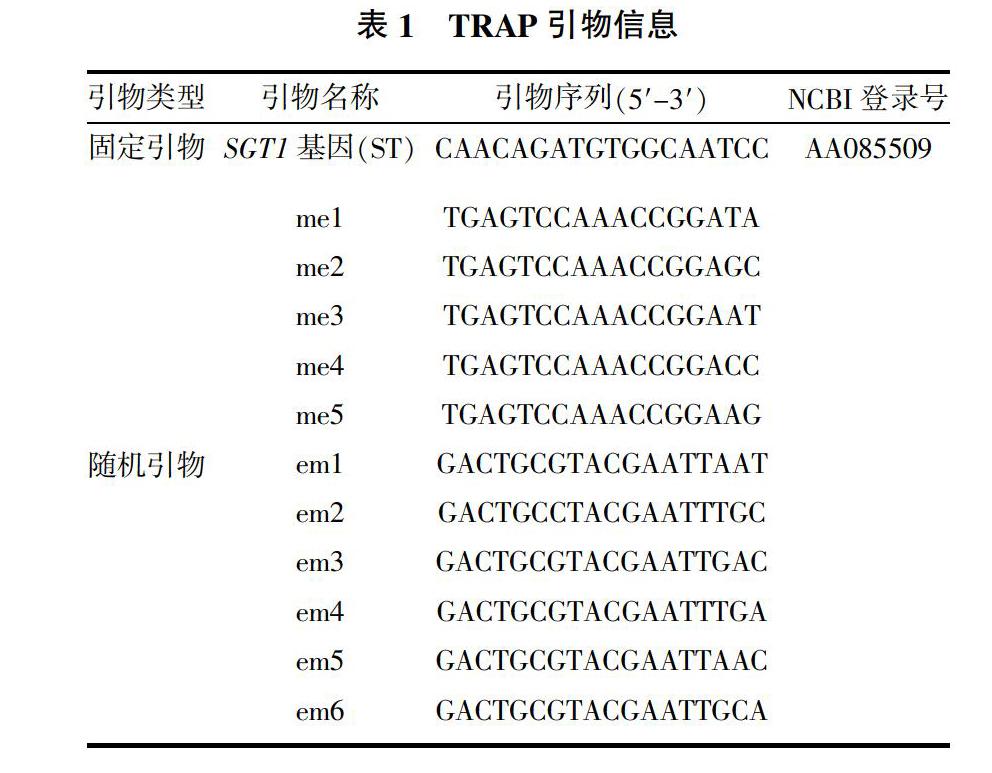
利用Premier 5软件设计引物。根据NCBI登录号搜索到SGT1基因序列,以SGT1基因作为靶标基因设计1条固定引物,固定引物大小为18 bp,退火温度为50 ℃,参考引用Li等[17]已发表的11条随机引物(表1),均委托天一辉远生物科技有限公司(武汉)合成。
1.3 DNA的提取
采取鄂薯11、鄂紫薯13的新鲜幼嫩叶片,采用改良CTAB法[18]提取甘薯总DNA。将DNA稀释到PCR反应所需的浓度(50~60 ng/μL),在-20 ℃冰箱中保存备用。
1.4 TRAP反应体系和PCR扩增产物分析
使用S1000TM Thermal Cycler(BioRad)PCR扩增仪进行扩增,反应体系为:10×buffer(Mg2+)5.0 μL,dNTPs(10 mmol/L each)4.0 μL,固定引物1.0 μL,随机引物1.0 μL,基因组 DNA(50~60 ng/μL)1.0 μL,Easy-Taq DNA Polymerase 0.5 μL,dd H2O补至总体积为50 μL;TRAP反应的PCR扩增程序为94 ℃预变性4 min;94 ℃变性45 s,35 ℃退火45 s,72 ℃延伸1 min,5个循环;然后94 ℃变性1 min,52 ℃退火1 min,72 ℃延伸1 min,35个循环;最后72 ℃延伸7 min,慢慢冷却至10 ℃。PCR扩增产物在0.5%非变性聚丙烯酰胺凝胶(PAGE)上电泳,105 V电泳4 h,银染,通过凝胶成像系统观察并拍照。
2 结果与分析
2.1 TRAP标记的多态性分析
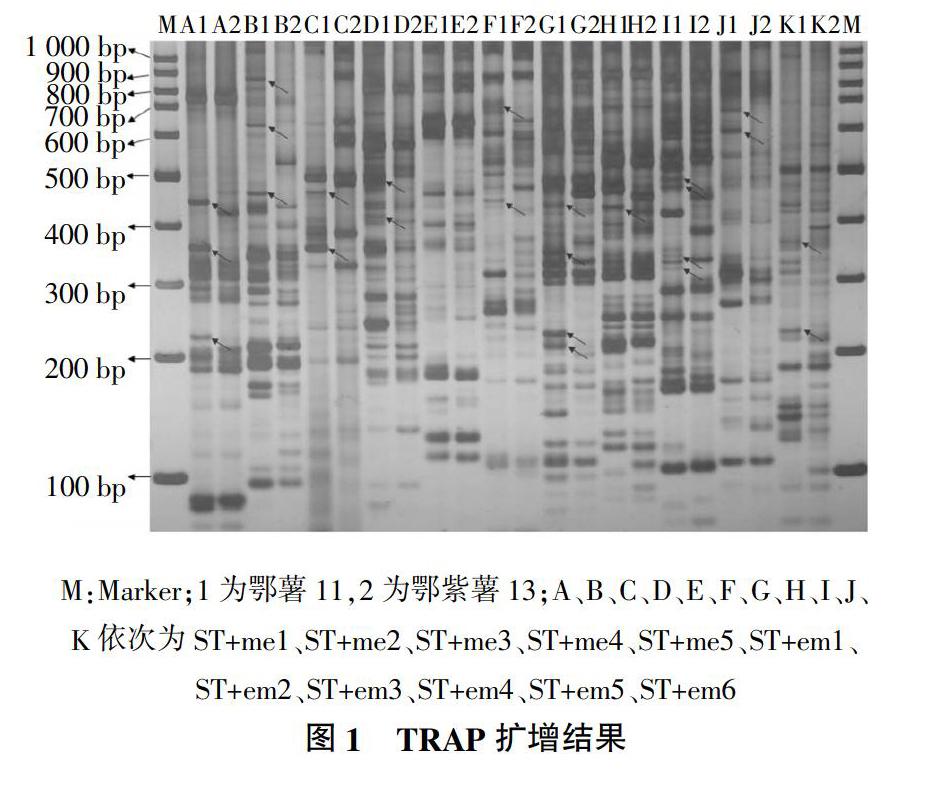
使用11对TRAP引物对2个试验材料进行分析,发现11对引物扩增片段主要集中在100~900 bp范围内,每对引物组合可产生8~20条清晰、稳定条带,不同TRAP引物组合扩增出的带型、条带数量和分布均匀程度具有较大差异。11对引物组合均表现出多态性,产生的多态性条带为4~10条,其中ST/me1、ST/me2、ST/em2、ST/em4等4对引物组合的多态性更为良好,可用于进一步分析研究(图1)。
2.2 SGT1基因的TRAP分析
对11对引物组合扩增出的多态性条带进行分析,主要集中在200~700 bp。鄂薯11与鄂紫薯13的抗性存在差异,发现某些条带在鄂薯11中出现,而在鄂紫薯13中没有产生条带。如图1箭头所指,组合ST/me1在鄂薯11中扩增出3条特异性条带,鄂紫薯13中没有出现;引物组合ST/em2在鄂薯11中扩增出4条特异性条带,而鄂紫薯13中没有出现这4条特异性条带。在这些多态性条带中,可能存在SGT1基因的特异扩增条带,这些片段可能与SGT1基因紧密相关。接下来可以对这些特异性片段进行回收、测序,做进一步的研究。
3 小结与讨论
通过基因克隆,发现SGT1基因在植物抗病中起到重要作用。李为民等[19]发现GbSGT1基因在海岛棉的抗黄萎病中发挥着重要作用。当经过黄萎病菌诱导后,GbSGT1基因的转录水平表达量显著提高。王凯等[20]研究发现,小麦体内SGT1基因的超量表达可以提高对黄矮病、白粉病的抗性。Xing等[21]将来自簇毛麦的SGT1基因导入普通小麦,发现对白粉病的抗性增强。蒋明等[22]以青花菜为试验材料,发现霜霉菌和核盘菌诱导BoSGT1的过量表达,表明BoSGT1基因对霜霉菌和核盘菌具有抗性。Uppalapati等[23]发现,在抑制SGT1表达的情况下,在受丁香假单胞菌侵染時,番茄和拟南芥叶片出现失绿和细胞死亡等病症。这些研究揭示了SGT1基因的多种抗病反应机制,为植物抗病基因工程的应用提供了更多的选择。
TRAP技术的固定引物是基于EST序列设计的,是表达基因的一部分,因此通过开发与表型或目标基因连锁的TRAP标记,被有效地应用于特定的基因定位。Miklas等[24]通过设计大豆的TRAP固定引物,在106个TRAP标记中有17个定位了与R基因相邻的基因,2个调节灰茎枯萎病抗性,1个具有大豆花叶病毒抗性和共同的细菌抗性,证明TRAP标记具有标记抗病性基因的潜力。Saleh等[25]通过构建小麦Yecora Rojo和Pavon 76的F4群体,发现3个与叶绿素含量性状相关联的标记,4个与叶片衰老相关联的标记和3个与叶片细胞膜稳定性相关联的标记,表明TRAP标记可以用于小麦的抗干旱育种。Chen等[26]结合SSR、SRAP和TRAP标记,发现了3个与小麦抗条锈病相关联的标记,证实YrSph可能是一个新型的小麦抗条锈病基因。张娜等[27]将TRAP技术在小麦抗叶锈病基因中Lr24的分子标记中应用成功,得到1个与Lr24紧密连锁的TRAP标记,可用于筛选抗叶锈病基因的群体。
甘薯小象甲是甘薯的一类重要病害,能造成甘薯的大面积减产[28]。研究发现SGT1和HSP90共同介导参与根线虫、马铃薯蚜虫和甘薯白蝇的抗性[29],据此推测,SGT1基因可能也参与甘薯抗小象甲的调控。TRAP标记在种质资源的鉴定评价、遗传图谱构建、重要性状基因标记、图位克隆、cDNA与gDNA指纹分析等方面具有重要的应用价值[30,31]。本研究利用TRAP技术对鄂薯11和鄂紫薯13进行检测,发现某些条带在鄂薯11中出现,而在鄂紫薯13中无法检测到,这些条带可能与甘薯抗病和抗虫基因有着紧密联系,表明TRAP标记可能在甘薯的抗病和抗虫遗传改良中有着应用价值。
参考文献:
[1] 王红意,翟 红,王玉萍,等.30个中国甘薯主栽品种的RAPD指纹图谱构建及遗传变异分析[J].分子植物育种,2009,7(5):879-884.
[2] 李 强,刘庆昌,马代夫.甘薯近缘野生种研究利用现状及展望[J].分子植物育种,2006,4(6):105-110.
[3] KITAGAWA K,SKOWYRA D,ELLEDGE S J,et al. SGT1 encodes an essential component of the yeast kinetochore assembly pathway and a novel subunit of the SCF ubiquitin ligase complex[J].Molecular cell,1999,4(1):21-33.
[4] 王 凯,张增艳,黄 璜,等.小麦SGT1基因的克隆与表达特性分析[J].麦类作物学报,2007,27(6):952-956.
[5] SEO Y S,LEE S K,SONG M Y,et al. The HSP90-SGT1-RAR1 molecular chaperone complex:A core modulator in plant immunity[J].Journal of plant biology,2008,51(1): 1-10.
[6] KADOTA Y,SHIRASU K,GUEROIS R. NLR sensors meet at the SGT1-HSP90 crossroad[J].Trends in biochemical sciences,2010,35(4):199-207.
[7] AUSTIN M J,MUSKETT P,KAHN K,et al. Regulatory role of SGT1 in early R gene-mediated plant defenses[J].Science,2002,295(5562):2077-2080.
[8] AZEVEDO C,SADANANDOM A,KITAGAWA K,et al. The RAR1 interactor SGT1,an essential component of R gene-triggered disease resistance[J].Science,2002,295(5562):2073-2076.
[9] LEISTER R T,DAHLBECK D,DAY B,et al. Molecular genetic evidence for the role of SGT1 in the intramolecular complementation of Bs2 protein activity in Nicotiana benthamiana[J].The plant cell,2005,17(4):1268-1278.
[10] DECROOCQ V,SICARD O,ALAMILLO J M,et al. Multiple resistance traits control Plum pox virus infection in Arabidopsis thaliana[J].Molecular plant-microbe interactions,2006,19(5):541-549.
[11] HU J,VICK B A. Target region amplification polymorphism:A novel marker technique for plant genotyping[J].Plant molecular breeding reporter,2003,21(3):289-294.
[12] SUN S J,GAO W,LIN S Q,et al. Analysis of genetic diversity in Ganoderma population with a novel molecular marker SRAP[J].Applied microbiology and biotechnology,2006,72(3):537-543.
[13] WRIGHT E M,AWALE H E,KELLY J D. Use of TRAP markers to map resistance to a new race of common bean rust in Michigan[J].Annual report-bean improvement cooperative,2008,51:210.
[14] MENZO V,GIANCASPRO A,GIOVE S,et al. TRAP molecular markers as a system for saturation of the genetic map of durum wheat[J].Euphytica,2013,194(2):151-160.
[15] 王一伊,向秋燕,閻文昭,等.利用TRAP标记分析83份烟草种质资源遗传多样性及遗传演化关系[J].西南农业学报,2016, 29(3):503-510.
[16] YU J,YU S,LU C,et al. High-density linkage map of cultivated allotetraploid cotton based on SSR,TRAP,SRAP and AFLP markers[J].Journal of integrative plant biology,2007,49(5):716-724.
[17] LI G,QUIROS C F. Sequence related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction:Its application to mapping and gene tagging in Brassica[J].Theoretical and applied genetics,2001,103:455-461.
[18] 张世安,邢智峰,刘永英,等.苔藓植物DNA不同提取方法的比较分析[J].河南科学,2009,27(5):559-562.
[19] 李为民,王志兴,张永强,等.海岛棉GbRar1和GbSgt1基因的过量表达提高烟草离体叶片对赤星病的抗性[J].自然科学进展,2004,14(4):410-416.
[20] 王 凯,杜丽璞,张增艳,等.中间偃麦草SGT1基因的克隆及其抗病功能的分析[J].作物学报,2008,34(3):520-525.
[21] XING L,QIAN C,CAO A,et al. The Hv-SGT1 gene from Haynaldia villosa contributes to resistances towards both biotrophic and hemi-biotrophic pathogens in common wheat (Triticum aestivum L.)[J].PLoS one,2013,8(9):e72571.
[22] 蒋 明,张志仙,潘小翠,等.青花菜抗病防卫基因BoSGT1的克隆、序列分析与诱导表达[J].浙江大学学报(理学版),2015, 42(4):453-458.
[23] UPPALAPATI S R,ISHIGA Y,RYU C M,et al. SGT1 contributes to coronatine signaling and Pseudomonas syringae pv. tomato disease symptom development in tomato and Arabidopsis[J].New phytologist,2011,189(1):83-93.
[24] MIKLAS P N,HU J,GR?譈NWALD N J,et al. Potential application of TRAP (targeted region amplified polymorphism) markers for mapping and tagging disease resistance traits in common bean[J].Crop science,2006,46(2):910-916.
[25] SALEH M S,AL-DOSS A A,ELSHAFEI A A,et al. Identification of new TRAP markers linked to chlorophyll content,leaf senescence,and cell membrane stability in water-stressed wheat[J].Biologia plantarum,2014,58(1):64-70.
[26] CHEN S S,CHEN G Y,CHEN H,et al. Mapping stripe rust resistance gene YrSph derived from Tritium sphaerococcum Perc. with SSR,SRAP,and TRAP markers[J].Euphytica,2012, 185(1):19-26.
[27] 张 娜,冀紅柳,杨文香,等.小麦抗叶锈病基因Lr24的TRAP分析[J].中国农业科学,2009,42(5):1841-1848.
[28] 叶明鑫.甘薯小象甲发生特点调查与原因分析[J].中国农学通报,2015,31(4):195-199.
[29] BHATTARAI K K,LI Q,LIU Y,et al. The Mi-1-mediated pest resistance requires HSP90 and SGT1[J].Plant physiology,2007,144(1):312-323.
[30] YUE B,VICK B A,CAI X,et al. Genetic mapping for the Rf1 (fertility restoration) gene in sunflower(Helianthus annuus L.) by SSR and TRAP markers[J].Plant breeding,2010,129(1):24-28.
[31] MENZO V,GIANCASPRO A,GIOVE S,et al. TRAP molecular markers as a system for saturation of the genetic map of durum wheat[J].Euphytica,2013,194(2):151-160.
