川西北亚高山紫果云杉和岷江冷杉异龄叶性状及养分再吸收
2019-11-02王玉荧鲜骏仁曾瑞琪朱莹莹胡建成陈国鹏
王玉荧,鲜骏仁*,曾瑞琪,朱莹莹,胡建成,陈国鹏,马 俊
(1.四川农业大学环境学院,成都 611130;2.甘肃农业大学林学院,兰州 730070)
养分再吸收是植物将养分从衰老叶片转移到其他部位的过程[1],普遍存在于多年生植物中[2],是养分不足地区的陆生植物[3]保持和提升净初级生产的重要策略[4]之一。近年来,我国对植物叶片养分再吸收的研究已陆续展开,研究区域从东南海岸带[5-6]到青藏高原[7]、从南方的长江流域[8]和红壤侵蚀区[9]到北方草地[10];这些涉及森林、草地、经济植物和园林绿化植物的研究表明:养分再吸收与林龄[7]、元素类别[11]和植物种类[12]等因素有关,亦受土壤肥力[13]、降水量和温度[14]以及群落演替[15]等外部环境影响。但也有研究得出不一致的结论,如气候基本不影响瑞典北部高山湿地植物的叶片养分再吸收[16]。因此,现有研究尚不足以揭示陆地生态系统的植物再吸收特征。
川西亚高山地区位于青藏高原东南缘,是我国第二大林区和长江上游生态屏障的主体[17],良好的营养物质循环是其森林发挥多重效益的重要基础。前人在此就凋落物分解[18]、土壤性质[19]、森林碳功能[20]、物种多样性[21]以及水文[22]等方面进行了大量研究,但植物养分再吸收的研究鲜见报道。川西北亚高山原始林中紫果云杉(Picea purpurea)和岷江冷杉(Abies faxoniana)的叶片以异龄形式存在,这是否会导致叶片性状和养分含量的差异、进而影响叶片养分再吸收效率? 基于此,本研究以川西北亚高山原始针叶林建群种——紫果云杉和岷江冷杉为对象,测定异龄叶和凋落叶的性状和养分,阐明这2种植物的养分再吸收特征,以期为川西北原始针叶林的养分管理提供基础数据和科学依据。
1 材料和方法
1.1 研究区概况
本研究位于四川省平武县王朗国家级自然保护区(32°59'N,104°01' E,海拔 2 300~4 980 m),地处青藏高原东缘,属山地季风性气候,干季(11月—翌年4月)日照强、降水少、气候寒冷、空气干燥,湿季(5—10月)降雨集中、多云雾、日照少。年均气温2.5~2.9 ℃,绝对最高和绝对最低温为26.2 ℃和-17.8 ℃,年降雨量约801~825 mm;该地区山地植被垂直分布明显,海拔2 600 m 以下为针阔叶混交林带,2 600~3 800 m 分布有大量的原始针叶林;主要植被为亚高山针叶林,主要土壤类型包括棕色冲积土、棕壤、暗棕壤、亚高山草甸土等[23]。
1.2 样品采集与前处理
在2017年秋叶片凋落季,于王朗国家级自然保护区海拔为2 800~3 000 m 的原始针叶林中,随机采集森林中间成熟植株上部的当年生(1 a)和次年生(2 a)的紫果云杉和岷江冷杉叶片(3 a 及多年生的紫果云杉和岷江冷杉叶片仅存于幼苗幼树),紫果云杉及岷江冷杉枝条末端当年生小枝上的叶片为1 a 叶,紧靠当年生、芽鳞隔开的小枝上的叶片为2 a 叶[24];凋落叶为从林下收集的刚凋落、发黄但未腐坏分解的叶片(野外调查时发现成熟植株的1 a及2 a 叶少有凋落,凋落叶多为3 a 叶)。采集的正常叶片来自5 个采集点的至少15 个植株,分别装入自封袋,记录编号后迅速带回实验室。
1.3 样品测定
在每组样品中随机选取30 个叶片用数显卡尺(±0.01 mm)测量叶长、叶宽、叶厚,采用叶面积仪(CI-203,CID,USA)测定单叶面积。将测量完叶片性状的样品放入烘箱,105 ℃杀青20 min 后于70 ℃烘至恒量,测定完单叶干质量后粉碎过筛用于测定N、P 和 K 含量。叶片 N、P 和 K 含量分别采用浓H2SO4-H2O2消煮-凯氏定氮法(NY/T 2017-2011)、酸溶-钼锑抗比色法(NY/T 2017-2011)和火焰光度法(NY/T 2017-2011)测定,所有测定均重复3 次。
1.4 数据处理
比叶面积(specific leaf area,SLA)用 SLA=s/m(s指叶面积,m 指叶片干质量)计算[25],养分再吸收效率(resorption efficiency,RE)采用 RE(%)=(1-A2/A1)×100%(A1和A2为异龄叶和凋落叶的元素含量)[26]计算。利用箱图来展现叶片性状;采用双因素显著性方差和多变量因果模型检验物种、叶龄对N、P、K及其计量的影响;采用Spearman 相关系数分析叶片性状与营养元素及其生态计量之间的相关性。所有分析在 SPSS 20.0(SPSS Inc.,Chicago,IL,USA)上进行,采用 Origin 8.0(Origin Lab,MA)作图。
2 结果与分析
2.1 叶片性状
3 种紫果云杉叶的长度有极显著差异(表1,P<0.01):凋落叶(6.10~11.00 mm)>2 a 叶(4.50~7.00 mm)>1 a 叶(4.10~6.50 mm);1 a 叶叶长分布更汇聚、凋落叶叶长分布更分散(图1Ⅰ)。1 a 叶叶宽(1.05~2.20 mm)和 2 a 叶(1.11~2.13 mm)无显著差异(P=0.973),均极显著大于凋落叶(0.85~1.80 mm;表1,P<0.01);凋落叶叶宽分布更汇聚、1 a 叶叶宽分布更分散(图1Ⅲ)。1 a 叶叶厚(0.54~1.07 mm)显著小于2 a 叶(0.62~1.21 mm,P=0.017),两者均极显著(表1,P<0.01)大于凋落叶(0.42~0.83 mm);凋落叶厚度分布更汇聚、2 a 叶厚度分布更分散(图1Ⅴ)。2 a 叶的SLA(36.67~70.00 cm2/g)与凋落叶(27.27~57.14cm2/g)无显著差异(P=0.926),均极显著小于 1 a 叶(26.67~118.18 cm2/g;表1,P<0.01);凋落叶的 SLA 分布更汇聚、1 a 叶的SLA 分布更分散 (图1Ⅶ)。
岷江冷杉凋落叶叶长(15.70~26.50 mm)极显著大于 1 a 叶(7.20~16.80 mm)和 2 a 叶(10.10~19.10 mm;表1,P<0.01),后两者之间无显著差异(P=0.155);2 a 叶叶长分布更汇聚、凋落叶叶长分布更分散(图1Ⅱ)。1 a 叶叶宽(1.10~2.55 mm)和凋落叶(1.49~2.49 mm)无显著差异(表1,P=0.942),均极显著小于 2 a 叶(1.98~2.65 mm,P<0.01);2 a 叶叶宽分布更汇聚、1 a 叶叶宽分布更分散(图1Ⅳ)。1 a 叶厚度(0.34~0.78 mm)与凋落叶(0.40~0.76 mm)无显著差异(P=0.649),都极显著小于 2 a 叶(0.51~0.95 mm;表1,P<0.01);凋落叶厚度分布更汇聚、1 a 叶及 2 a叶厚度分布更分散(图1Ⅵ)。2 a 叶的 SLA(40.00~65.91 cm2/g)和凋落叶(32.97~75.00 cm2/g)无显著差异(P=0.587),都极显著小于 1 a 叶(26.09~107.14 cm2/g;P<0.01);2 a 叶的 SLA 分布最汇聚、1 a 叶最分散(图1Ⅷ)。
2.2 叶片养分含量及生态计量
紫果云杉3 种叶片之间的N、P 和K 含量有极显著差异(P<0.01),均表现为 1 a 叶>2 a 叶>凋落叶(表2)。紫果云杉 2 a 叶的 N/P 略高于凋落叶(P=0.070),均极显著高于 1 a 叶(P<0.01,图2Ⅰ)。3 种叶片之间的 N/K 和 K/P 均有极显著差异(P<0.01),N/K 表现为 1 a 叶<2 a 叶<凋落叶(图2Ⅱ),而 K/P则为凋落叶<2 a 叶<1 a 叶(图2Ⅲ)。
岷江冷杉3 种叶片的N、P 和K 含量有极显著差异(P<0.01,表2),P、K 表现为:1 a 叶>2 a 叶>凋落叶。岷江冷杉 3 种叶片之间的 N/P、N/K 和 K/P 均有极显著差异(P<0.01):N/P 和 N/K 均表现为 1 a 叶<2 a 叶<凋落叶(图2Ⅰ、Ⅱ);而 K/P 则表现为凋落叶<2 a 叶<1 a 叶(图2Ⅲ)。
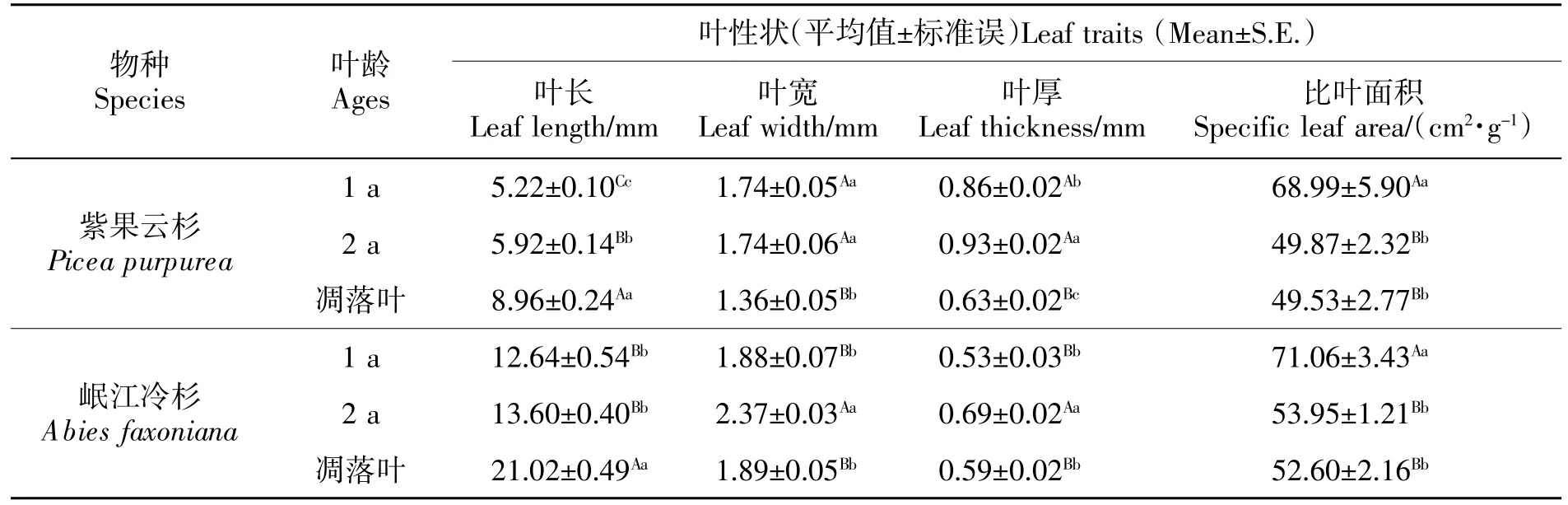
表1 叶片性状特征Table 1 The characteristics of leaf traits

图1 叶片性状变化(n=30)Figure 1 Changes of leaf traits(n=30)

表2 叶片养分含量(n=3)Table 2 Nutrient concentrations in leaves(n=3) mg·g-1

表3 物种和叶龄对叶片元素含量及生态计量影响的双因素分析表Table 3 Results(P-value)of two-way ANOVAs on the effects of species,age and their interactions on the leaf nutrient concentrations and ecological stoichiometric ratios
双因素显著分析表明(表3):除物种×叶龄显著影响N/P 值,物种、叶龄以及两者交互效应都极显著影响叶片中的 N、P、K 含量及其计量(P<0.01)。

图2 营养元素的生态计量Figure 2 Ecological stoichiometries between nutrient elements of different species
2.3 元素再吸收效率
本文以1 a 叶、2 a 叶以及两者的平均值(以下简称“平均值”)分别计算了紫果云杉和岷江冷杉叶片的N、P、K 再吸收效率(表4),结果表明除岷江冷杉 N 值外,均分别表现为 2 a 叶<平均值<1 a 叶,紫果云杉和岷江冷杉对叶片N 再吸收效率无显著差异,对叶片P、K 再吸收效率存在极显著差异性,这说明采用不同的再吸收效率计算基准将导致巨大的差异。此外,本文还以2 a 叶为基础值计算1 a 叶和2 a 叶之间的再吸收效率,结果表明紫果云杉1 a叶对 N、P、K 有明显的再吸收,吸收率分别为5.31%、29.77%和 34.29%;岷江冷杉仅对 P、K 有明显的再吸收,吸收率分别为12.17%和32.11%,2 a叶中N 的含量较高,可能是2 a 叶仍有较强的光合能力。双因素显著性分析结果(表5)表明除叶龄和物种×叶龄对N 再吸收效率影响不显著,其他因素对叶片N、P、K 的再吸收效率均有极显著影响。
2.4 叶性状与营养元素及再吸收效率之间相关性
紫果云杉叶长与N、P、K 含量以及N/P 比呈负相关,与 N/K、K/P 比以及 N、P、K 再吸收效率呈正相关,均不显著;叶宽与 K/P 比以及 N、P、K 再吸收效率呈负相关,与 N、P、K 含量、N/P、N/K 比呈正相关,均不显著;叶厚与 N、P 含量、N/P 比以及 N/K 比呈正相关,与 K 含量、K/P 比以及 N、P、K 再吸收效率呈负相关,均不显著;比叶面积与N、P、K 含量以及K/P比呈显著正相关,与P 再吸收效率呈显著负相关,与N/K 比以及K 再吸收效率呈极显著负相关(表6)。岷江冷杉叶长与N/P、N/K 比以及P 再吸收效率呈负相关,与 N、P、K 含量、K/P 比以及 N、K 再吸收效率呈正相关,均不显著;叶宽与N、P、K 含量以及N再吸收效率呈负相关,与 N/P、N/K、K/P 以及 P、K 再吸收效率呈正相关,均不显著;叶厚与P、K 含量以及 K/P 比呈负相关,与 N 含量、N/P、N/K 比以及 N、P、K 再吸收效率呈正相关,均不显著;比叶面积与N、K 含量、K/P 比以及 P、K 再吸收效率呈负相关,与 P 含量、N/P、N/K 比以及 N 再吸收效率呈正相关,均不显著(表6)。
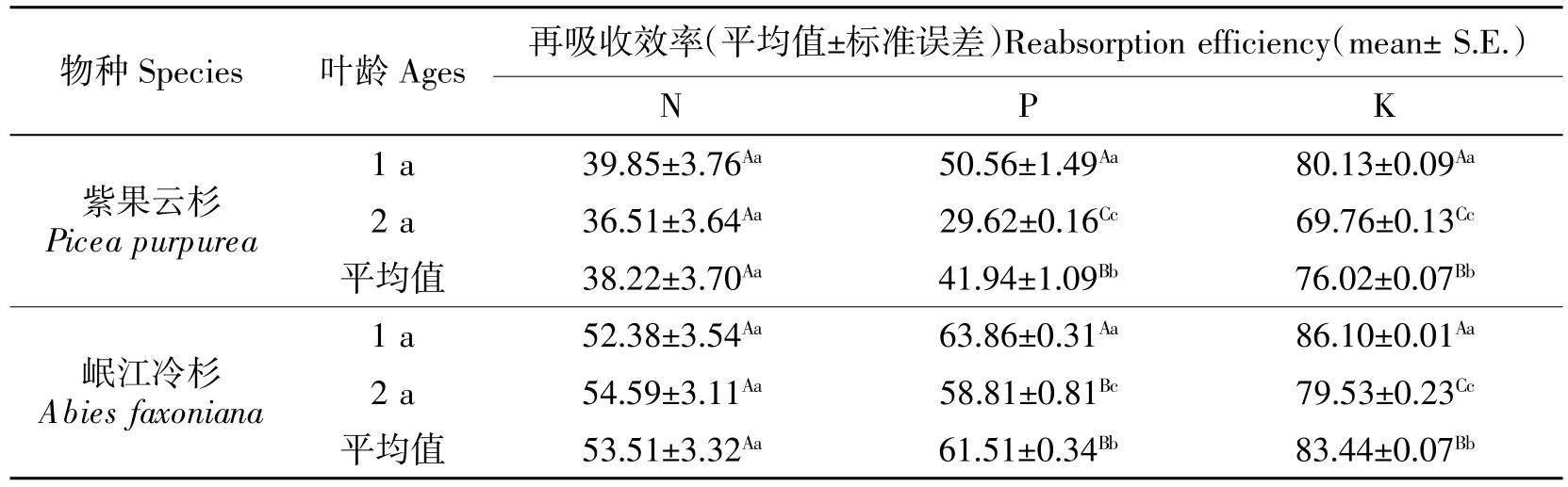
表4 养分再吸收效率(n=3)Table 4 Nutrient reabsorption efficiency of different species(n=3) %

表5 物种和叶龄对养分再吸收效率影响的双因素分析表Table 5 Results(P-value)of two-way factorial ANOVA on the effects of species,age and their interactions on the leaf nutrient reabsorption efficiency

表6 叶性状与营养元素及其再吸收效率之间的相关关系Table 6 Correlation between the leaf traits and the nutrient elements
3 讨论
3.1 叶片养分含量及其再吸收效率
本研究中,叶片的N、P 含量均高于岳桦(Betula ermanii)和牛皮杜鹃(Rhododendron aureum)[27]、湿地松(Pinus elliottii)[28]和马尾松(Pinus massoniana)[29],低于桂南喀斯特植被[30],远低于杉木(Cunninghamia lanceolata)[31],与宁秋蕊等[1]对国内文献综述的平均结果相当。与Yuan Z.Y.等[32]的结果不一致,后者研究中白云杉(Picea glauca)叶N、P 含量高于香脂冷杉 (Abies balsamea)。异龄叶的K 含量远低于湿地松[28]和马尾松[9];但凋落叶的K 含量则高于湿地松[28]而低于马尾松[9]。这表明叶片的养分含量受生境、树种和元素种类的影响。
本研究还表明,从 1 a 叶、2 a 叶到凋落叶,P、K养分含量逐渐下降,这与宁秋蕊等[9]对马尾松、Yuan Z.Y.[32]对白云杉和香脂冷杉的研究结果相一致;基于生长季节内叶片生长过程中养分含量的研究也表明,春季新生叶养分含量远高于秋季成熟叶[33],其原因是新生叶生长旺盛、代谢较快,需要更多物质用于生长,故养分含量较高[32]。成熟叶/老龄叶保存有一定量的养分,也可能是常绿植物的一种适应方式,即通过延长叶片寿命,既可以将叶片作为养分的储存器官以加速养分的周转[34],也可在一定程度加强光合以合成更多的有机物供植物生长利用[31]。
本研究中,两种植物养分再吸收效率总体趋势与我国亚热带南方红壤侵蚀区马尾松[9]和湿地松的养分再吸收效率[28]基本一致,即N<P<K。本研究中紫果云杉的叶片N、P 再吸收均低于全球平均的44.8%和53.5%、而岷江冷杉均高于全球平均水平[14],这表明叶片养分再吸收效率的大小有明显的物种差异。紫果云杉叶片的N、P、K 再吸收的Spearman相关性整体表现为极显著正相关(rN-P=0.769、P=0.003,rN-K=0.813、P=0.001,rP-K=0.907、P<0.001),而岷江冷杉叶片仅有P、K 再吸收表现为极显著正相关(r=0.981、P<0.001),这也进一步表明叶片养分再吸收有显著的物种差异。另外,凋落物养分含量越低,植物对其回收利用程度越高,这与K.T.Killing beck[35]的结果一致。
植物叶片养分的生态计量(如N/P)的临界比值可以判断环境对植物生长的养分供应状况的指标[36]。W.Koerselman[37]通过对欧洲施肥实验分析得出“植物鲜叶 N/P 小于 14,受到 N 的限制;N/P 大于16,受到 P 的限制;当 N/P 介于 14~16 之间,受到N、P 两种元素的限制”。这个结论被广泛应用于生态系统养分限制因子的判断。本研究中,紫果云杉异龄叶的N/P 小于岷江冷杉,后者的N/P 更接近于14~16 之间,表明紫果云杉受N 的限制不如岷江冷杉、且岷江冷杉还受P 的限制;基本不同计算方式的再吸收效率都表明岷江冷杉对养分的需求远高于紫果云杉。H.O.Venterink 等[38]对湿地的综述也指出,当 N/P<14.5、N/K>2.1,则 N 受限;当 N/P>14.5、K/P>3.4,则 P 或 P+N 受限;当 N/K>2.1、K/P<3.4,则 K 或 K+N 受限。参照此标准,则本研究中紫果云杉和岷江冷杉均受到N 限制。但也有研究表明,叶片养分再吸收与土壤养分有效性关系不大[9,30]。进一步对本研究中养分再吸收效率及其与生态计量的关系进行Spearman 相关分析,结果表明:紫果云杉仅N 的再吸收与凋落叶的N/P 呈极显著负相关(r=-0.843,P=0.001);而岷江冷杉则表现为N 再吸收与异龄叶中除N/P 的生态计量呈显著正相关(rN/P=0.503、P=0.096,rN/K=0.650、P=0.022,rP/K=0.699、P=0.011),N 再吸收与凋落叶的 P/K 呈极显著正相关(r=0.979,P<0.001),P 和 K 再吸收仅与凋落叶的 N/K 呈显著正相关(rP=0.629、P=0.029,rK=0.641、P=0.025)。因此,叶片养分再吸收与生态计量的关系复杂需要更多的研究进一步揭示。
3.2 异龄叶性状差异对营养元素及其再吸收效率的影响
采用Spearman 相关系数对两种植物叶片的长、宽、厚和SLA 与N、P、K 及其生态计量、再吸收效率进行了分析,结果表明两种植物的叶长、叶宽以及叶厚与其营养元素含量、生态计量及其再吸收效率之间没有显著相关性;SLA 与紫果云杉的N、P、K 含量和K/P 显著正相关,与P 再吸收效率显著负相关,与 K 和 N/K 呈极显著负相关(P<0.01);而与岷江冷杉的养分含量及其生态计量无显著相关。这说明叶性状对植物叶片养分的影响与植物种类和所选取的指标有关。实际上,气候变化(从年际到长期)都会对常绿植物异龄叶的性状[24]和小枝(twig)性状[39]产生重要影响,小枝的结构特征及其与叶性状之间的关系决定着植物对光照、空间等资源的利用和适应策略[40]。因此,叶性状与养再吸收的关系可能需要在叶片-小枝-植株的不同尺度进行揭示。比如长期升温没有增加细穗柳(Salix eriostachya)单叶面积,但增加了整个植株的叶面积和生物量[41]。
4 结论
本研究证明在川西北亚高山原始针叶林中,建群植物紫果云杉和岷江冷杉叶片性状和养分含量在异龄叶和凋落叶之间存在极大差异,并且异龄叶之间性状特征及养分含量存在差异,仅比叶面积对紫果云杉的养分再吸收效率有显著性影响。从叶养分的生态计量和养分再吸收可以初步判断紫果云杉和岷江冷杉的生长均受N 不足的影响,叶片的养分再吸收是高寒地段植物适应养分贫瘠的一种重要策略,但紫果云杉的再吸收能力差于岷江冷杉,因此,在将来的养分管理中,紫果云杉应受到更多关注。
