三七叶总皂苷抗老年焦虑症的作用及机制研究
2019-10-30杜瑶李欣张桂仙袁娅金周宁娜
杜瑶 李欣 张桂仙 袁娅金 周宁娜

[摘要] 目的 探討三七叶总皂苷(tSPNL)抗老年焦虑症的作用及其作用机制。 方法 12月龄雄性小鼠75只按照随机数字表法分为群养(n = 15,正常对照组)和孤养(n = 60),4周后孤养动物随机分为模型组(溶媒)、地西泮(BDZ)组、tSPNL高剂量组(500 mg/kg tSPNL)及tSPNL低剂量组(100 mg/kg tSPNL),每天灌胃给药1次,连续给药14 d后用高架十字迷宫(EPM)进行行为学测定;行为学实验结束后,采用酶联免疫吸附法检测海马内γ-氨基丁酸(GABA)、脑源性神经营养因子(BDNF)的含量。另取12月龄雄性小鼠135只孤养,4周后随机分为模型组(溶媒)、BDZ组、氟西汀组、tSPNL高剂量组,每天灌胃给药1次,连续灌胃给药14 d后脑室内注射BDNF受体TrkB阻断剂K252a、GABAA受体阻断剂荷包牡丹碱(Bic),1 h后EPM进行行为学检测。 结果 与模型组比较,tSPNL高剂量组小鼠开臂时间百分比(OT%)增加,差异有高度统计学意义(P < 0.01);海马BDNF、GABA水平增加,差异有统计学意义(P < 0.05或P < 0.01)。tSPNL组脑室内同时注射K252a及Bic后OT%降低,差异有统计学意义(P < 0.05)。 结论 tSPNL具有抗老年焦虑症的作用,其抗焦虑作用与BDNF-GABA信号通路有关,可能通过增加BDNF水平改善GABA能神经的可塑性,从而降低老龄小鼠焦虑易感性。
[关键词] 老年焦虑症;三七叶总皂苷;BDNF-GABA信号通路;γ-氨基丁酸能神经可塑性
[中图分类号] R284.2 [文献标识码] A [文章编号] 1673-7210(2019)08(a)-0007-04
[Abstract] Objective To explore the effect and mechanism of total saponins of Panax notoginseng leaves (tSPNL) on late-life anxiety. Methods Fifty aging male mice (12 months) were reared in group (n = 15, normal control group) or isolation (n = 60) according to the random number table method. After 4 weeks, the isolated rearing animal were then randomly divided into model group (vehicle), Diazepam (BDZ) group, tSPNL 500 group (500 mg/kg tSPNL) and tSPNL 100 group (100 mg/kg tSPNL). All mice were intragastrically administered desired compounds once a day for consecutive 14 days. The anxiety-like behaviors were evaluated by elevated plus maze (EPM), and levels of GABA and BDNF in hippocampus were determined by enzyme linked immunosorbent assay after the behavior test. In another set, aging male mice (n = 135) were rearing isolated for 4 weeks and were then randomly divided into model (vehicle), BDZ group, fluoxetine group, tSPNL 500 group. After administration of desired compounds intragastrically once a day for 14 days, animals underwent intraventricular injection of either BDNF receptor TrkB antagonist K252a or GABAA receptor antagonist Bicuculline (Bic), and the anxiety-like behaviors were measured by EPM 1 hour later. Results Compared with model group, the percentage of open arm time (OT%) increased significantly in tSPNL 500 group (P < 0.01), and the levels of BDNF and GABA also increased in hippocampus significantly (P < 0.05 or P < 0.01). After intraventricular injection of K252a and Bic, OT% in tSPNL 500 group decreased significantly (P < 0.05). Conclusion tSPNL can alleviate the anxiety-like behaviors in aging mice, and the mechanism is involved in BDNF-GABA signaling pathway, which may restore the GABAergic plasticity by increasing BDNF levels, thereby reducing the anxiety susceptibility of aging mice.
[Key words] Late-life anxiety; Total saponins of Panax notoginseng leaves; GABA-BDNF signaling pathway; GABAergic plasticity
焦虑症是一种以焦虑情绪为主的神经症,在老年人群中的发病率逐年上升[1],预计到2050年全球患焦虑症的老年人数量将达到20亿[2]。老年焦虑症发病机制与一般焦虑症不同,与其神经退行性病变有关。γ-氨基丁酸(γ-aminobutyricacid,GABA)是中枢神经系统中调节精神活动的主要神经递质,其中GABA A受体(GABAA-R)是产生抗焦虑效应的主要功能受体[3-4]。脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)是GABA能神经在海马区的效应分子,对GABA能神经的可塑性起着关键作用。GABAA-R可激活BDNF-TrkB-ERK1/2信号通路,触发BDNF释放,上调BDNF表达水平;反之BDNF通过突触前TrkB受体也可促进GABA的释放并增加细胞面表GABAA-R的表达,参与情绪调节[5-7]。老年人脑内BDNF水平下降,导致GABA能神经可塑性障碍,焦虑易感性增加,认知功能受损[8]。因此通过增加脑内BDNF水平恢复GABA能神经可塑性可能是老年焦虑症治疗的新策略。
研究发现三七叶总皂苷(total saponins of Panaxnotoginseng leaves,tSPNL)可上调BDNF及其受体TrkB的表达而发挥抗抑郁作用[9-10]。因此,我们推测tSPNL可能通过增加BDNF水平改善GABA能神经可塑性治疗老年焦虑症。本研究主要通过观察tSPNL对老年焦虑小鼠行为学的影响,探讨其与BDNF-GABA信号通路相关的作用机制,为其进一步的实验研究提供理论基础。
1 材料与方法
1.1 动物
昆明种小鼠,SPF级,雄性,体重40~50 g,12月龄,由四川省医学科学院实验动物研究所提供,合格证号:0016699。本动物研究方案经云南中医药大学动物使用和护理委员会批准(许可证号:R-062014054)。
1.2 药物与试剂
三七叶皂苷(云南红云生物工程技术有限公司);地西泮(BDZ,江苏济川制药有限公司,批号:080411);氟西汀(FXT,礼来苏州制药有限公司);蛋白定量测定试剂盒(南京建成生物工程研究所,货号:A045-2);GABA酶联免疫吸附试验(ELISA)试剂盒(南京建成生物工程研究所,货号:H168-96T);BDNF ELISA试剂盒(南京建成生物工程研究所,货号:H069-96T);荷包牡丹碱(sigma公司,货号:2076197);K252a(abcam公司,货号:ab120419)。
1.3 仪器
小鼠高架十字迷宫,西班牙Panlab;SMARTUPG V2.5小动物行为学记录分析系统,西班牙Panlab;电子天平(精度0.1mg),余姚市金诺天平仪器有限公司;脑立体定位仪,深圳瑞沃德生命科技有限公司。
1.4 方法
75只12月龄雄性小鼠按照随机数字表方分成孤养和群养。群养15只,作为正常对照组;孤养小鼠60只在不透明笼内单笼单只隔离饲养复制社会隔离焦虑症模型,4周后随机分为模型组、BDZ组(BDZ 2 mg/kg)、tSPNL高剂量组(tSPNL 500 mg/kg)及tSPNL低剂量组(tSPNL 100 mg/kg),每组15只,模型组给予等剂量溶媒,每天灌胃給药1次,连续给药14 d,于末次给药后1 h进行小鼠行为学实验。
1.4.1 小鼠高架十字迷宫实验 参照文献[18]报道方法,第14天末次给药1 h后,进行小鼠高架十字迷宫实验。测试时将小鼠置于高架十字迷宫中央平台区域,分析系统记录分析5 min内动物进入开臂与闭臂的次数及在开臂与闭臂内的停留时间,计算进入开臂次数与总入臂次数的百分比(open arm entry,OE%)及在开臂内运动时间与开臂闭臂内的总时间的百分比(open arm time,OT%)。以OE%和OT%代表药物抗焦虑作用指标,总入臂数作为动物的运动活性指标,每只小鼠测5 min。
1.4.2 小鼠脑内GABA、BDNF水平的测定 行为学测试后1 d,颈椎脱臼处死,分离出海马,海马体重(g)与匀浆液(mL)=1∶9的比例在冰浴条件下机械匀浆,4℃离心15 min后,取上清液,考马斯亮兰法测定蛋白含量,调整待测样品蛋白浓度,按试剂盒说明书进行小鼠脑内GABA、BDNF含量测定。
1.4.3 脑室内注射受体阻断剂阻断BDNF-GABA信号通路 氟西汀可通过增加脑内BDNF水平发挥抗焦虑作用,而地西泮则主要通过作用于脑内GABA受体发挥作用,为探讨tSPNL抗焦虑作用是否通过BDNF-GABA信号通路,本研究选用FXT和BDZ进行对照。实验另取135只12月龄雄性小鼠参照“1.4”项复制社会隔离焦虑症模型,模型复制后将小鼠随机分为模型组(生理盐水,n = 15)、FXT组(FXT 10 mg/kg,n = 30)、BDZ组(BDZ 2 mg/kg,n = 30)和tSPNL 500 mg/kg(tSPNL 500,n = 60)组,模型组给予等剂量溶媒,连续灌胃给药14 d。将各给药组分为实验组(n = 15)和tSPNL对照组(n = 15),末次给药30 min后FXT组脑室注射K252a 0.5 μg/只(FXT+K252a),BDZ组脑室注射荷包牡丹碱(Bicuculline,Bic)0.5 μg/只(BDZ+Bic),tSPNL实验组分为脑室注射K252a 0.5 μg/只(tSPNL+K252a)或Bic 0.5 μg/只(tSPNL+Bic)或同时注射K252a和Bic 0.5 μg/只(tSPNL+K252a+Bic)。FXT、BDZ组、tSPNL组对照组和模型组脑室内注射等剂量生理盐水。注射方法:乙醚麻醉小鼠,将小鼠固定于手术台,碘伏消毒小鼠头部,剪开头皮暴露前囟和矢状缝,取前囟后0.25 mm,矢状缝1.0 mm位置,用颅钻将该位点头骨打磨薄,将微量注射器缓慢进入脑内,深度2.5~3.0 mm,注射持续15 s。注射完成后留针5 min,缓慢旋转出针,缝合伤口。脑室注射1 h后,进行行为学检测。
1.5 统计学方法
采用GraphPad Prism 5.0统计学软件进行数据分析,计量资料用均数±标准差(x±s)表示,非正态分布采用非参数检验,正态分布则进行方差齐性检验,方差齐采用One-way ANOVA Post-Hoc Tukey法进行多组间两两比较检验,方差不齐则采用秩和检验(Kruskal-Wallis检验);计数资料用率表示,组间比较采用χ2检验。以P < 0.05为差异有统计学意义。
2 结果
2.1 tSPNL对老龄小鼠行为学的影响
与正常对照组比较,模型组小鼠OE%和OT%明显减少,差异有统计学意义(P < 0.05或P < 0.01)。与模型组比较,BDZ组小鼠OE%、OT%均明显增加,差异有统计学意义(P < 0.05或P < 0.01);tSPNL 500组小鼠OT%增加,差异有高度统计学意义(P < 0.01);tSPNL 100组小鼠OE%、OT%均有升高的趋势,但差异无统计学意义(P > 0.05)。提示tSPNL具有抗焦虑作用,在500 mg/kg剂量下作用明显。见图1。
2.2 tSPNL对小鼠大脑海马GABA、BDNF水平的影响
与正常对照组比较,模型组小鼠海马GABA、BDNF水平明显降低,差异有统计学意义(P < 0.05或P < 0.01)。与模型组比较,BDZ组GABA、BDNF水平增加,差异有统计学意义(P < 0.05或P < 0.01);tSPNL 500组GABA、BDNF水平均明显升高,差异有统计学意义(P < 0.05或P < 0.01);tSPNL 100组GABA、BDNF水平有升高的趋势,但差异无统计学意义(P > 0.05)。提示在500 mg/kg剂量下tSPN能升高小鼠海马内BDNF和GABA水平。见图2。tSPNL具有升高老龄小鼠皮层BDNF、GABA的水平,但差异无统计学意义,在本文中未列出该数据图表。
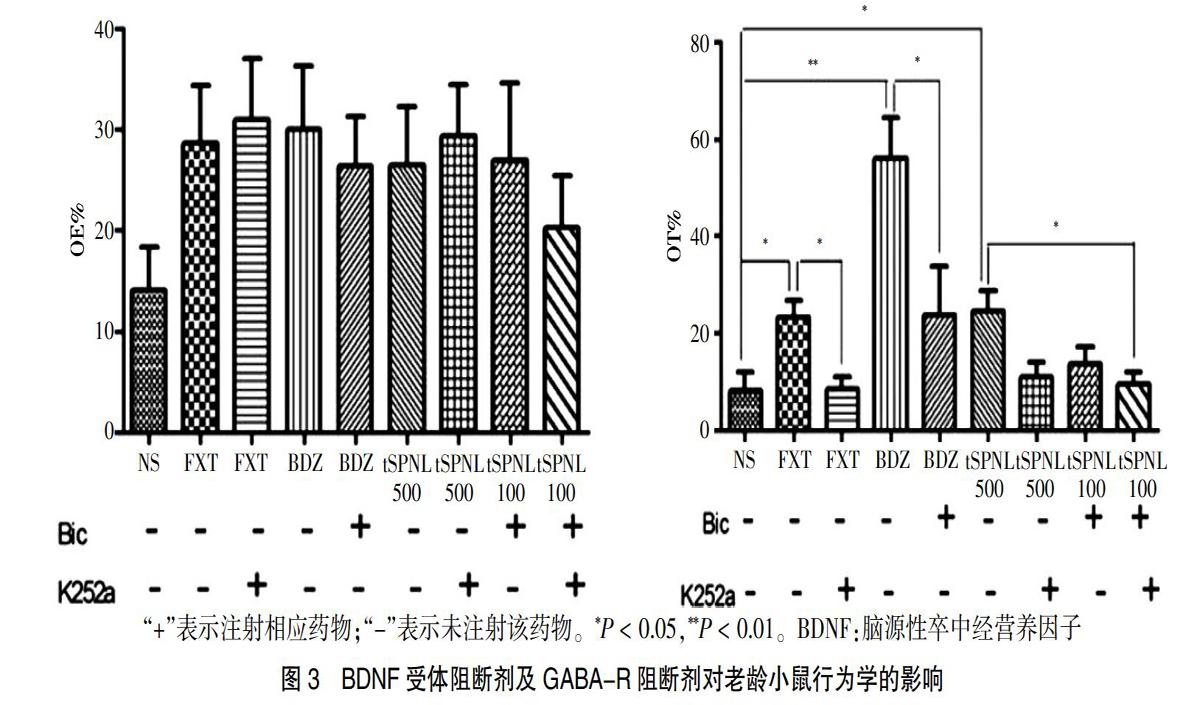
2.3 阻断BDNF-GABA通路对tSPNL抗老年焦虑作用的影响
与模型组比较,FXT组、BDZ组及tSPNL 500均可增加老龄小鼠OT%。各给药组在给予相应的受体阻断剂后:①BDZ组可显著提高小鼠OT%,注射Bic后OT%降低,差异有统计学意义(P < 0.05);②FXT组的OT%增加,注射K252a后OT%降低,差异有统计学意义(P < 0.05);③tSPNL 500组的OT%增加,分别注射K252a或Bic后,其OT%有降低趋势,但差异无统计学意义(P > 0.05);同时注射K252a及Bic后OT%降低,差异有统计学意义(P < 0.05)。提示tSPNL抗焦虑作用与BDNF-GABA信号通路有关。见图3。
3 讨论
临床上治疗老年焦虑症的药物主要是苯二氮卓类(BDZ)和五羟色胺再摄取抑制剂(SSRIs)SSRIs两类。但老年人对BDZ类药物的不良反应敏感性增加,容易导致认知障碍、精神运动性障碍以及依赖性,使其应用受到限制[11]。SSRIs起效慢、治疗初期有兴奋和致焦虑的风险以及增加性功能障碍、诱发老年人缺血性脑卒中的危险[12-13],实际上仍不能成為理想的抗老年焦虑药。开发疗效显著、特异性针对老年焦虑症的新型抗焦虑药物至关重要。
脑内GABA能神经可塑性障碍是老年焦虑症的重要发病机制之一[14],本研究选用老龄小鼠焦虑模型研究tSPNL抗老年焦虑症的作用,并探讨其对GABA能神经可塑性障碍的影响。行为学结果显示,与正常对照组比较,模型组动物在高架十字迷宫中的OT%及OE%减少,表现出明显的焦虑行为。与模型组比较,BDZ组和tSPNL组小鼠OT%与OE%增加,提示tSPNL具有抗老年焦虑的作用,且在500 mg/kg的剂量下抗焦虑作用最佳。研究证明,BDNF通过GABAA-R调控GABA能神经的可塑性,其分泌减少导致GABA能神经受损可塑性降低[15-16]。给予外源性BDNF可促进神经再生、GABAA-R表达增加、焦虑症状减轻[17-18],改善认知功能[19]。此外,近期研究[20]报道,利用具有神经保护作用的二甲双胍可能过CREB/BDNF信号通路对抗甲基苯丙胺所致大鼠焦虑、抑郁、认知障碍和神经退行性变,提示药理干预可通过BDNF信号通路发挥抗焦虑作用。本课题组前期研究发现老龄小鼠脑内BDNF、GABA水平下降,焦虑易感性增加,其焦虑水平与BDNF、GABA水平成正相关关系[8]。本研究结果显示,tSPNL组老龄小鼠海马内GABA、BDNF水平升高,与行为学结果一致,提示tSPNL抗焦虑作用与增加脑内GABA、BDNF水平有关。脑室注射分别注射TrKB阻断剂K252a或GABAA-R阻断剂Bic后,tSPNL抗焦虑作用降低,但差异无统计学意义,该结果提示BDNF-TrKB或GABAA-R通路可能不是其独立的抗焦虑作用机制。细胞内信号通路并不是独立的,各条信号通路之间存在交叉通信,BDNF和GABA都有其不同的上、下游通路。同时注射K252a及Bic后,可消除tSPNL的抗焦虑作用,OT%明显降低,提示tSPNL抗老年焦虑的作用与BDNF-GABA信号通路有关。以上结果提示,tSPNL具有抗焦虑作用,可能通过增加BDNF水平恢复GABA能神经可塑性,从而减轻老年焦虑行为的易感性。
本研究结果为老年焦虑症的治疗提供了潜在的药物靶点,即通过补充BDNF恢复GABA能神经可塑性从而改善老年焦虑症症状;同时本研究结果也为老年焦虑症的治疗提供了潜在的候选药物。但本研究也存在局限性:①未进行tSPNL与BDZ、SSRIs优势作用的比较,如BDZ可引起认知功能障碍,tSPNL是否可改善认知功能;②使用受体阻断剂后未进行相关分子生物学指标检测。更多的机制尚需进一步研究。
[参考文献]
[1] Craske MG,Stein MB,Eley TC,et al. Anxiety disorders [J]. Nat Rev Dis Primers,2017,3:17024.
[2] Blazer,DG. Depression in late life:review and commentary [J]. J Gerontol A Biol Sci Med Sci,2003,58(3):249-265.
[3] Nuss P. Anxiety disorders and GABA neurotransmission:a disturbance of modulation [J]. Neuropsychiatr Dis Treat,2015,11:165-175.
[4] Duveau V,Laustela S,Barth L,et al. Spatiotemporal specificity of GABAA receptor-mediated regulation of adult hippocampal neurogenesis [J]. Eur J Neurosci,2011,34(3):362-373.
[5] Nikolaus S,Wittsack HJ,Beu M,et al. GABAergic Contyol of Nigrostriatal and Messolimbic Dopamine in the Rat Brain [J]. Front Behav Neurosci,2018,12:38.
[6] Luscher B,Fuchs T,Kilpatrick CL. GABAA receptor trafficking-mediated plasticity of inhibitory synapses [J]. Neuron,2011,70(3):385-409.
[7] Porcher C,Hatchett C,Longbottom RE,et al. Positive feedback regulation between gamma-aminobutyric acid type A (GABA(A) receptor signaling and brain-derived neurotrophic factor (BDNF) release in developing neurons [J]. J Biol Chem,2011,286(24):21 667.
[8] Zhou NN,Zhu GB,Lin Q,et al. GABAergic dysfunction in late-life anxiety:the role of BDNF [J]. European Neuropsychopharmacology,2015,25(2):S253.
[9] 缪吉,雷涛,杨光显,等.三七皂苷对运动性贫血大鼠海马BDNF表达的影响[J].中国康复,2017,32(6):484-487.
[10] 张华林,李中,周中流,等.三七葉总皂苷对抑郁大鼠脑内单胺及神经因子的影响[J].现代食品科技,2015(12):32-41.
[11] Allain H,Bentue FD,Polard E,et al. Postural instability and consequent falls and hip fractures associated with use of hypnotics in the elderly:a comparative review [J]. Drugs Aging,2005,22(9):749-765.
[12] Nutt DJ. Overview of diagnosis and drug treatments of anxiety disorders [J]. CNS Spectr,2005,10(1):49-56.
[13] Tanguay Bernard MM,Luc M,Carrier JD,et al. Patterns of benzodiazepines use in primary care adults with anxiety disorders [J]. Heliyon,2018,4(7):88.
[14] 石安华,孙晓菲,周宁娜.老年焦虑症GABA能神经可塑性障碍及BDNF的潜在治疗作用[J].中华行为学与脑科学杂志,2016.25(6):572-576.
[15] Shinoda Y,Sadakata T,Nakao K,et al. Calcium-dependent activator protein for secretion 2 (CAPS2) promotes BDNF secretion and is critical for the development of GABAergic interneuron network [J]. Proc Natl Acad Sci U S A,2011,108(1):373-378.
[16] Zhu G,Sun X,Yang Y,et al. Reduction of BDNF results in GABAergic neuroplasticity dysfunction and contributes to late-life anxiety disorder [J]. Behav Neurosci,2019,133(2):212-224.
[17] Muller HD,Neder A,Sommer C,et al. Different postischemic protein expression of the GABA(A) receptor alpha2 subunit and the plasticity-associated protein MAP1B after treatment with BDNF versus G-CSF in the rat brain [J]. Restor Neurol Neurosci,2009,27(1):27-39.
[18] Casarotto PC,Bortoil VC,Correa FM,et al. Panicolytic-like effect of BDNF in the rat dorsal periaqueductal grey matter:the role of 5-HT and GABA [J]. Int J Neuropsychopharmacol,2010,13(5):573-582.
[19] Xu AH,Yang Y,Sun YX,et al. Exogenous brain-derived neurotrophic factor attenuates cognitive impairment induced by okadaic acid in a rat model of Alzheimer′s disease [J]. Neural Regen Res,2018,13(12):2173-2181.
[20] Keshavarzi S,Kermanshahi S,Karami L,et al. Protective role of metformin against metham-phetamine induced anxiety,depression,cognition impairment and neurodegeneration in rat:The role of CREB/BDNF and Akt/GSK3 signaling pathways [J]. Neurotoxicology,2019,72:74-84.
(收稿日期:2018-12-06 本文編辑:任 念)
