闽西山区养殖棘胸蛙两性异形
2019-10-30王茂元赖铭勇黄洪贵黄柳婷胡振禧吴妹英林学文
王茂元,赖铭勇,黄洪贵,黄柳婷,田 田,胡振禧,吴妹英,林学文
(福建省淡水水产研究所,福建 福州 350002)
动物界普遍存在着两性异形现象[1],这主要表现为形态大小、体色以及局部特征等外部生物学特征上的差异,尤其是两栖动物两性异形存在多种类型[2-4]。长期以来,决定两性异形的原因倍受人们关注,而对于个体大小的性别差异,生育力选择(Fecundity selection)假说被认为是促成偏向雌性为较大个体的两性异形的驱动力[5];性选择(Sexual selection)假说认为雄性间的争斗促进了雄体大于雌体的两性异形的形成[6];生态位分离(Niche-separation)假说认为两性个体在生态位上的分离与差异是两性异形出现的原因[7]。然而两性异形的形成,还可能与生态位、年龄或生长率、食性、繁殖策略及其生长发育各阶段的物质和能量分配上的分离与差异相关。因此,开展养殖动物两性亲体差异研究,探讨两性异形与动物性选择之间的关系具有重要学术价值。
棘胸蛙(Paaspinosa)俗称石蛙、石鳞、石鸡、石冻等,属两栖纲(Amphibian),无尾目(Anura),蛙科(Ranidae),棘蛙属(Rana),主要分布在我国南方丘陵溪流区。棘胸蛙肉质细腻,味道甘美,具有很高的食用和药用价值,是传统医食兼用的名贵动物,素有“百蛙之王”的美誉,不仅经济价值高,而且市场需求量大[8]。近年来由于人为捕杀和生存环境被破坏,野生棘胸蛙数量逐渐减少,被中国物种红色名录列为易危(Vu)等级[9]。为解决棘胸蛙野生种质资源衰退和市场消费需求扩大的矛盾,国内外一些学者在棘胸蛙种质生物学、繁殖与发育生物学以及人工养殖技术等方面开展了较为深入的研究[10-13],而对于其异形的研究较少,路庆芳等[14]指出平江性成熟棘胸蛙的雄性个体体长和体质量显著大于雌性;杨程等[15]采用聚类分析、主成分分析和判别分析方法研究了四明山脉溪流区棘胸蛙同生群两性亲体间的形质差异;Yu等[1]从生态地理环境方面揭示了棘胸蛙两性异形现象。
虽然有些学者就国内其他地区的棘胸蛙异形现象进行了报道,但也多是集中在生长繁育与异形关系的研究[14-16],未详细分析形态性状与成体体质量的关系以及建立多元回归模型。为此,本文通过测定养殖棘胸蛙的外部形态特征指标,分析了棘胸蛙两性成体间的形质差异,并分别建立了雌雄棘胸蛙体质量与相关体尺的多元回归方程,旨在为今后指导棘胸蛙两性选育及系统化培育提供参考依据。
1 材料与方法
1.1 研究材料
2017年10月从闽西山区棘胸蛙养殖场随机选取3龄当地棘胸蛙养殖群体,该群体为野生群体经过2代繁殖选育后的子代,放养在水泥池中(密度30只/m2),投喂黄粉虫。试验时取雌、雄个体各82只作为测定群体。
1.2 生物学指标测定
将采集到的活蛙进行编号,使用数显游标卡尺(精确度0.01 mm)和高精度电子天平(精确度0.01 g)进行生物学指标测定,由同一人完成。测定指标为:体质量(Body weight,BW)、头体长(吻端至泄殖腔孔后缘的水平距离,Snout-urostyle length,SUL)、吻长(吻端至眼前角的水平距离,Snout length,SL)、头长(吻端至颈褶的水平距离,Head length,HL)、肩肛距(颈褶至肛孔后缘的的水平距离,Shoulder-to-anal length,SAL)、前足长(前肢基部至第三指末端的直线距离,Froeofoot length,FFL)、前肢长(肩关节至第三指末端的直线距离,Fore limb length,FLL)、后足长(后肢基部至最长趾末端的直线距离,Hindfoot length,HFL)、后肢长(体后端正中部位至第四趾末端的直线距离,Hindlimb length,HLL)、眼间距(左右上眼睑内侧缘间的水平距离,Interorbital space,IS)、鼻间距(左右鼻孔间的水平距离,Internasal distance,INS)、头宽(头部左右两侧的最大水平距离,Head width,HW)、肩宽(左右肩点间的水平距离,Shoulder width,SW)和眼径(与体轴平行的眼的长度,Eye diameter,ED)。具体测量起始位置见图1。

注:1.头体长;2.头长;3.头宽;4.前肢长;5.前足长;6.后足长;7.后肢长;8.肩宽;9.眼间距;10.鼻间距;11.吻长;12.眼径;13.肩肛距。
Notes:1.Body weight;2.Head length;3.Head width;4.Forelimb length;5.Froeofoot length;6.Hindfoot length;7.Hindlimb length;8.Shoulder width;9.Interorbital space;10.Internasal distance;11.Snout length;12.Eye diameter;13.Shoulder-to-anal length.
1.3 数据处理和分析
利用Excel 2007和SPSS 17.0软件进行统计处理与检验分析。用Kolmogorov-Smirnov和Bartlett进行数据的正态性和方差同质性检验,用单因子方差分析(One-way ANOVA)比较两性间外部形态差异,用其余性状与体质量进行相关性分析。通过偏回归分析上述数据,保留那些偏回归系数达到甚至超过显著水平(P<0.05)的形态性状并建立这些表型性状对体质量的多元回归方程,以此进行拟合度检验。描述性统计值用平均值±标准差表示,显著性水平设置为α=0.05。
两性异形指数[17]计算公式为:两性异形程度指数=1-体型较小性别的平均头体长/体型较大性别的平均头体长。
2 结果与分析
2.1 生物学测量指标间的性别差异比较
从表1可以看出,闽西山区棘胸蛙雌雄两性在外部生物学性状测量指标间存在一定程度的差异。棘胸蛙雌性平均头体长为(98.17±5.84)mm,雄性的平均头体长为(105.36±6.79)mm,雄性体长明显大于雌性头体长,雄性头体长与雌性头体长的比值为1.073,异形程度指数为0.068。除吻长、眼间距和眼径等3项形态性状测量值在雌、雄两性之间无差别外,其余11项测量值均呈现出雄性显著大于雌性(P<0.05)现象;除眼间距、前肢长、后肢长、眼径、肩肛距等5项形态性状测量值的变异系数呈现出雌性略微大于雄性外,其余9项测量值的变异系数均呈现出雌性小于雄性的现象。上述结果表明:棘胸蛙在生长速度和体型可塑性上均存在较为明显的性别差异;雌性的体型稳定性好于雄性,但增长速度慢于雄性。

表1 棘胸蛙两性生物学测量性状参数统计
注:同行上标不同小写字母表示组间差异显著(P<0.05)。下同。
Notes:The different letters on the parameters on the same row meant significant difference(P<0.05).The same as below.
2.2 体尺性状比的性别差异比较
根据所测结果,计算棘胸蛙体尺性状比的性别差异指标。由表2可以看出,雌雄两性亲体在体尺性状比指标间基本没有差异。除前足长/前肢长、头宽/肩宽比值略小于雌性外,雄性的其余形质比都等于或大于雌性;除肩肛距/头宽、头宽/肩宽、鼻间距/眼间距变异系数大于雄性外,雌性的其余6项形质比均小于雄性。

表 2 棘胸蛙两性体尺性状比参数统计
2.3 生物学测量指标与体质量间的相关分析
对棘胸蛙雌雄两性生物学测量指标与体质量进行相关分析。由表3可见,棘胸蛙雌、雄两性形态性状测量指标与体质量间的相关性存在一定程度的异同。从Pearson相关性来看,雄性测定群体中,除肩宽与体质量未呈显著相关(P>0.05)外,其余12项测量指标与体质量均呈现显著相关(P<0.05);雌性测定群体中,除眼间距、眼径与体质量未呈显著相关(P>0.05)外,其余11项测量指标与体质量呈现显著相关(P<0.05)。但从偏相关分析结果可以看出,排除其他测量指标对体质量的影响,在13项测量指标中,雄性头体长、吻长和眼径与体质量间存在显著相关(P<0.05);雌性头体长、前肢长和肩宽与体质量间存在显著相关(P<0.05)。

表 3 棘胸蛙两性生物学测量性状与体质量的相关性(n=82)
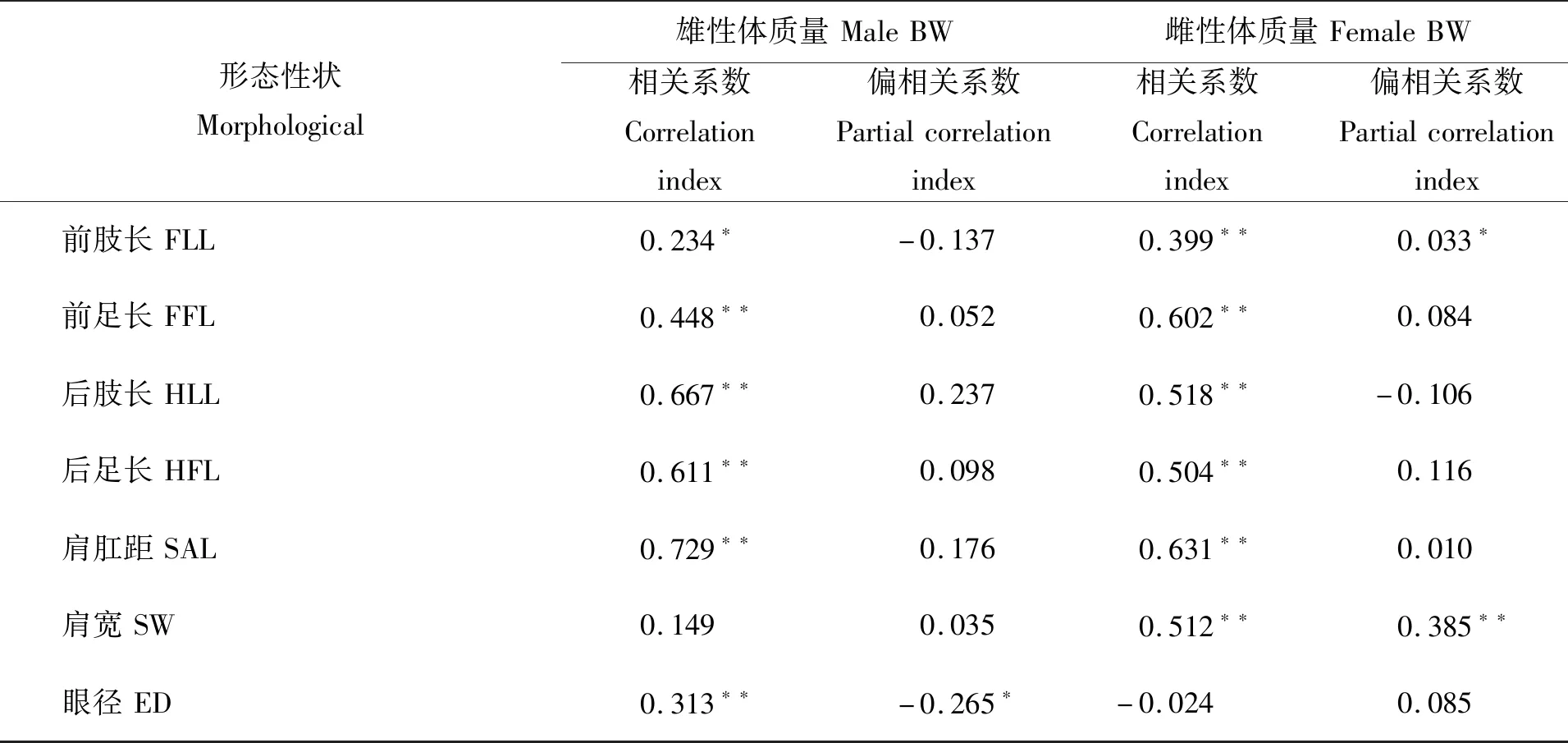
续表3
注:*表示性状间相关系数达到显著水平(P<0.05),**表示性状间的相关性达到极显著水平(P<0.01)。
Notes:*meant the correlation coefficient between traits reached the significant level(P<0.05),**meant the correlation between traits reached the extremely significant level(P<0.01).
2.4 多元回归分析
对形态性状进行回归分析,各回归系数t检验结果表明:雄性棘胸蛙的头体长、眼间距、吻长和头宽对体质量影响显著;雌性棘胸蛙的头体长、肩宽和前肢长对体质量影响显著。因此,从回归方程中剔除不显著形态性状,以建立最优模型。
回归中,R2值越大反映自变量与因变量的共变量比率越高,模型与数据拟合度越好。根据表4,可以看出雄性棘胸蛙回归模型4的R2值最大,定为最优模型,从模型4回归系数显著性结果(表5)可得回归方程:雄性体质量=3.620×头体长+0.626×后肢长-5.847×眼径+0.728×吻长-269.868,回归显著性检验结果表明建立的回归方程有效。同时从表5可见,雄性棘胸蛙被保留下来的形态性状对体质量的影响,依次呈现为体长(3.620)>吻长(0.728)>后肢长(0.626)>眼径(-5.847),表明体长对体质量的贡献最大,而眼径最小。

表 4 雄性棘胸蛙体质量相关性多元回归方程模型汇总

表 5 雄性棘胸蛙多元回归模型4的回归系数以及显著性检验结果
同理,根据表6,可以看出雌性棘胸蛙回归模型3的R2值最大,定为最优模型,从模型3回归系数显著性结果(表7)可得回归方程:雌性体质量= 2.914×头体长+1.502×前肢长+1.521×肩宽-304.518,回归显著性检验结果表明建立的回归方程有效。同时从表7可见,雌性棘胸蛙被保留下来的形态性状对体质量的影响,依次呈现为体长(2.914)>肩宽(1.521)>前肢长(1.502),表明体长对体质量的贡献最大,肩宽和前肢长次之。

表 6 雌性棘胸蛙体质量相关多元回归方程模型汇总

表 7 雌性棘胸蛙多元回归模型3的回归系数以及显著性检验结果
综上所述,雌雄棘胸蛙的体长是影响棘胸蛙体质量的决定性公共变量,也是影响其体质量的关键性状。
3 讨论
动物的两性异形现象非常普遍,主要表现为个体大小、体色以及局部形态特征(婚垫、胸腹部分泌物和声囊、肛门异形、四肢异形等)差异等方面[18-21]。有研究显示,两栖动物中约80%的有尾目和95%的无尾目存在两性异形[6]。两栖类体型异形主要存在3种类型:成年雄性体型大于成年雌性;成年雌性体型大于成年雄性;成年两性个体大小没有显著差异[22]。其中雌性体型偏大类型较为普遍,而雄性体型偏大类型在无尾两栖类中较为少见,约占5%[6]。从异性程度看,闽西山区棘胸蛙养殖群体雄性头体长与雌性头体长的比值为1.073,异形程度指数为0.068,与泽陆蛙(Fejervaryalimnocharis)的0.080[23]相当,而与花臭蛙(Odorranaschmackeri)的0.496、大绿臭蛙(O.margaretae)的0.476和凹耳蛙(O.tormota)的0.418异性程度[24-25]相比较低。从其他外部生物学形态测量指标来看,除吻长、眼间距和眼径等3项形态性状测量值在雌、雄两性之间无明显差别外,体质量、头长、肩肛距、肩宽、前肢长、后足长、后肢长、鼻间距、头宽、前足长等指标均呈现出雄性显著大于雌性(P<0.05)。此外,头部异形在两栖动物中较为常见,而本研究中闽西山区雄性棘胸蛙的头长和头宽显著大于雌性(P<0.05),与Schoener 等[26]认为头部较大的个体具有摄取更大食物的潜能,而有利于增加可供选择的食物种类,获得更大的摄取优质食物资源的机会,并在资源竞争中获得优势的观点相吻合。因此,闽西山区棘胸蛙应归为成年雄性体型大于成年雌性的类型,且属于两栖蛙类中雌雄异形程度较低的一类。
福建野生棘胸蛙在低海拔和高海拔地区均有分布[27],表明棘胸蛙与其他种类蛙相似,栖息环境因子的地理差异对两性异形的影响可能不是主要的,自然选择和系统演化历史可能是两性异形的决定因子[28-29]。目前,解释动物两性异形的主要理论为性选择假说[7],两性异形可能是通过同性成员间的配偶竞争(性内选择)、或通过偏爱异性的某个独特特征(性间选择)、或者两条途径兼而有之[30]而实现的。动物繁殖过程中两性都要消耗能量,但消耗方式不同[31],两栖动物在繁殖投入上雄性远低于雌性,较大的雌性个体具有较高的生育力和繁殖成功率[32-33],这也可能导致不同性别的个体在性成熟之后表现为两性异形。此外,在繁殖过程中,为了提高受精和繁殖的成功率,雄蛙往往通过增大个体以增强在婚配斗争中的有利地位[6],而雌蛙则更多将能量分配于繁殖,限制了个体生长,最终表现出雄大雌小的两性异形。本研究中,除头体长外,其他的形态性状对体质量的直接影响作用均较小,无疑揭示了闽西山区棘胸蛙雌、雄成熟个体为谋求繁殖利益的最大化,在性选择压力的作用下强化并保留了各自具有一定有利特征的形态性状。
综上所述,闽西山区棘胸蛙亲体间两性异形,且雄性大于雌性,更多地倾向于性选择假说和生育力选择假说,其本质是物种自然选择和系统演化的结果。
