不同林龄油松土壤微生物、酶活性和养分特征
2019-10-19范媛媛李启迪
范媛媛, 李 懿, 李启迪
(1.山东师范大学 济南 250000; 2.阿坝师范学院 资源与环境学院, 四川 阿坝 623002; 3.山东职业学院生物系,济南 250000)
1904年,德国科学家Lorenz Hiltner最早提出了根际(Rhizosphere)概念,将土壤学研究扩展至根际微域环境,引发学界瞩目[1-3]。与此同时,根际研究的内容偏重于土壤中的物质循环、微生物含量及养分的作用、土壤酶含量等方面,并使用了更多、更新的研究方法。这些研究方法的运用,对优化与调节土壤生态系统起到了至关重要的作用,并为其提供了相关的理论基础,为实践提供了借鉴[4]。目前,学界针对土壤退化、土壤修复、园林绿化、人工造林等领域的植物根际研究比较多,并且大多集中在根际土壤与非根际土壤之间土壤特性的差异性方面[5-6]。如果想了解土壤的质量和养分情况,在研究植物的根际土壤与非根际土壤的特性时,特别要注意土壤酶活性、微生物数量及养分等特点[7]。浏览研究的报道会发现,想要探究造林灌木树种在根际效应与土壤特性之间的改良情况,需要考虑灌木树种间根系活动的规律、土壤肥力影响机制等因素[8-9]。要想提高造林后土壤肥力,需要探寻根际土壤和与非根际土壤之间的关系,挖掘两者对土壤肥力的影响与作用。作为大陆高原半干旱气候的代表,岷江流域的土壤情况不良,贫瘠且流失严重,植被绿化率也不理想,迫切需要进行人工造林植被恢复工作,树种筛选则是其中的重中之重[10]。所以,想要有效筛选与评价造林树种,必须要从植物种植后根际和非根际土壤的特性着手,梳理其人工种植植物对土壤的改善效果和影响才能实现[11-12]。
油松(Pinustabuliformis)常年保持绿色,是一类乡土树种,广泛分布于我国亚热带地区,隶属于松科Pinaceae松属Pinus,环境适应性很强,具有较强的生长发育特性[13],对修复土地具有奇效,往往作为首选树种出现,能够在一定程度上使森林保持良好的生态系统。但是,研究者发现,多年来油松人工林地力衰退问题日益突出,营造油松人工林作为一项关键技术而被重视,其目的是提高土壤肥力和酶活性[14]。
林木和土壤在进行物质与能量交换时,往往需要借助根部达成,根部通过吸收周围土壤中的营养或者分泌各种有机物与元素,使土壤的性质发生一定的变化[13]。文章的着重点在于分析不同林龄的油松林根际与非根际土壤在酶活性、微生物和养分等方面的异同,从而发现土壤肥力变化的内在规律,林木与土壤之间互相影响的关系,为我国人工油松林的培育工作提供数据支撑。
1 材料与方法
1.1 研究区概况
岷江是长江支流之一,从岷山南麓流出,沿线形成杂谷脑河、大渡河、黑水河、马边河等支流,曲折回环,落差可达3 560 m,占据了13.3万km2的面积,其干流长度为735 km。岷江的上中下游三部分各有特色,上游在都江堰市,中游在都江堰市至乐山市,下游在乐山至宜宾。上游河段多河流峡谷,地形较复杂,不适宜人类耕种,因此耕地和人口较少;而中游和下游河段的地形单一,多平原,利于开展农耕,人口较多,对农业和工业都有带动作用,同时交通较为发达,开展研究的主要区域。该区域主要为温带—亚寒热带气候,从上游至下游,气温逐渐升高,年平均气温达17℃左右,最高和最低气温分别为-4℃和38℃左右。该区域的汛期集中在6—9月的夏秋季节,多暴雨,降水量较大,占全年雨量的80%以上,所以水能资源和自然资源丰富,耕地较多,且多为紫色土。
试验时间为2018年8月,选取地域范围为岷江上游,选取对象为(幼林、中林、成熟林和过熟林)油松样地,针对每种林龄油松,选择了3个重复样地(50 m×50 m),并在每个样地上设置3个10 m×10 m的小样方,并各选择3株油松,采取Riley和Barber的抖落法,在做好标记的每株油松附近挖取根际与非根际土壤,在挖取根际土壤时,必须要保证根系的完整,采取各种办法,如使用小刷子等尽可能清除外围土壤,尤其注意防止损害根系。对于采集好的土壤,先去除杂物,之后分为两部分,分别处理。一部分平铺风干并过筛后,用于测定土壤中养分的情况和酶活性,另一部分在4℃环境中下贮藏,可测定土壤中微生物的数量以及pH值。
1.2 测定方法
平板梯度稀释法是测定土壤微生物数量的基本方法,往往会选取高氏一号琼脂培养基、马丁氏培养基和牛肉膏蛋白胨琼脂等,分别作为放线菌、真菌和细菌的培养基。还要在每一浓度梯度处进行特别设置,即3个重复。而测定有机碳、土壤pH、有效钾和有效磷等元素,则要参考包士旦编《土壤农化分析》中的标准[15]。
通过在Biolog-Eco下进行72 h的温育处理,能够得到土壤多样性的数据,然后选取大约10 g烘干土壤的土样,将其放入NaCl溶液中,调至浓度为0.145 mol/L,再进行30 min左右的振荡,再选择其中的100 μl,于Eco板后读数,并在25℃下恒温培养216 h,读数间隔为12 h;接着,还要将NaCl溶液稀释,继续进行恒温培养,做好吸光值记录。
在分析微生物对碳源的利用情况时,需要依据其对碳源的反应得出结论,一般用平均每孔颜色变化率AWCD表示,当微生物丰度越高时,该值的数额越大,计算如下方式[7-9]:
AWCD=∑[(Ci-R)/31]
式中:Ci,R分别表示第i孔、对照孔的吸光值。
H=-∑Pi(lnPi)
式中:Pi为第i孔的相对吸光值比值,计算公式:
Pi=(Ci-Ri)/∑(Ci-Ri)
S=被利用碳源的总数
Ds=1-∑Pi
1.4 数据分析
进行数据统计和方差检验,需要采用Excel 2007和SPSS 18.0等软件,检验各指标之间的相关性,则采用平均值±标准误差(Mean±SE),单因素方差分析(One-way ANOVA),Pearson相关系数法等方式。
2 结果与分析
2.1 不同林龄油松根际与非根际土壤养分
由表1可知,生长着不同林龄的油松土地上,土壤中的pH值因是否为根际土壤而不同,与根际土壤的pH值较低相比,非根际土壤的pH值较高,这说明油松根际在持续地分泌有机化合物,土壤的pH值不稳定,处于变化中,呈现弱酸性,并且随着林龄的增大而增长。土壤中根际和非根际有机碳含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有机碳含量显著高于非根际(p<0.05),过熟林根际有机碳含量与非根际差异并不显著(p>0.05);根际土壤有机碳含量在幼林、中林、成熟林和过熟林分别高于非根际15.98%,17.63%,10.81%和15.31%。土壤中根际和非根际全氮含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际全氮含量显著高于非根际(p<0.05),过熟林根际全氮含量与非根际差异并不显著(p>0.05);根际全氮含量在幼林、中林、成熟林和过熟林分别高于非根际13.95%,12.39%,11.57%和3.33%。土壤中根际和非根际有效磷含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有效磷含量显著高于非根际(p<0.05),过熟林根际有效磷含量与非根际差异并不显著(p>0.05);根际有效磷含量在幼林、中林、成熟林和过熟林中分别高于非根际16.98%,10.90%,24.28%和1.81%。土壤中根际和非根际有效钾含量随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际有效钾含量显著高于非根际(p<0.05),过熟林根际有效钾含量与非根际差异并不显著(p>0.05);根际有效钾含量在幼林、中林、成熟林和过熟林分别高于非根际20.23%,12.59%,10.14%和6.37%。
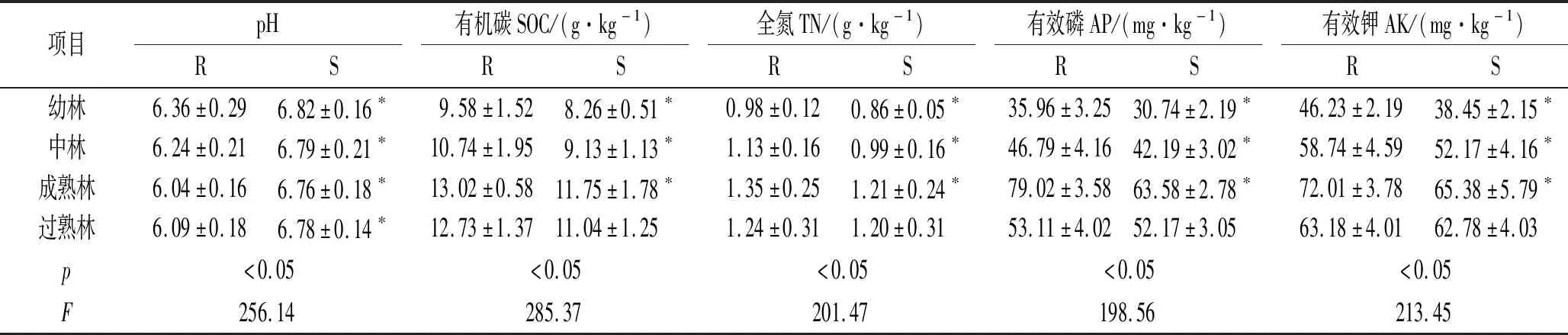
表1 不同林龄油松根际与非根际土壤养分
注:R表示根际,S表示非根际,*表示根际与非根际差异显著(p<0.05),同列不同小写字母表示不同林龄差异显著(p<0.05),下同。
2.2 不同林龄油松根际与非根际土壤酶活性
由表2可知,土壤中根际和非根际蔗糖酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际蔗糖酶活性显著高于非根际(p<0.05),过熟林根际蔗糖酶活性与非根际差异并不显著(p>0.05);根际蔗糖酶活性在幼林、中林、成熟林和过熟林分别高于非根际28.00%,33.73%,27.66%和12.48%。土壤中根际和非根际碱性磷酸酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际碱性磷酸酶活性显著高于非根际(p<0.05),过熟林根际碱性磷酸酶活性与非根际差异并不显著(p>0.05);根际碱性磷酸酶活性在幼林、中林、成熟林和过熟林分别高于非根际23.53%,12.62%,39.44%和4.69%。土壤中根际和非根际脲酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际脲酶活性显著高于非根际(p<0.05),过熟林根际脲酶活性与非根际差异并不显著(p>0.05);根际脲酶活性在幼林、中林、成熟林和过熟林分别高于非根际20.97%,41.83%,16.43%和8.47%。土壤中根际和非根际过氧化氢酶活性随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际过氧化氢酶活性显著高于非根际(p<0.05),过熟林根际过氧化氢酶活性与非根际差异并不显著(p>0.05);根际过氧化氢酶活性在幼林、中林、成熟林和过熟林分别高于非根际27.57%,14.73%,9.60%和0.31%。
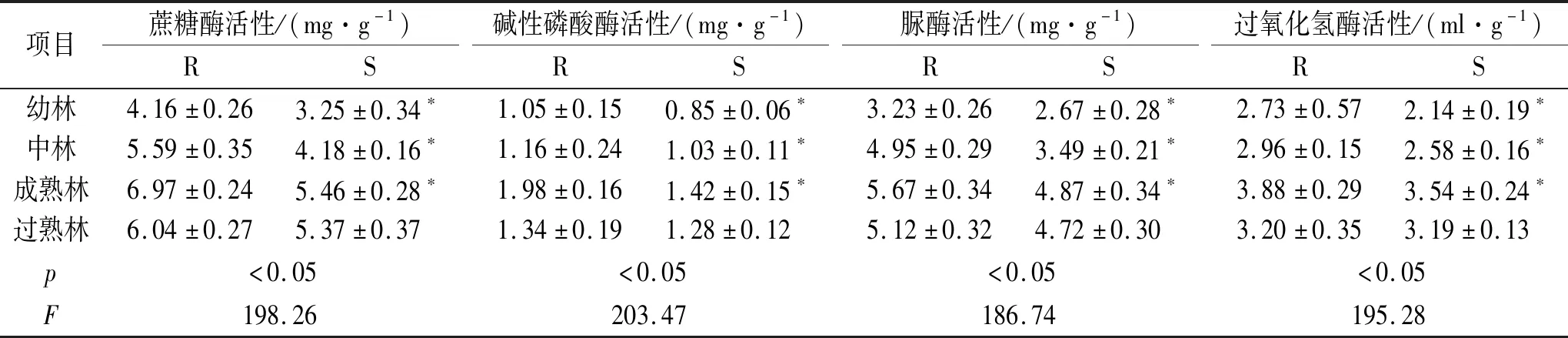
表2 不同林龄油松根际与非根际土壤酶活性
2.3 不同林龄油松根际与非根际微生物数量
通过研究比较不同林龄油松的根际和非根际土壤中的微生物数量后(表3),可知两种土壤中的微生物数量情况不同,并且与林木的生长发育阶段相关,这是受到根际分泌物影响的结果。相对而言,根际土壤更能恰如其分地表现微生物的变化规律,且总量较高。在不同林龄油松的根际土壤中,微生物数量由多到少为放线菌、真菌和细菌;在不同林龄油松的非根际土壤中,变化情况与根际土壤趋同。林龄逐渐增大,土壤中微生物总量呈先增加后降低趋势,在中龄林的根系生长状况良好,分泌物较多,活力较强,导致土壤中的微生物含量也较多,从而大大促进了有机物的分解与转化。
2.4 不同林龄油松根际与非根际土壤微生物群落
结果表明(表4),土壤中根际和非根际丰富度指数(H)随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际丰富度指数(H)显著高于非根际(p<0.05),过熟林根际丰富度指数(H)与非根际差异并不显著(p>0.05);根际丰富度指数(H)在幼林、中林、成熟林和过熟林分别高于非根际63.64%,10.76%,22.22%和11.39%。土壤中根际和非根际均匀度指数(E)随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际均匀度指数(E)显著高于非根际(p<0.05),过熟林根际均匀度指数(E)与非根际差异并不显著(p>0.05);根际均匀度指数(E)在幼林、中林、成熟林和过熟林分别高于非根际21.57%,16.42%,18.27%和1.02%。土壤中根际和非根际优势度指数(Ds)随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际优势度指数(Ds)显著高于非根际(p<0.05),过熟林根际优势度指数(Ds)与非根际差异并不显著(p>0.05);根际优势度指数(Ds)在幼林、中林、成熟林和过熟林分别高于非根际43.33%,26.83%,19.18%和2.90%。土壤中根际和非根际碳源利用丰富度指数(S)随着油松林龄的增长表现出先增加后降低趋势,在成熟林阶段达到最大值,其中幼林、中林和成熟林根际碳源利用丰富度指数(S)显著高于非根际(p<0.05),过熟林根际碳源利用丰富度指数(S)与非根际差异并不显著(p>0.05);根际碳源利用丰富度指数(S)在幼林、中林、成熟林和过熟林分别高于非根际9.40%,8.91%,9.89%和3.98%。

表3 不同林龄油松根际与非根际微生物数量
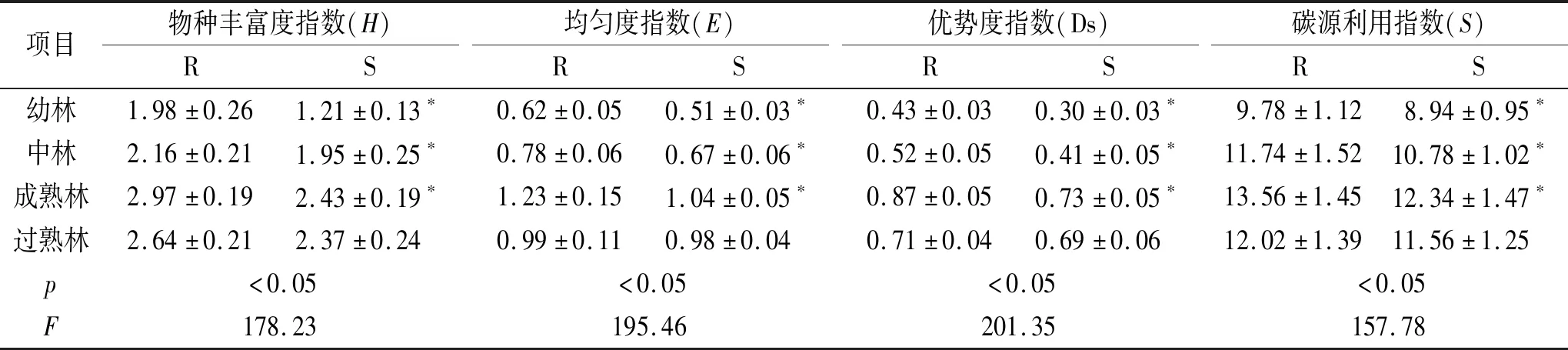
表4 不同林龄油松根际与非根际土壤微生物群落
2.5 土壤养分与微生物多样性之间相关性
土壤养分为微生物提供碳源和氮源,要对土壤微生物群落多样性与土壤养分的相关性进行分析(表5)。通过研究可发现,土壤微生物群落功能多样性与土壤养分呈正比例关系,与土壤pH值呈负比例关系,还与土壤中的微生物总数、细菌数目、蔗糖酶活性、有机碳和全氮、碱性磷酸酶活性等具有一定的关联性。由此可得出结论,微生物群落功能多样性受到微生物数量、土壤养分和土壤酶活性的直接影响。其中,土壤pH值的作用为负,微生物数量、土壤酶活性和土壤养分的作用为正,这种情况引发了不同土层土壤微生物群落多样性存在较大差异,而微生物总数、细菌数目、蔗糖酶活性、土壤有机碳和全氮以及碱性磷酸酶活性等,都会对其产生影响。

表5 土壤养分与微生物多样性之间相关性
注:*,**分别表示在0.05,0.01水平上差异显著,ns表示没有相关性(p>0.05)。
3 讨论与结论
土壤肥力的现实状况与变化情况,可以从微生物的活跃与变化中窥知一二,尤其要注意其数量、种类、生态分布及活性等内容[16],在查看微生物的总体数量时,要注意由于同种类微生物与植物根系互相作用而产生的效果,一般表现为细菌和真菌在不同土壤中的数量是不同的。产生这种情况的原因在于,放线菌的环境适应能力强,不受高温、干燥、碱性等条件的影响,占据了优势地位[17]。与此同时,研究还发现,针对不同林龄的油松,非根际土壤中的有机质和有效养分含量均低于根际土壤,并且出现一些根际聚集的现象[17]。从油松的林龄来看,土壤中的养分含量随着幼龄林长至中龄林而呈现增加现象,而在中龄林至成熟林期间,养分含量会逐渐降低。从土壤中养分的变化情况来看,在根际聚集的作用和影响下,油松能够提高以有效磷和有效钾为主要代表的土壤养分含量。油松处于幼龄时,由于根际不够发达,所分泌的产物少,转化的有机物更少,主要以地表枯落物的分解为主,而在中龄林和成熟林中,根际发达,根产物的量增加,增加了有机质的含量。而树木在不同的生长阶段,根系所释放出的有机物含量会受到光合作用的影响而产生变化,中龄林土壤中的有机物含量较高,而成熟林土壤中的有机物含量呈减少趋势,并且非根际土壤中的有机质含量低于根际土壤中,这种现象与有机质的变化类似,根际土壤有机质、全氮含量等均会受到植物生长期间根系、枯落物、根毛及根表皮分解等因素的影响[18-19]。而在研究土壤酶活性时,可以发现,该种有机质的活性大小与土壤是否处于根际相关,就不同林龄油松而言,非根际土壤与根际土壤中的有机物情况不同,这与林木根系分泌酶类物质有关,营造出比较特殊的微生物环境。一般情况下,土壤根系附近的微生物含量要高于非根际附近,而微生物一旦收到外界的刺激,会分泌大量的酶,从而使根际与非根际土壤中的酶活性产生较大的变化[20-21]。据研究,脱氢酶、过氧化氢酶及脲酶的活性在根际和非根际土壤中,大部分都会出现“低—高—低”的特征。
土壤是一个鲜活的、循环的小生态系统,无时无刻都发生着物质的分解与转化活动,这都依赖于土壤微生物的作用,不仅能够促进土壤中物质的转化与影响,还能增进土壤肥力。土壤酶最为突出地反映了土壤的生物活性,使土壤产生生物或化学反应,产生有机物,使土壤养分发生变化[22-23]。而研究也发现,微生物数量、土壤养分和酶活性存在一定的关联[22-23]。研究还发现,土壤微生物的数量多少并未受到土壤酶活性的主要作用,两者未必正相关。据推测,土壤微生物的数量受到多重因素的影响,例如根系分泌和动植物残体腐解等,其中的原因还需要深入探究[24]。很多报道认为,土壤养分的变化离不开微生物数量与酶活性的影响。本研究则发现,土壤微生物群落功能多样性的指标,与微生物总数、细菌数目、蔗糖酶活性、土壤有机碳和全氮以及碱性磷酸酶活性等因素关系显著,而与土壤pH呈负比例关系,而土壤养分、土壤酶活性和微生物数量对微生物群落功能多样性具有积极影响,这从侧面反映了不同土层土壤微生物群落多样性差异的根源。其中,微生物总数、细菌数目、蔗糖酶活性、土壤有机碳和全氮以及碱性磷酸酶活性,都对土壤产生作用,可知土壤养分与微生物、酶活性存在专性作用[25-26]。在进行研究时,要关注简单相关系数和两者之间的线性关系,也会与其他变量产生反应,所以需要深入探究,才能真正发现两个变量之间的线性相关关系,再将其运用至实践中。因此,未来应重点关注和监测根际和非根际土壤在不同季节的变化情况,从而探究在植被恢复进程中,根际和非根际土壤中的养分变化情况。
