异源表达CmSAMDC基因拟南芥的耐盐性分析
2019-10-14刘长命路梦梦明田田
刘长命,杨 娜,文 丹,路梦梦,明田田
(商洛学院,陕西 商洛 726000)
盐胁迫是植物生长发育中最为严重的非生物胁迫因素之一。目前,全球被盐渍化的陆地面积约占10%,中国的盐渍土面积近1亿hm2,并且有日趋扩大的趋势。对植物耐盐机理进行深入研究,将有助于培育耐盐性强的植物品种,并对生态环境改善也具有积极的指导意义[1]。
近年来,多胺代谢途径已被作为植物潜在抗性的研究目标,涉及多胺代谢的相关基因也越来越被重视。研究发现,在遭受冷害[2]或盐胁迫[3]时,黄瓜和向日葵的内源多胺合成与代谢途径都发生了改变,并且抗性品种能合成更多的多胺来抵御胁迫[4-6]。
19-腺苷甲硫氨酸脱羧酶(SAMDC)是精胺和亚精胺合成过程中的关键酶,该酶参与了多数植物的抗性反应过程[7]。作者前期研究[8-11]显示,转CmSAMDC基因的拟南芥植株对白粉病的抗性明显增强。为了进一步探明CmSAMDC基因在响应非生物胁迫中的作用,本研究以已获得的转基因拟南芥为材料,对其耐盐性进行了分析,以期为利用该基因进行甜瓜耐盐性改良提供参考。
1 材料与方法
1.1 材料
转基因拟南芥株系TA1、TB1为前期研究获得,将野生型和转基因拟南芥T3代种子消毒和清洗干净,待用。
1.2 方法
1.2.1 目标基因的定量和半定量表达分析 选取6周龄T3代转基因拟南芥植株叶片,利用引物CmSAMDC F:5′-ATCAAAACTTGCGGCACTAC-3′和CmSAMDC R:5′-AGCACCCTCACAATCAACTTAG-3′进行定量和半定量分析。实时定量程序为:95 ℃ 30 s;95 ℃ 20 s,60 ℃ 20 s,72 ℃ 20 s,40个循环;以拟南芥的Actin2基因作为内参。半定量程序为:94 ℃ 2 min;94 ℃ 20 s,60 ℃ 30 s,72 ℃ 1 min,25个循环;72 ℃ 10 min。
1.2.2 转基因拟南芥发芽率的测定 将T3转基因植株和野生型拟南芥的种子用75%酒精和10% NaClO消毒,然后随机取100粒,播于含不同浓度(0、100、150、200、300 mmol/L)NaCl的MS培养基上,6 d后统计发芽情况。
1.2.3 转基因拟南芥幼苗的根长及侧根数测定 将MS培养基上生长15 d的T3代转基因幼苗和野生型拟南芥幼苗移至含不同浓度(0、100、150、200、300 mmol/L)NaCl的MS培养基上,继续生长15 d后测定其根长和侧根数。
1.2.4 转基因拟南芥幼苗的耐盐性鉴定 将MS培养基上生长15 d的T3代转基因幼苗和野生型拟南芥幼苗移栽入盆(培养基质为蛭石∶壤土∶草木灰=1∶1∶1),培养至4周龄时,每穴分别浇灌15 mL 200 mmol/L或400 mmol/L NaCl溶液,每2 d浇灌1次,在处理12 d后测定脂质过氧化水平(MDA含量)。
1.2.5 转基因拟南芥激素含量的测定 油菜素内酯(Brassinolide, BR)、茉莉酸甲酯(Methyl jasmonate, JA-me)、脱落酸(Abscisic acid, ABA)、赤霉素(Gibberellin, GA)、吲哚丙酸(Indole propionic acid, IPA)、玉米素(Zeatin riboside, ZR)和吲哚乙酸(Indole acetic acid, IAA)的提取参照黄志[12]的方法。
2 结果与分析
2.1 目标基因的表达分析
通过转基因植株筛选,并对转基因拟南芥T3代植株进行CmSAMDC基因表达分析,结果(图1)显示,CmSAMDC基因成功在转基因株系TA1和TB1中表达。
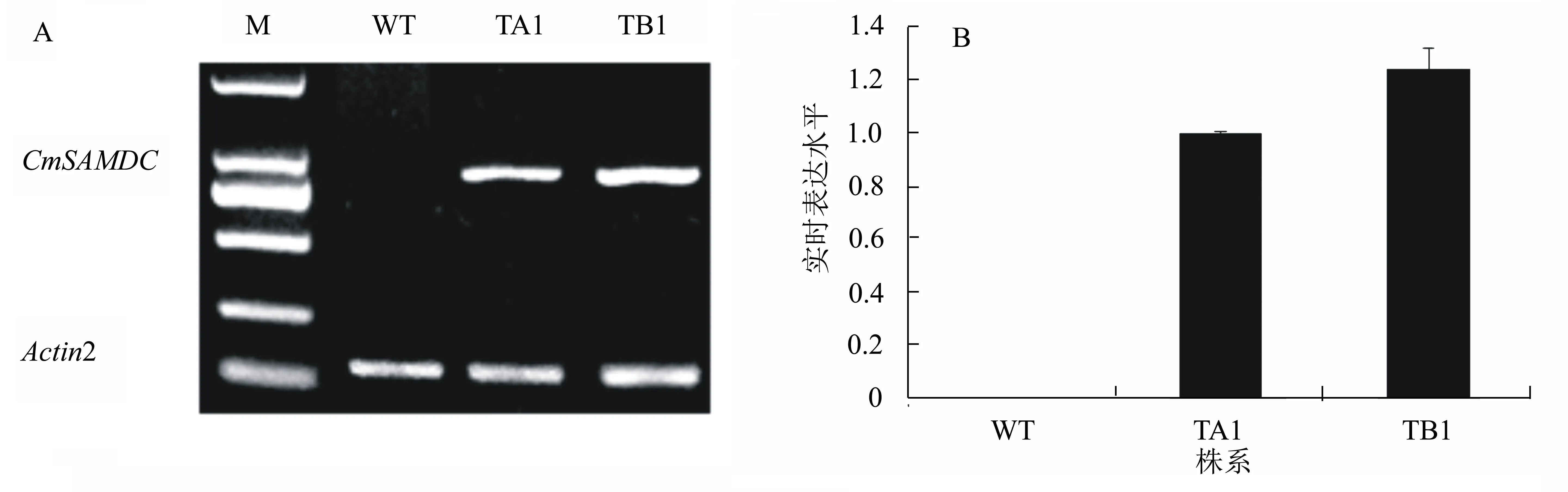
TA1和TB1代表转基因株系;WT代表野生型拟南芥。下同。
2.2 盐胁迫下转基因拟南芥的发芽率
对转基因T3代株系TA1、TB1和野生型拟南芥植株(WT)的种子发芽率进行观察,发现:在含100、150、200 mmol/L NaCl条件下,转基因T3代种子的发芽率分别为86%、70%和45%,而野生型拟南芥种子的发芽率分别为75%、58%和25%,T3代种子的发芽率均显著高于野生型植株的;而在不含盐溶液(0 mmol/L)的MS培养基上,两者的种子发芽率无显著性差异(图2)。
2.3 盐胁迫下转基因拟南芥幼苗的根长及侧根数
对根系观察结果(图3)显示:在不含盐溶液(0 mmol/L)的MS培养基上,转基因T3代植株的根长略大于野生型植株,但差异未达到显著性水平,侧根数也无显著性差异;在100 mmol/L盐胁迫条件下,T3代植株的侧根数明显多于野生型植株,差异达到了极显著水平,而根长无显著差异;在150 mmol/L盐胁迫条件下,T3代植株的侧根数和根长均显著高于野生型植株;在200 mmol/L盐胁迫条件下,两种材料的根系和侧根生长均受到了明显抑制。
2.4 盐胁迫下转基因拟南芥植株的生长情况
将MS培养基上15日龄的幼苗进行移栽,缓苗3 d后隔日浇1次不同浓度的NaCl溶液。用200 mmol/L盐溶液处理8 d后,转基因植株与对照植株(CK)生长基本一致,而野生型植株的叶片呈现轻微黄化;在处理16 d后,野生型植株明显小于转基因植株。用400 mmol/L盐溶液处理8 d后,转基因植株生长受阻,但较野生型植株的受阻程度要轻;处理16 d后,野生型植株已经死亡,而转基因植株仍能继续存活(图4A)。经不同浓度盐溶液处理后12 d,植株的脂质过氧化程度(MDA含量)存在显著性差异,即经200 mmol/L与400 mmol/L浓度盐溶液处理12 d后,TA1植株的MDA含量比对照分别上升了50%和196%, TB1比对照分别上升了28%和150%,野生型植株比对照分别上升了92%和376%。总体来看,转基因拟南芥植株的耐盐性明显强于野生型拟南芥。

图2 盐胁迫下转基因拟南芥的发芽率

A:转基因T3代和WT幼苗根系的生长情况; B:转基因T3代和WT幼苗的相对根长; C:转基因T3代和WT幼苗的侧根数。

A:盐胁迫后8 d和16 d的植株表型; B:盐胁迫后12 d的MDA含量(鲜重)。
2.5 转基因拟南芥植株激素含量的变化
对转基因株系TA1和TB1的内源激素含量测定的结果(图5)表明:GA、ZR、IPA、IAA的含量均表现为TA1>WT>TB1; BR含量表现为两个转基因植株高于WT植株,而JA-me含量却同时低于WT; ABA的含量呈现为WT>TB1>TA1。各激素含量在转基因植株和野生型植株间未表现出规律性的对应关系。

图5 转CmSAMDC基因拟南芥植株激素含量(鲜重)的变化
3 讨论
植物在长期进化中,常通过调控细胞内基因表达水平来调节胁迫应答保护机制。SAMDC是Spd和Spm合成的关键基因,它涉及到众多植物对非生物胁迫和生物胁迫的抗性反应过程,在植物生长发育、代谢调控及胁迫响应中均具有重要的作用[13-15]。Hazarika研究表明[4],SAMDC基因广泛涉及冷害、高温及盐胁迫等的抗性反应,且一些转SAMDC基因的植物表现出了对外界胁迫的广谱抗性。Roy and Wu[16]利用六倍体小麦的SAMDC基因进行水稻转化,发现转基因水稻在盐胁迫下仍能正常生长发育,其内源亚精胺和精胺含量增加达3~4倍。Franceschetti等[17]将拟南芥的SAMDC基因转入烟草,发现转基因烟草表现出了多种胁迫抗性。Waie and Rajam[18]用人的SAMDC基因转化烟草,发现转基因烟草对干旱和盐胁迫表现出更高的抗性。Peremarti等研究还认为, spd和spm在抵御干旱胁迫中可能起着清除自由基的作用[19]。本研究也发现,在100或150 mmol/L NaCl胁迫下,转CmSAMDC基因拟南芥植株的根长和侧根数均多于野生型植株;在对4周龄转基因植株幼苗浇灌不同浓度的盐溶液时,发现转基因植株较野生型植株具有更强的耐盐性。
在植物受到胁迫后,多胺在次生代谢调控及信号转导中起着重要的作用[20]。本研究发现,转基因拟南芥植株在盐胁迫下的MDA含量较野生型低,说明转CmSAMDC基因拟南芥植株在受到外界胁迫时,为了减少活性氧积累造成的伤害,会激发多种信号物质,如多胺,协调各种活性氧清除系统,达到体内活性氧的代谢平衡。但是,CmSAMDC基因和多胺类物质是如何参与植株的耐盐性调控还不得而知。今后有必要对多胺代谢调控网络,以及由其直接或间接调控的潜在抗性基因或物质进行进一步探讨。
