少花蒺藜草在中国北方地区的不同种群遗传多样性分析
2019-10-10宋振张衍雷付卫东张瑞海张国良
宋振,张衍雷,付卫东,张瑞海,张国良
中国农业科学院农业环境与可持续发展研究所,北京 100081
少花蒺藜草在分类学上属于禾本科(Gramineae)、蒺藜草属(Cenchrussp.),原产于北美洲及热带沿海地区(Matocha et al.,2010;周立业等,2014)。少花蒺藜草可以作为一种牧草,然而其刺苞表面密被许多小刺,因此会对人及牲畜造成伤害,影响农牧生产。少花蒺藜草适应性强,具有耐旱、抗寒、耐贫瘠和繁殖能力强等特点,尤其在沙漠地带也可以生长繁衍,对沙漠化治理和生态恢复建设具有重要的意义;同时其适应性强、繁殖扩散快的特点也会使其入侵到农田和牧场造成危害,成为一种很难根绝的杂草(张婷等,2017;张衍雷等,2015)。少花蒺藜草大约在20世纪40年代后期入侵到中国,但其入侵原因尚不确定,有学者认为是在二战期间由日本人在东北地区垦殖时人为带入,还有观点认为少花蒺藜草是在中国北方进行牧草引种时随货物流入(安瑞军等,2015;齐凤林等,2011)。少花蒺藜草入侵后迅速繁殖蔓延,目前在中国北方农牧交错带干旱半干旱沙地分布广泛。很多研究人员发现,在此地区的蒺藜草属杂草根据表型主要分为两种:一种直立生长(直立型),一种匍匐生长(匍匐型)。国内学者对这两种类似的蒺藜草属植物是否为同一物种观点不一,甚至对少花蒺藜草称谓和拉丁名也存在争议,有人认为是同一物种的不同表型变异,也有人认为为同一属的两个不同种(安瑞军,2013;徐军等,2011)。《中国植物志》(中国科学院中国植物志委员会,1990)等权威文献中关于蒺藜草属植物形态学的具体细节描述认为,入侵中国北方地区的两种表型蒺藜草属植物分别为少花蒺藜草(C.spinifex)和蒺藜草(C.echinatus),是不同的两个种。两种植物的表型差异主要有以下两点:(1)蒺藜草植株无分蘖或分蘖较少,表现为直立生长,而少花蒺藜草植株分蘖数量较多,表现为匍匐生长;(2)蒺藜草刺苞上刚毛轮状着生,刺苞背部具较密的细毛和长绵毛,而少花蒺藜草刺苞几乎无毛。但是表型的差异可能是由于环境因素等条件造成的,并不能完全确定二者是否为相同的种。
入侵生物的种类和数量具有显著的地理区域性和时间动态性,而同一物种在不同环境条件下的生长也会产生较大的表型可塑性差异(齐艳红等,2004;黄成成等,2018)。研究入侵植物的不同地理种群的种类和进化特征的手段多种多样,除了常规利用形态、解剖结构和生理生化方法外,分子生物学手段也发挥着重要作用。近年来,DNA条形码(DNA barcode)技术在植物分类鉴定中的应用越来越广泛(宁淑萍等,2008;任保青等,2010)。DNA条形码是指生物体内能够代表该物种的、标准的、有足够变异的、易扩增且相对较短的DNA片段,目前最常用的是ITS、rbcL、trnH-psbA等基因序列。这些序列具有长度高度保守性等特点,可以较好的用来进行生物种群的分类研究(Baldwin,1992;Group,2009;Kress et al.,2007;Wang et al.,2010)。另外一种常用的方法是随机扩增多态性DNA标记法(Random Amplified Polymorphic DNA,RAPD)(Lynch et al.,2010;Powell et al.,1996)。该方法于上世纪90年代初诞生,是建立在PCR基础之上的一种可对整个未知序列的基因组进行多态性分析的分子技术,现已广泛的应用于生物的品种鉴定、系谱分析及进化关系的研究上。利用上述分子生物学手段来进行生物遗传多样性、生态型进化等分析是一种可靠快速的方法,尤其对入侵物种的扩散途径和种群分类进化等方面的研究具有重要的应用意义。
目前对中国北方地区分布的不同地理种群少花蒺藜草的入侵、扩散和进化等研究报道较少,尤其两种不同表型的蒺藜草属植物的种群差异和进化分类学研究尚未见报道。本研究利用ITS、rbcL、trnH-psbA等序列保守和变异的特点对中国北方地区两种表型的蒺藜草属植物进行进化差异分析,并结合其生物学特性来确定其物种分类学地位。同时采用RAPD技术对来源于中国北方不同地理区域的少花蒺藜草种群遗传多样性进行了分析,初步摸索了少花蒺藜草在北方农牧交错带干旱半干旱地区的扩散传播路径及种群遗传多样性特点,以期为揭示少花蒺藜草入侵机制、防治少花蒺藜草的危害提供理论依据。
1 材料和方法
1.1 材料

图1 不同地理种群采样点分布图 Fig. 1 Distribution map of sampling points of different geographical populations
在中国北方农牧交错带地区选取蒺藜草属杂草(包括直立型和匍匐型两种表型)入侵较为严重的16个地理种群进行取样,收集这些样本的成熟种子及植物叶片。具体地理种群和命名见图1所示,16个取样点具体信息为:内蒙古赤峰市敖汉旗(AH-A,42°28.935′N,119°20.353′E;AH-B,41°99.307′N,120°07.978′E;AH-C,42°47.007′N,120°52.357′E)、辽宁省朝阳市(CY,41°32.050′N,120°27.301′E)、辽宁省锦州市凌海县(LH,41°18.541′N,121°27.301′E)、辽宁省锦州市易县(YX,41°32.878′N,121°14.651′E)、辽宁省阜新市彰武县(ZW,42°48.454′N,121°58.608′E)、内蒙古通辽市科尔沁区大林镇(TL-Ⅰ,43°43.691′N,122°41.191′E)通辽市科尔沁左翼后旗金宝屯镇(TL-Ⅱ,43°22.596′N,123°29.339′E)通辽市科尔沁左翼后旗(TL-Ⅲ,43°00.867′N,122°25.094′E)、内蒙古通辽市库伦旗澳伦新村(TL-Ⅳ,42°48.454′N,121°58.608′E)、内蒙古通辽市科尔沁左翼中旗小努 日 木村(TL-Ⅴ,43°96.938′N,123°10.286′E)、内蒙古通辽市科尔沁左翼中旗花吐古拉镇(TL-Ⅵ,43°86.697′N,122°8.384′E)、内蒙古通辽市四合 屯 牧场(TL-Ⅶ,43°39.292′N,122°03.710′E)、内蒙古通辽市开鲁县大泡子村(TL-Ⅷ,43°31.583′N,121°28.904′E)、内蒙古通辽市奈曼旗八仙筒镇(TL-Ⅸ,43°11.857′N,121°15.469′E)。扩增DNA条形码保守序列及RAPD分析的材料取自两种不同表型的植物叶片组织。
1.2 方法
1.2.1 DNA的提取
将采集的种子播种于光照培养箱,温度设定白天30 ℃,夜间20 ℃,光照周期16 h。取出苗后生长14 d的幼苗叶片组织,采用改良CTAB方法提取植物样本总DNA(Stewart,1993;王涛等,2017)。
利用ND5000超微量紫外分光光度计(Beijing BioTeke Corporation)测定DNA纯度和浓度,将DNA提取液稀释至终浓度30 ng·μL-1待用。
1.2.2 DNA条形码分析
采用文献报道的方法分别扩增ITS、rbcL、trnH-psbA基因序列(Baldwin,1992;Group,2009;Kress et al.,2007;Wang et al.,2010)。
ITS扩增通用引物ITS5(上游引物,5′GGAA GTAA AAGT CGTA ACAAGG3′)和ITS4(下游引物,5′TCCT CCGC TTAT TGAT ATGC3′)。PCR扩增反应体系(25 μL):10×Buffer(含2 mM Mg2+)12.5 μL,正反引物各1 μL,0.12 mM dNTPs 2 μL,Taq DNA聚合酶(5 U·μL-1)1 μL,DNA模板1 μL,ddH2O 7μL。反应程序如下:预变性95 ℃,5 min;变性94 ℃,1 min;复性54 ℃,1 min;延伸72 ℃,1 min;共30个循环,最后72 ℃延伸10 min。
rbcL引物序列为:1F:ATGTCACCACAAAC AGAAAC,724R:TCGCATGTACCTGCAGTAGC,25 μL反应体系:10×Buffer(含2 mM Mg2+)12.5 μL,正反引物各1 μL,0.12 mM dNTPs 2 μL,Taq DNA 聚合酶(5 U·μL-1)1 μL,DNA模板1 μL,ddH2O 7 μL。反应程序如下:预变性95 ℃,4 min;变性94 ℃,30 s;复性53 ℃,1 min;延伸72 ℃,1 min;共35个循环,最后72 ℃延伸10 min。
trnH-psbA引物序列为:psbA(F):CTTATGCAT GAACGTAATGCTC,trnH2(R):CGCGCGCATGGT GATTCACAATCC。25 μL反应体系:10×Buffer(含2 mM Mg2+)12.5 μL,正反引物各1 μL,0.12 mM dNTPs 2 μL,Taq DNA 聚合酶(5 U·μL-1)1 μL,DNA模板1 μL,ddH2O 7μL。反应程序如下:预变性95 ℃,4 min;变性94 ℃,30 s;复性53 ℃,1 min;延伸72 ℃,1 min;共35个循环,最后72 ℃延伸10 min。
1.2.3 RAPD分析
RAPD随机引物购自北京鼎国昌盛生物技术有限责任公司。利用这些引物进行RAPD扩增,对条带清晰试验进行至少3次的重复实验。
PCR反应体系:总体积为25 μL,包括10×Buffer(含2 mM Mg2+)2.5 μL,0.2 mM引物1 μL,0.12 mM dNTPs 1μL,1 U Taq DNA聚合酶,DNA模板1 μL。PCR反应程序:95 ℃预变性5 min;94 ℃变性45 s,36 ℃复性45 s,72 ℃延伸90 s,35个循环;最后,72 ℃延伸5 min。每次PCR反应均设不含模板DNA的空白对照。扩增产物以1.5%琼脂糖凝胶电泳分离,Goldview染色,在凝胶成像系统下观察并拍照记录。
1.2.4 数据处理
数据处理主要参照文献所述方法(Nei et al.,1979),将RAPD电泳谱带位点上有扩增位点的记为1,无扩增位点的记为0,建立原始数据矩阵,用Pop Gene 32、NTsys 2.1软件进行数据统计分析。主要数据指标为:群体的多态位点百分率P=扩增的多态位点数/扩增的总位点数×100%、Nei’s基因多样性(He)、Shannon’s多态性信息指数(H0)、Nei’s遗传一致度(I)和遗传距离(D)的参数分析;最后用NTsys 2.1软件进行基于遗传距离的聚类分析,构建亲缘进化遗传图谱。
2 结果与分析
2.1 两种不同表型蒺藜草属植物的分类学鉴定
入侵中国北方地区的蒺藜草属植物分布着许多地理生态群,主要包括两种表型:直立型和匍匐型。这两种表型的蒺藜草属植物外形上存在较大的区别,直立型相比匍匐型具有较大的叶片、茎直立、株高较高和一级分蘖数高,但是单株种子量和二级分蘖数低(图2)。通过DNA条形码保守序列差异分析进一步确定这两种不同表型的植物在分类学上的地位。3种保守序列引物PCR扩增的结果如图3显示,两种表型的蒺藜草属植物均得到单一的特异扩增条带。对扩增的条带进行测序,将测序结果与NCBI数据库中相关序列进行比对(图4)。其中,两种表型的蒺藜草属植物的ITS序列比对结果均与Khan S报道的蒺藜草(C.echinatus)[NCBI登录号Sequence ID: gb|KF850611.1|]关系最近,序列相似度 均为99%。而两种蒺藜草属的保守序列之间相似度也为99%。因此笔者综合判断得出,入侵中国北方地区的两种表型蒺藜草属植物均为少花蒺藜草(C.spinifex)。造成两种表型差异的原因可能与光照、温度、水分等不同环境因素的长期影响有关。对于少花蒺藜草的拉丁名,目前也有多种表述。笔者根据文献资料,并沿用目前国际上较为通用的说法(Simon,2010;张婷等,2017),建议统一使用C.spinifex作为少花蒺藜草的拉丁名。

图2 少花蒺藜草的两种不同表型差异 Fig. 2 Differences between the two phenotypic types of C. spinifex

图3 两种表型的蒺藜草属植物DNA条形码保守序列扩增电泳图 Fig. 3 Amplified electrophoresis of DNA barcode conservative sequences of the two phenotypic types

图4 两种表型的植物保守序列在NCBI的比对结果 Fig. 4 Comparison of DNA barcode sequences of the two phenotypic types in NCBI
2.2 入侵中国北方地区不同地理种群少花蒺藜草的进化关系分析
对200条RAPD随机引物进行扩增筛选,其中有12条引物能够扩增出清楚、多态性明显的条带,获得扩增产物长度为200-2000 bp的条带37个,其中具有多态性的有35个。扩增结果表明不同引物所扩增出的谱带数不同,不同材料之间的扩增带数存在多态性。对12条引物的扩增结果进行统计,结果见表1,多态性条带比率(P)为94.59%,每条引物的多态性位点在2-4个之间,平均每个引物扩增出3.08个位点。各条带均有较大的Nei’s遗传多样性指数及Shannon’s多态性信息指数,反应了各个样点之间存在丰富的多态性。
采用Nei’s遗传一致度(I)和遗传距离(D)对入侵中国北方地区不同少花蒺藜草种群遗传进化程度进行分析,结果见表2。16个地理种群中,样点TL-Ⅵ与的样点TL-Ⅷ遗传一致度最高,遗传距离最近(I=0.8919,D=0.1144),亲缘关系近;而样点TL-Ⅵ与样点TL-Ⅲ、样点AH-A与样点YX及样点YX与样点TL-Ⅱ 3组的遗传距离远,亲缘关系稍远。样点AH-C与其他各样点遗传距离均较远。

表1 12条随机引物的扩增结果(5′-3′) Table 1 The amplification results of 12 random primers (5′-3′)
为了进一步确立这16个地理种群的亲缘进化关系,基于Nei’s遗传距离来构建UPGMA聚类图谱(图5)。当遗传相似度阈值为0.6时,可将16个地理种群划分成5个类,即内蒙古赤峰市敖汉旗西部地区、通辽市奈曼旗、通辽市左翼中旗样本;通辽市库伦旗样本;通辽市科尔沁左翼后旗、辽宁省朝阳市、彰武县样本;凌海市、易县样本;以及赤峰市敖汉旗北部地区。当遗传距离阈值为0.79时,可基本将所有地理种群分开。
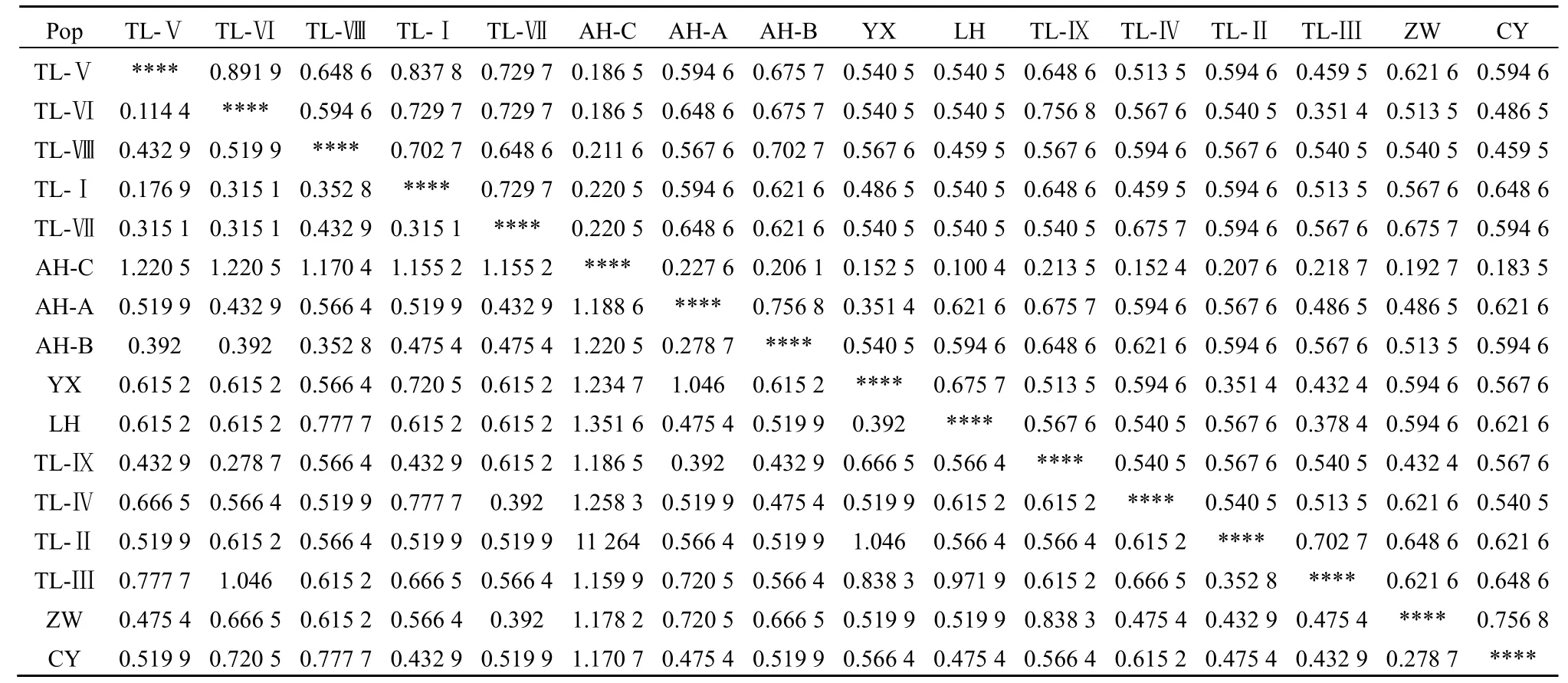
表2 种群遗传一致度和遗传距离 Table 2 Nei's original measures of genetic identity and genetic distance

图5 中国北方地区不同地理种群的少花蒺藜草UPMGA聚类图 Fig. 5 UPGMA dendrogram for the 16 population based on Nei’s genetic distance
通过上面的分析,我们可以初步判断出中国北方地区少花蒺藜草的入侵路线。研究结果表明,赤峰市敖汉旗所取的3个样点少花蒺藜草入侵途径不同。南部地区(AH-B)少花蒺藜草与辽宁省朝阳市的样本亲缘关系较近(D=0.4754),北部地区(AH-A)与通辽市奈曼旗的样本亲缘较近(D=0.4329)。辽宁省凌海市与易县两地的少花蒺藜草亲缘关系较近(D=0.392),而易县的样本又与彰武县的样本亲缘关系较近(D=0.5199),同时,彰武县与通辽市左翼后旗样点间的遗传距离也相对较小,通辽市境内的各个取样点之间的关系也可以根据遗传距离明确。因此我们判断,入侵内蒙古、辽宁地区的少花蒺藜草传播扩散路径有两条主线,一条是凌海-易县-彰武-通辽市左翼后旗—通辽市左翼中旗,同时彰武地区的少花蒺藜草又扩散到朝阳市、通辽市左翼后旗地区扩散到库伦旗;另一条为通辽市左翼中旗-开鲁县-奈曼旗-赤峰市敖汉旗,同时库伦旗的少花蒺藜草也向敖汉旗方向扩散。
3 讨论
3.1 入侵中国北方地区的蒺藜草属植物分类学地位
对于入侵中国北方地区的少花蒺藜草的分类地位观点不一,争论不休。如徐军等(2011)认为,入侵地的蒺藜草应为光梗蒺藜草(C.incertus);而安瑞军(2013)认为,入侵地的少花蒺藜草为一个新种,拉丁名应为C.pauciflorus。两篇报道皆在总结前人文献,仅根据表型特征得出的推论,这种分类学混乱的局面不利于对少花蒺藜草的入侵机制的研究及防治技术的开发。因此,本研究根据入侵中国北方的两种不同表型植物的形态学特征,结合《中国植物志》的描述,初步判断其均应为蒺藜草属(Cenchrussp.)植物,又利用更为精确的分子生物学方法,通过对两种表型的蒺藜草属植物(直立型和匍匐型)的ITS等保守序列进行扩增测序和比较分析,判定这两种表型的蒺藜草属植物均为少花蒺藜草(C.spinifex)。根据笔者所在团队多年的调查研究结果显示,匍匐型少花蒺藜草主要在中国北方农牧交错带干旱半干旱沙地分布,而直立型少花蒺藜草在中国分布范围较广,从东北地区、东部沿海,直到海南岛均有分布。这可能与不同环境条件下表型的变异有关,这也与《中国植物志》中关于两种表型植物的分布规律描述一致(中国科学院中国植物志委员会,1990)。
3.2 中国北方地区少花蒺藜草的地理种群遗传多样性及传播扩散路径分析
利用RAPD技术能够有效地对一些物种进行遗传多样性分析,这种分子生物学手段从DNA序列变异和进化水平来分析生物种群之间的进化地位差异,所获得的结果较为客观和真实,该方法可广泛应用于动物、植物和微生物的分类鉴定研究(Jones et al.,2015;Lynch et al.,2010;Powell et al.,1996;Shiue et al.,2015;Stewart et al.,1993)。目前,国内已有较多报道利用RAPD技术分析研究不同粮食作物及经济作物的遗传多样性(林新春等,2003;肖猛等,2015;徐影等,2013;翟亚辉等,2013),但是对于入侵植物少花蒺藜草的分类及进化地位方面还未见报道。本实验通过RAPD技术对入侵中国北方地区的少花蒺藜草不同地理种群进行比对研究,发现其具有丰富的遗传多样性特点。
少花蒺藜草种子可通过农具、动物携带、水流等方式迅速向外扩散传播,加上其自身所具有的耐旱、抗寒、耐贫瘠及繁殖能力强等特点,在中国北方地区形成了不同的地理种群。对这些地理种群的遗传变异和进化进行研究,可以揭示少花蒺藜草在中国北方地区的入侵路线,有利于对其入侵、传播和危害的遏制(张衍雷等,2015;张志新等,2015)。本研究结果初步描绘了中国北方地区入侵少花蒺藜草的入侵传播路线。从传播路径图上可以看出,少花蒺藜草的主要扩散方式是跨区域车辆运输活动,如凌海-左翼中旗路线主要是沿G101和G304国道进行,敖汉旗-左翼中旗路线主要沿G111国道进行。所以,加强地区间的检疫工作是防止少花蒺藜草扩散蔓延的重要手段。另外一种人为扩散方式是少花蒺藜草种子随货物夹杂或随牧草引种传播。本研究发现,凌海市的少花蒺藜草可能是内蒙古、辽宁地区少花蒺藜草传播的源头。由于凌海市紧邻锦州市,不排除少花蒺藜草种子可能是随进出口货物或牧草引种而带入的,随后向其他各地传播。然而,本研究并未对锦州地区及其他周边地区的少花蒺藜草进行分析,也未采集国外少花蒺藜草的生物样本,所以具体推论还仍需进一步试验验证。
4 结论
本研究结合形态学特征分析,采用DNA条形码技术,研究了中国北方农牧交错带地区两种表型的蒺藜草属杂草的分类学地位,通过保守序列测序和分析比对,确定了两种不同表型(直立型、匍匐型)植物均为少花蒺藜草(C.spinifex)。采用RAPD技术对来源于不同地理区域的少花蒺藜草种群遗传多样性进行了分析,构建了亲缘进化遗传图谱,发现入侵中国东北三省及内蒙古地区的少花蒺藜草沿不同国道方向的传播主线主要有两条:一条是凌海-易县-彰武-通辽市左翼后旗-通辽市左翼中旗,同时彰武地区的少花蒺藜草又扩散到朝阳市、通辽市左翼后旗地区扩散到库伦旗;另一条为通辽市左翼中旗-开鲁县-奈曼旗-赤峰市敖汉旗,同时库伦旗的少花蒺藜草也向敖汉旗方向扩散。研究结果为进一步揭示少花蒺藜草入侵的生态学机制奠定了理论依据。
