广西贝江大型底栖动物群落结构时空分布特征
2019-09-17赵学敏马千里姚玲爱梁荣昌
赵 瑞,赵学敏,苟 婷,马千里,姚玲爱,梁荣昌
生态环境部华南环境科学研究所,广州 510655
大型底栖动物是河流生态系统的重要组成部分,位于食物链的关键环节,在物质循环和能量流动中扮演重要角色,通过取食加速碎屑分解,促进水体自净,在维持生态系统平衡中发挥着重要作用[1-2]。同时,底栖动物因生命周期长,活动缓慢,分布范围广,生物多样性较高,对环境变化比较敏感,能够较好地反映过去一段时间内水体受干扰和胁迫的累积效应,已被广泛应用于水质监测的国家标准中[3-4],是评价河流健康最为重要的类群之一。欧盟水框架协议(European Water Framework Directive,WFD)早在2000年就提出水生态健康评价时除了要综合考虑水环境质量,水生生物群落结构与分布特征也是评价的关键部分[5]。大型底栖动物的很多评价指数是依据物种出现与否或者群落结构特征来指示河流水质的优劣。如果对污染敏感的物种出现,表明该河流水质良好;当水质恶化时,敏感物种会被耐污能力更强的物种替代。因此,大型底栖动物群落结构的差异可以准确地反映出河流水体受污染的性质和程度以及人类干扰对水生态系统的影响[6-8]。
贝江流域位于广西中北部的柳州融水苗族自治县境内,是柳江主流融江的支流之一,发源于黔桂交界的九万大山。唐鑫等[9]和苟婷等[10]分别对贝江的硅藻和浮游藻类进行了相关生态研究,有关底栖动物的研究工作相对薄弱。此外,在2012年龙江河镉污染事件之后,柳州的水源安全问题日益突显[11],贝江被柳州市政府选作备用水源。因此,在贝江流域开展基于大型底栖动物的生态调查,分析底栖动物在季节和空间上的群落结构变化特征,揭示大型底栖动物群落演变规律,探讨水利工程对水生态系统的干扰,并通过底栖生物评价指数对贝江水环境状况进行评估,以期为贝江流域水环境管理和河流生态系统保护修复提供理论依据和数据支持。
1 实验部分
1.1 研究区域
贝江是柳江流域融江的支流之一,流经融水县汪洞、三防、怀宝、四荣、融水等乡镇,于融水县江门附近汇入融江,流域地理位置为108°43′~109°16′E,25°07′~25°27′N。贝江干流范围为融水县三防镇至贝江河口,干流流域集水面积为1 790 km2,河长96 km,河流落差为150 m,平均坡降为1.35‰,河道干流坡降为10.0%[12]。因河流落差大,蕴藏丰富水能,贝江目前已建成拉川和江门电站,鱼窝电站和落久电站正在建设中。
为了研究贝江流域底栖动物群落结构特征,根据贝江的流域形态、地形地貌特征和电站分布特点,研究沿贝江流域从上游兰马采样点(B1)至下游江门坝下采样点(B17)共设置17个采样点,其中贝江干流布设15个采样点,贝江2条主要支流(都郎河和香粉河)各布设1个采样点,基本覆盖贝江整个流域各种生境状况(图1)。贝江干流拉川电站采样点(B3)和江门电站采样点(B16)属于库区河段,其余采样点为自然河段。
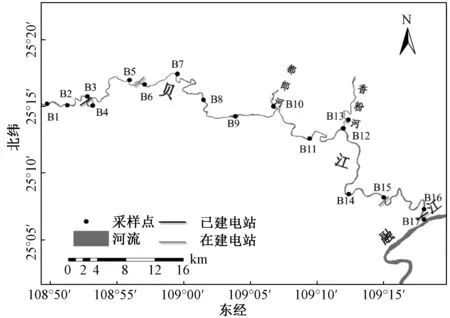
注:B1.兰马;B2.三防镇;B3.拉川坝上;B4拉川坝下;B5.鱼窝坝上;B6.鱼窝坝下;B7.河村口桥;B8.怀宝镇;B9.喷滩底;B10.田头河;B11.马安底;B12.四荣乡;B13.香粉河;B14.榄口码头;B15.落久坝址;B16.江门坝上;B17.江门坝下。图1 贝江流域大型底栖动物采样点分布Fig.1 Sampling sites of macroinvertebrate in the Beijiang River basin
1.2 样品采集
在可涉水区域,利用索伯网(0.3 m×0.3 m,孔径为0.5 mm),选择不同生境类型的采样断面收集3个平行样品;在不可涉水区域,于近岸处用开口面积为1/16 m2的改良型彼得森采泥器采集3个平行样品。收集后将样品冲洗并用0.25 mm筛网过滤,放入500 mL广口瓶,加75%酒精保存。在实验室条件下,利用体式显微镜(OlympusSZ61)将绝大多数物种鉴定到种或属。并参照BARBOUR等[4]的分类方法划分底栖动物的功能摄食类群。
1.3 数据分析
1.3.1 生物多样性评价指数
包括Shannon-Weiner多样性指数(H′)[13]和Margalef丰富度指数(D)[14]。
D=(S-1)/lnN
式中:N为群落中所有物种的个体总数;ni为第i个物种的个体数;S为总物种数。H′水质评价标准为>3,清洁;>2~3,轻污染;>1~2,中污染;>0~1,重污染。
1.3.2 水质生物学评价指数
包括Shannon-Weiner多样性指数(H′)和Hilsenhoff 生物指数(BI)[15]
式中:N为群落中所有物种的个体总数;ni为第i个物种的个体数;ti为第i个物种耐污值。BI水质评价标准为<5.5,清洁;5.5~<7.5,轻污染;7.5~<8.5,中污染;8.5~<10,重污染。
1.3.3 优势度分析
Y=fi×(ni/N)
式中:Y为优势度;N为群落中所有物种的个体总数;fi为第i种底栖动物在各采样点出现的频度,当Y≥0.02时,物种为优势种。
1.3.4 统计分析
大型底栖动物群落结构分析采用多种非参数变量分析方法,包括单因子ANOSIM分析和SIMPER分析,以上分析均由Primer V5.2.6软件完成。
同时运用Pcord软件中的非度量多维尺度转换排序(NMDS)进行采样点群落组成分析,检验不同季节底栖动物群落结构的差异性。采用相似性矩阵分析各采样点群落组成的相似性,该矩阵的构建基础为Bray-Curtis相似性系数,并采用多响应置换过程(MRPP)法检验各采样点间群落组成的差异性。
2 结果与分析
2.1 群落组成及优势种
2015年冬季(1月)和夏季(8月)2次调查共采获大型底栖动物2 132头,隶属于3门8纲20目52科146属(种),其中昆虫纲为优势类群,为9目45科127属(种),占总分类单元的86.99%,昆虫纲主要由双翅目、蜻蜓目和蜉蝣目组成,分别占昆虫纲分类单元数的36.22%、24.41%和16.54%;软体动物门为3纲4目6科9属(种),占6.16%;寡毛纲为2目2科5属(种),占3.42%;蛭纲为3目3科3属(种),占2.05%,最少为甲壳纲,1属(种),占1.36%(图2和表1)。
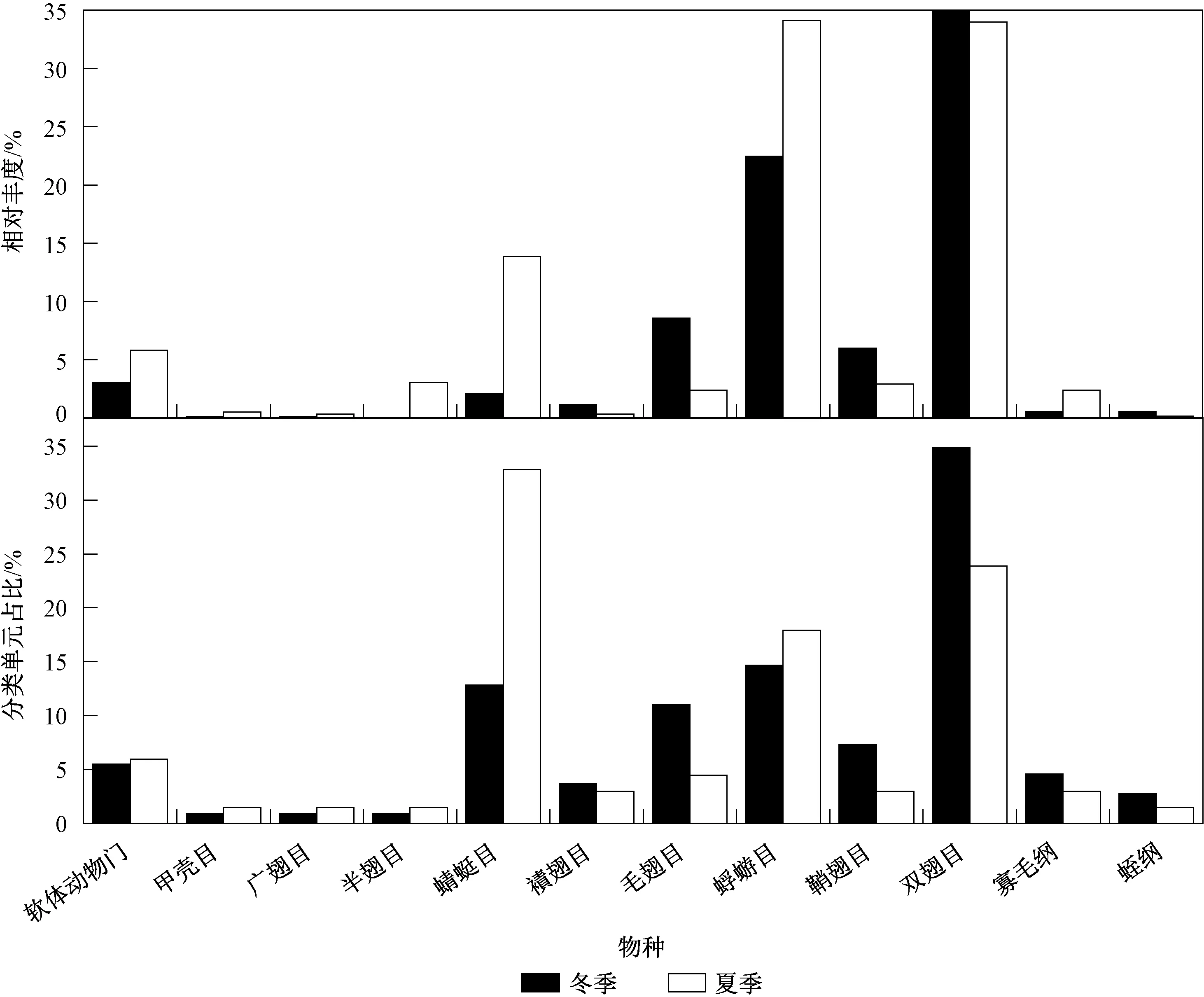
图2 贝江流域2015年冬季和夏季底栖动物群落结构组成Fig.2 Macroinvertebrate community structure in winter and summer of 2015 in the Beijiang River basin

分类单元冬季夏季分类单元冬季夏季软体动物门MolluscaMiniperla sp.+腹足纲GastropodaPerlomyia sp.++铜锈环棱螺Bellamya aeruginosa+Gibosia sp.+方格短沟蜷Semisulcospira cancellata+Perlomyica sp.小土蜗Galba pervia+毛翅目 Trichoptera萝卜螺属Radix sp.+侧枝纹石蛾属Ceratopsyche sp.+椭圆萝卜螺Radix swinhoei+纹石蛾属Hydropsyche sp.+折叠萝卜螺Radix plicatula+短脉石蛾属Chumatopsyche sp.++大脐圆扁螺Hippeutis umibilicalis+Plectrocnemia spp.+双壳纲Bivalvia姬石蛾属Hydroptila sp.+淡水壳菜Limnoperna lacustris+Helicopsyche sp.++河蚬Corbicula fluminea++舌石蛾属Glossosoma sp.+节肢动物门Arthropoda Triplectidae sp.甲壳纲 Crustacea直角弯突石蛾Setodes sp.+沼虾属Macrobrachium sp.++角石蛾属Stenopsyche sp.+昆虫纲Insecta石蛾科 Phryganeidae广翅目Megaloptera等翅石蛾科 Philopotamidae中华班鱼蛉Neochauliodes sinensis++黑毛剑石蛾属Melanotrichia sp.蜻蜓目Odonata蜉蝣目 Ephemeroptera黑山蟌属Philosina sp.++七鳃假二翅蜉Pseudocloeon morum++箭蜓科 Gomphidae sp.+原二翅蜉属Procloeon sp.+鼻蟌属Rhinocypha sp.四节蜉属Baetis sp.++纤春蜓属Leptogomphus sp.+花翅蜉属Baetiella sp.+歧角纤春蜓Leptogomphus divaricatus+生米蜉Ephemera shengmi++安春蜓属Amphigomphus sp.+徐氏蜉Ephemera hsui+邵春蜓属Shaogomphus sp.+白背锯形蜉Serratella albostriata++深山闵春蜓Fukionogonphus prometheus+小蜉属Ephmerella sp.+凶猛春蜓Labrogomphus sp.+扁蜉科1种 Heptageniidae sp.小叶春蜓Gomphidia sp.+扁蜉属Heptagenia sp.+扭尾曦春蜓Helingomphus retroflexus+高翔蜉属Epeorus sp.+闽春蜓属Fukionogonphus sp.+似动蜉属Cinygmina sp.++日春蜓属Nihonogomphus sp.+红斑似动蜉Cinygmina rubromacu+显春蜓属Phaenandrogom phus sp.+河花蜉属Patomanthus sp.++亚春蜓属Asiagomphus sp.+宽基蜉属Choroterpes sp.++索氏缅春蜓Burmagomphus sowerbyi+拟细裳蜉属Paraleptophlebia sp.+尖尾春蜓Stylogomphus tantulus++带肋蜉属Cincticostella sp.+弯尾春蜓Melligomphus sp.+锐利蜉属Ephacerella sp.+广东弯尾春蜓Melligomphus guangdognensis++天角蜉属Uracanthella sp.+海南弯尾春蜓Lamelligomphus hainanensis+小蜉科 Ephemerellidae sp.驼峰环尾春蜓Lamelligomphus camels+细蜉属Caenis sp.++双峰环尾春蜓Melligomphus ardens+半翅目Hemiptera佛鲁戴春蜓Davidius fruhstorferi+潜水蝽科Gelastocoridae sp.+双角戴春蜓Davidius bicornutus+鞘翅目 Coleoptera+长节阔腹春蜓Stylurus amicus+Elmid sp.++扩腹春蜓属Stylurus sp1.+Ordobrevia sp.+长足春蜓属Merogomphus sp.+Pseudamophilus sp.+内春蜓属Nepogomphus sp.+Sinopsephenoides sp.++弯钩弓蜻Macromia unca+Eubrianax sp.+红胭蜻Rhodothemis rufa+Psephenus sp.+彩虹蜻Zygonyx iris+Senocolus sp.+襀翅目Plecoptera双翅目 Diptera石蝇科 Perlidae sp.+大蚊属第一种Tipula sp1.++大蚊属第二种Tipula sp2.+下凸长跗摇蚊Tanytarsus chinyensis+Baeoura sp.+秃特维摇蚊Tvetenia calvescens+朝大蚊属第一种Antocha sp1.++弯松施密摇蚊属Krenosmittia sp.+朝大蚊属第二种Antocha sp2.+溪流直突摇蚊Othocladius rivulorum++

表1(续)Table 1 (continued)
冬季共采获并鉴定110个大型底栖动物分类单元,其中昆虫纲为主要类群,占比为85.45%,主要由双翅目、蜉蝣目和蜻蜓目构成,分别占昆虫纲分类单元数的40.43%、18.09%和14.89%;软体动物为6种,寡毛纲为5种,蛭纲为3种,甲壳纲为1种。夏季共采获并鉴定70个大型底栖动物分类单元,昆虫纲占比为88.57%,主要由蜻蜓目、蜉蝣目和双翅目组成,分别占35.48%、19.35%和29.03%;软体动物为4种,寡毛纲为2种,甲壳纲和蛭纲各1种。
与冬季相比,夏季底栖动物总分类单元数减少,但2个季节的优势类群均为昆虫纲,并且昆虫纲主要组成类群类似,都包括蜻蜓目、双翅目和蜉蝣目。
此外,大型底栖动物的总密度也呈现冬季(17 211.11 ind./m2)高于夏季(6 477.78 ind./m2)的现象,并且主要密度贡献类群组成不同,冬季双翅目密度最高,占比为55.3%,其次为蜉蝣目(22.5%)和毛翅目(8.6%);夏季则主要为蜉蝣目(34.1%)和双翅目(33.9%),其次为蜻蜓目(13.9%)。同时,独立样本非参数K-S检验也表明,各采样点的底栖动物密度也呈现极显著的季节性差异(P<0.01)。
该区域优势物种季节差异明显,不同季节出现的优势物种不同,同一优势种在不同季节优势度不同。冬季的优势物种为直突摇蚊属Orthocladiussp.(Y=0.07),朝大蚊属一种Antochasp1.(Y=0.04),生米蜉Ephemerashengmi(Y=0.05)和河花蜉属Patomanthussp.(Y=0.03),优势度大于0.001的物种为31种;夏季的优势物种为四节蜉属Baetissp.(Y=0.04),河花蜉属Patomanthussp.(Y=0.04)和异带小突长跗摇蚊Micropsectraatrofasciata(Y=0.03),优势度大于0.001的物种为25种。其中河花蜉属为2个季节共有物种,但优势度为夏季高于冬季。
2.2 群落空间分布特征及指示种分析
贝江流域大型底栖动物群落结构特征指数H′和D空间差异明显,上游河段(包含采样点B1、B2、B4~B6)底栖动物H′和D值最高,其次为中游河段(包含采样点B8~B14),相对较低的采样点出现在下游河段(B15和B17)和库区河段(B3和B16)。
对比贝江冬季与夏季底栖动物的H′和D值,发现大部分上游和中游河段冬季指数值高于夏季,下游河段和库区河段则相反(图3)。
基于底栖动物群落结构的相似性,贝江流域各采样点可以聚成2组,组1包括B1、B2、B4、B5、B7、B8、B10、B11、B12、B13和B14,采样点分布在贝江干流上游、中游卵石底质河段以及贝江的2条支流。组2包括B3、B6、B9、B15、B16和B17,其中采样点B3和B16分别属于贝江上游拉川电站库区和下游江门电站库区,河段水深,流速慢,底质多为淤泥;B15和B17采样点位于下游落久坝址和江门电站下游流速慢,底质为淤泥的河段,采样点B6和B9属于岩石底质。经MRPP检验表明,各组间差异显著(P<0.01)。组1内相似性较高,平均距离为0.75;组2内相似性较差,平均距离为0.94(图4)。
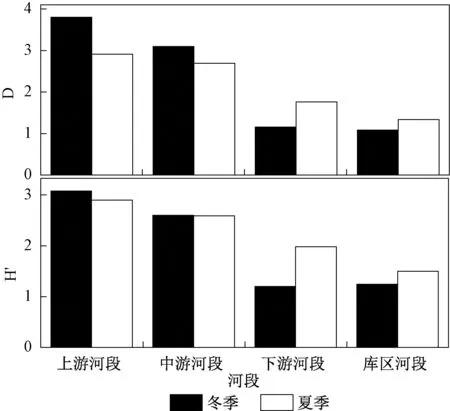
图3 贝江流域大型底栖动物H′和D空间分布Fig.3 Spatial distribution of macroinvertebrates Shannon-Wiener index and species richness in the Beijiang River basin
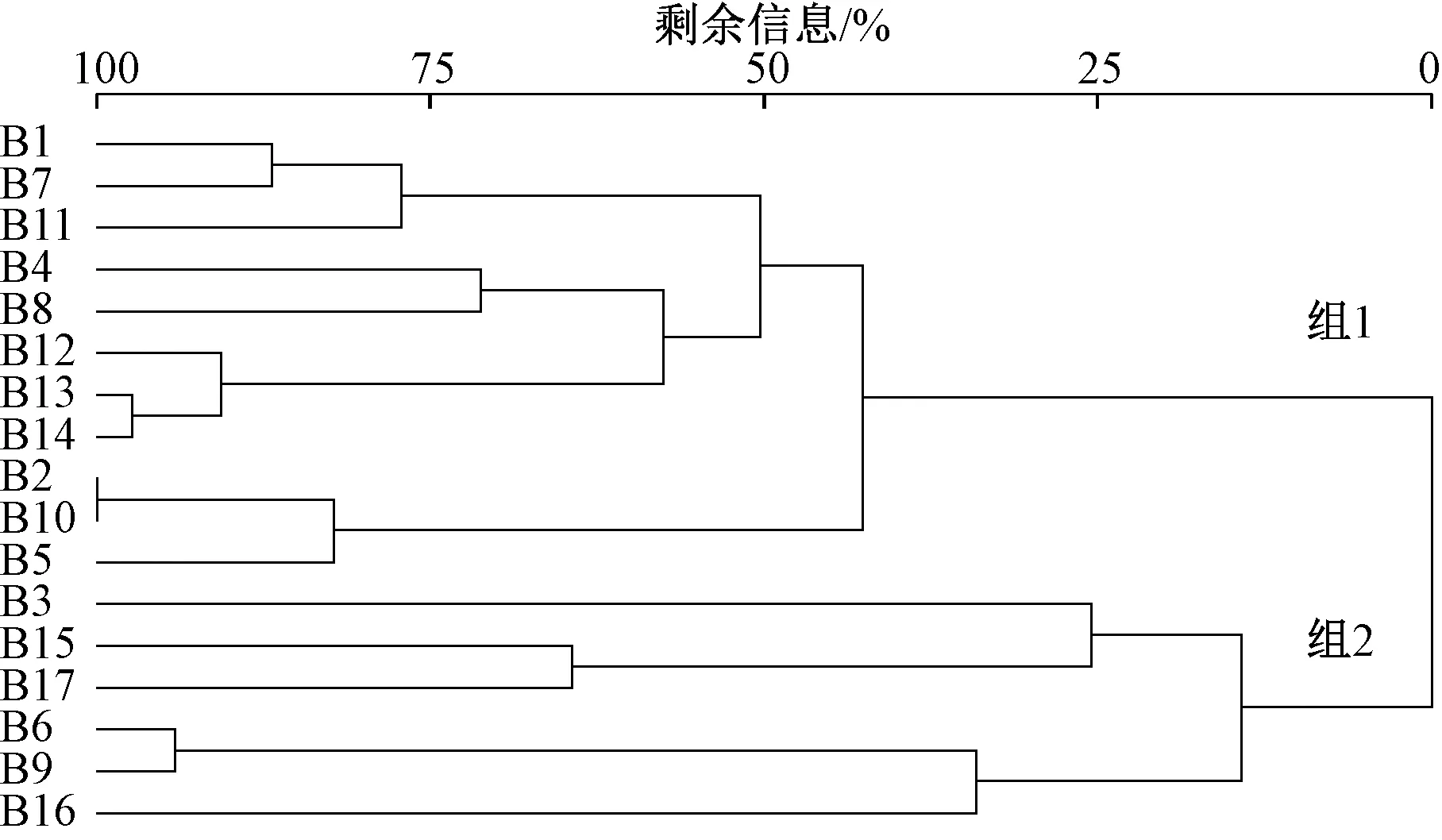
图4 贝江流域大型底栖动物聚类分析结果Fig.4 Cluster analysis for macroinvertebrate assemblage in the Beijiang River basin
不同分组底栖动物的分类单元数和密度差异显著,组1共有99个分类单元数,总密度为12 911.11 ind./m2,组2共计26个分类单元数,总密度为8 556 ind./m2。不同分组的优势类群和指示种各异,组1的优势类群为蜉蝣目、襀翅目和毛翅目,三者共占30%,指示种种类丰富,主要为营沙石底生活的水生昆虫和软体动物,并且以清洁敏感物种为主。组2优势类群为双翅目,占到50%,指示种均为摇蚊幼虫,充分体现出贝江电站附近上下游河流生境组成较为单一(表2)。贝江各采样点底栖动物群落结构相似性分析表明,该流域底栖动物群落结构空间差异显著。基于底栖动物物种组成的各采样点NMDS排序分析(图5),结合ANOSIM分析发现,贝江流域冬季和夏季底栖动物群落组成存在极显著差异(r=0.26,P<0.001)。通过SIMPER分析得到造成2个季节大型底栖动物群落结构差异的主要贡献物种为河花蜉属Patomanthussp.、朝大蚊属一种Antochasp1.、生米蜉Ephemerashengmi、宽基蜉Choroterpessp.、四节蜉Baetissp.、异带小突长跗摇蚊Micropsectraatrofasciata、溪流直突摇蚊Othocladiusrivulorum、无突摇蚊Ablabesmyiasp.、喙隐摇蚊Cryptochiromomussp.和扩腹春蜓属Stylurussp1.,这些物种共解释了2个季节50%的底栖动物群落结构组成的差异。
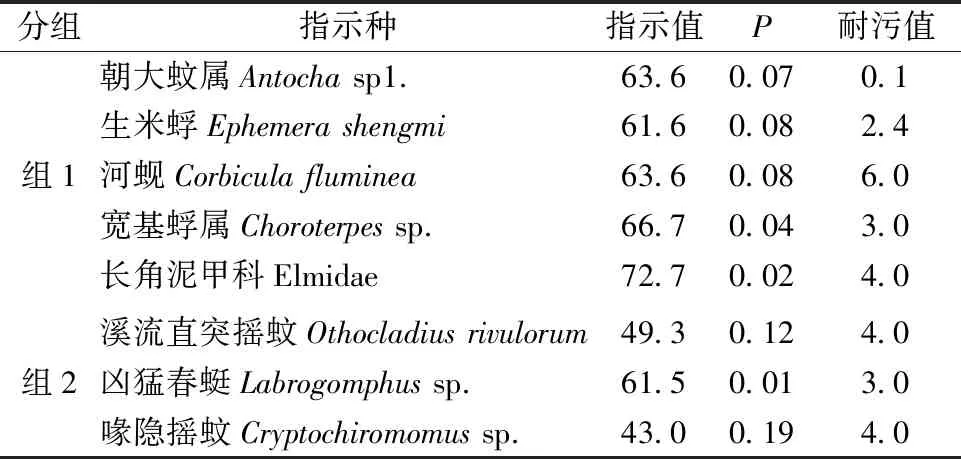
表2 贝江流域大型底栖动物指示种Table 2 Indicator species in the Beijiang River basin
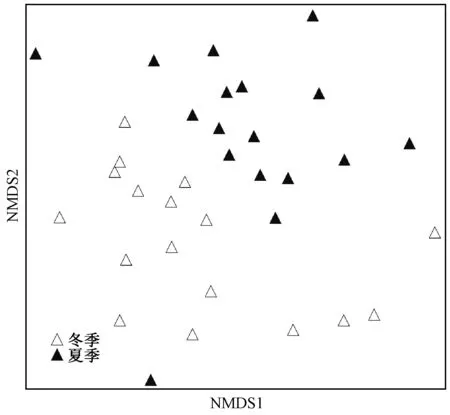
图5 基于物种多度的冬季和夏季底栖动物NMDS采样点排序Fig.5 Nonparametric multidimensional scaling (NMDS) ordination of sampling sites based on macroinvertebrates abundance in winter and summer
2.3 功能摄食类群特征
贝江大型底栖动物在2个季节的摄食功能群分析结果显示,该流域底栖动物中以集食者和撕食者为主要摄食功能群,个体数比例分别为55%和16%,其次为滤食者(14%)和捕食者(11%),刮食者所占比例较小。贝江流域不同季节底栖动物摄食功能群组成整体相似,冬季底栖动物优势类群为集食者(58%)和撕食者(16%),夏季也为集食者(47%)和撕食者(18%),但夏季撕食者和捕食者个体数比例有所升高。
贝江流域不同采样点底栖动物摄食功能群组成存在空间差异,冬季摄食功能群B1和B2,B4~B14基本以集食者和撕食者为主要类群,两者个体数之和占总数60%以上,B3、B15~B17以滤食者为主。夏季B1和B2,B4~B14摄食功能群组成均以撕食者、集食者和捕食者为主,B3,B15~B17则以捕食者、滤食者和刮食者为主,具体信息如图6所示。

图6 贝江流域不同季节大型底栖动物摄食功能群Fig.6 Functional feeding groups of the macroinvertebrate community in different seasons of the Beijiang River basin
2.4 水质生物学评价
整体看,2种水质评价指数对于贝江水质的评价呈现H′比BI评价结果略差(表3)。

表3 各采样点生物指数及水质生物学评价Table 3 Biotic indexes and bioassessment of water quality in different stations
贝江各采样点H′评价结果表明,贝江有76.5%的采样点属于清洁-轻污,其余采样点属于中污-重污;BI评价则显示贝江整个流域水质清洁。
从贝江水质评价结果的采样点分布情况看,H′与BI评价结果相对较吻合,中上游区域采样点的水质基本为清洁-轻污。
3 讨论
3.1 底栖动物群落结构变化
大量研究表明,季节差异是底栖动物不同群落的生活史特征与生境条件协同作用的结果[16-19]。贝江底栖动物群落结构存在显著的季节变化,物种分类单元数和丰度均表现为冬季>夏季。昆虫纲作为主要贡献类群,其双翅目、蜉蝣目、毛翅目和蜻蜓目的种类数和相对丰度均呈现冬季高于夏季,说明贝江冬季的生境条件(如水温、流速、营养物循环等)可能更适于这些水生昆虫类群的生长和繁殖,这与香溪河[16]底栖动物季节变化特点一致。贝江夏季正值华南区的雨季,暴雨引发的洪水可能是导致该季节底栖动物丰度和种类数大量减少的原因之一[20]。另外,相关研究表明底栖动物群落结构还会受到温度变化的影响[21-23],贝江夏季底栖动物种类数和丰度较低可能与水温升高导致适应低温生存的水生昆虫数量减少有关。
底栖动物对于生境的变化比较敏感,而人为干扰(如梯级电站的修建)会使河流的水文特征和底质条件发生改变,由原来的河流生态系统演变为水库生态系统,导致底栖动物密度降低,群落特征随时间的推移向静水群落特征演替[24-26]。贝江流域底栖动物群落结构相似性分析发现,河段上未修建电站的采样点聚成1组,这些采样点主要分布在贝江上游、中游和支流的自然河段,水位浅,水流速度相对较快,底质类型为卵石,底栖动物物种组成相对丰富,以附石性适应流水生活的水生昆虫(如蜉蝣目、毛翅目、襀翅目和鞘翅目等)类群为主;而另1组采样点主要分布在贝江梯级电站(拉川、江门电站)和下游河段附近,底质类型以淤泥、细沙为主,水位较深,水流较缓,底栖动物类群以摇蚊科幼虫和水丝蚓类等静水类群为主。贝江具有山地溪流的典型特征,即坡降大,水流湍急,溶氧充足,水质清洁,底质多为卵石和漂砾[27],是水质敏感类群蜉蝣目、毛翅目和襀翅目水生昆虫的最适生境[28]。因此,在贝江自然河段区域,蜉蝣目、毛翅目和襀翅目成为优势类群,与同属山地溪流的鱼肚河[29]和香溪河[25]类似。而位于贝江上游区域的拉川电站采样点和下游的江门电站,因修建电站,导致原来的山地溪流生态系统转变为库区生态系统,水流变缓,底质为泥沙底,为摇蚊科幼虫和水丝蚓提供了适宜的生存环境。
3.2 功能摄食类群分析
VANNOTE等[30]在河流连续统概念中提出,对于山地溪流,河岸带凋落物为溪流生态系统的主要能量来源,大型底栖动物一般以撕食者和集食者为主,撕食者主要以河道底部的树叶及河岸带植被为食[31-32],集食者则主要取食有机碎屑。贝江属于山地性溪流,河流两岸植被密布,并且2个季节的底栖动物摄食功能群组成均以集食者和撕食者为主要类群,这与VANNOTE等[30]和王强等[29]的研究相符。
3.3 贝江水质生物学评价
研究采用H′和BI对贝江流域水质状况进行评价,评价结果存在差异,H′评价发现贝江多数采样点属于清洁-轻污,BI评价表明贝江流域整体水质清洁。两类指数评价结果中,BI评价结果与贝江流域实际水体理化指标状况最符合[10]。这是由于生物多样性指数属于生物群落结构性指数,是基于物种分类单元数,同时综合考虑物种数量的组成对水质状况进行评价,但未考虑底栖群落中敏感和耐污类群组成差异对于评价结果准确性的贡献[33]。而BI是基于底栖动物耐污值和物种多样性建立的,能够较准确地反映出河流水质状况,但耐污值的地域性差别较大,目前国内已有王备新等[34]和赵瑞等[35]分别建立了适于中国东南部和东北地区河流的底栖动物耐污值体系,而华南地区底栖动物的耐污值研究尚不足,研究中BI评价目前是参考中国东部地区耐污值,后续研究将对其进行补充完善。
4 结论
1)贝江底栖动物群落结构存在季节性差异,群落数量特征和结构复杂性均表现为冬季>夏季。
2)贝江流域底栖动物群落结构分布存在明显的空间差异,流域中自然河流采样点物种丰度高于梯级电站采样点,并且自然河流采样点的优势类群以营流水卵石底生活的蜉蝣目、襀翅目和毛翅目物种为主,而电站采样点则以喜静水的双翅目和寡毛纲为主。
3)底栖动物功能摄食类群以集食者和撕食者为主,其次为滤食者和捕食者,刮食者所占比例较小。
4)水质评价结果显示贝江流域水质整体处于较清洁状态。
