野生荞麦细柄野荞麦的繁殖生物学特性研究
2019-09-10周兵闫小红苏启陶张争光王宁陈丽花
周兵 闫小红 苏启陶 张争光 王宁 陈丽花



摘 要: 該研究通过野外观察和人工控制实验相结合的方法,从开花动态、花部基本特征、繁育系统、传粉生物学及种子性状等方面对荞麦属(FagopyrumMill.)植物细柄野荞麦(Fagopyrum gracilipes)的繁殖生物学特性进行了探究,并分析了各性状对其繁殖的贡献。结果表明:在贵州威宁,细柄野荞麦的花果期常为每年的6—10月,单花序和单花的花期分别为13~21 d和1~3 d。花较小,直径为(3.99 ± 0.12)mm,花柱和花药高分别为1.30和1.65 mm,花直径与花被片长和花被片宽呈显著正相关,花柱高与花药高呈极显著正相关。细柄野荞麦花粉胚珠比为371 ± 16.40,杂交指数为2,套袋实验显示其自交、异交亲和,表明其繁育系统为兼性自交,部分异交亲和。细柄野荞麦的访花昆虫较少,主要为膜翅目(Hymenoptera)、双翅目(Diptera)和鞘翅目(Coleoptera)7个科的9种昆虫,食蚜蝇科(Syrphidae)昆虫是其主要传粉昆虫。细柄野荞麦果实存在有翅和无翅两种类型,有利于其适应不同的传播方式,种子较小,千粒重为(1.05 ± 0.04)g,萌发率较低,播种后30 d的累积萌发率为(19.60 ± 2.14)%,但萌发整齐,主要集中在前5 d。综上所述,细柄野荞麦灵活的繁育系统为其产生大量种子提供了保障,多样的果实传播方式和整齐的种子萌发特性为其占据更广阔的生境成为群落优势种创造了基础。
关键词: 细柄野荞麦, 繁育系统, 传粉生物学, 种子, 繁殖
中图分类号: Q944.3, Q945.5
文献标识码: A
文章编号: 1000-3142(2019)05-0590-10
Reproductive biological characteristics ofwild Fagopyrum gracilipes
ZHOU Bing*, YAN Xiaohong, SU Qitao, ZHANG Zhengguang, WANG Ning, CHEN Lihua
( School of Life Sciences, Jinggangshan University, Ji’an 343009, Jiangxi, China)
Abstract: We investigated reproductive traits of Fagopyrumgracilipessuch as flowering dynamics, floral basic characteristics, breeding system, pollination biology and seed characteristics by field observations and artificial control experiments, also discussed their contribution to the reproduction of F. gracilipes. The results showed that F. gracilipes flo-wered and fruited during June to Octomber in Weining, Guizhou Province. The flowering lifespans of a single inflorescence and flower were about 13-21 d and 1-3 d respectively. The flower was small with the diameter of (3.99 ± 0.12)mm, the height of the style and anther was 1.30 and 1.65 mm respectively, the floral diameter had significant positive correlation with tepal length and tepal width (P< 0.05), the height of the style had highly significant position correlative with the height of anther (P< 0.01). The pollen-ovule ratio of F. gracilipeswas 371 ± 16.40, its out-crossing index was 2, and the results of bagging experiments showed it was self- and cross-compatible, which suggested that the bree-ding system of F. gracilipeswas facultative selfing, partial crossing-compatible. The main floral visitors were nine insects of Hymenoptera, Diptera and Coleoptera, the insects in family Syrphidae were its main pollination insects. The fruits of F. gracilipesincluded two types with and without wings which was helpful for its adaption to different spread manners, its seeds were smaller with the thousand seed weight of (1.05 ± 0.04) g, and had low but orderly germination rate with accumulated germination rate of (19.60 ± 2.14)% 30 d after sowing. It could be concluded that the flexible breeding system provided security for mass seeds production of F. gracilipes, various fruit spread manners and orderly seed germination characteristics created the basis for its colonizing vast habitats and becoming the dominant species in the community. The results can provide important reference for revealing the reproduction mechanism of F. gracilipesand buckwheat breeding.
Key words: Fagopyrum gracilipes,breeding system,pollination biology,seed, reproduction
繁殖成功是植物种群赖以延续和扩展的关键,是植物对其生存环境长期适应进化的结果。因此,一切与植物繁殖有关的特征及过程均是植物繁殖生物学的研究内容,包括开花生物学、繁育系统、传粉生物学、繁殖构件和分配等(张大勇和姜新华,2001;Barrett,2002;Huang,2006;任明迅等,2012)。虽然不同植物间的繁殖过程及细节存在差异,但最终植物常通过提高种子的数量与质量来获得较高的雌性适合度,或通过提高花粉获得量和效率来获得较高的雄性适合度(任明迅等,2012)。目前,繁殖生物学研究主要集中在造成濒危植物濒危的繁殖生物学特性(Pellegrino et al., 2006;Cursach & Rita,2012;莫权辉等,2016)、繁殖生物学特性与外来入侵植物入侵性的关系(周兵等,2013;Castro et al.,2013;Yan et al.,2016)以及特殊花部性状的繁殖生态学功能(文慧慧等,2015;Suetsugu,2015;Wang et al.,2016)等方面,而针对经济植物,尤其是经济植物的近缘野生种的繁殖生物学特性研究相对较少(张万灵等,2013)。实际上,揭示这些近缘野生种的繁殖生物学特性将为选育优良品种及性状基因提供重要理论基础。
中国是世界荞麦的起源中心,在西南地区,荞麦属(FagopyrumMill.)几乎所有种类均有分布。荞麦因具有较高的营养价值和保健药用功能而备受关注和广泛种植(阮景军和陈惠,2008;唐文等,2010)。该属植物分为大粒组和小粒组两个类群,两组间的种间杂交亲和性非常低,组内种间杂交亲和性相对较高(Chen,1999;Chen et al.,2004)。该属植物的资源学(吴振海等,2010;王安虎等,2011)、形态学(陈庆富,2004;任翠娟和陈庆富,2008;Tang et al.,2010)、遗传学(Zhou et al.,2012;Kishore et al.,2013;Li et al.,2013)以及开发应用(Park et al.,2012;Tsai et al.,2012)等方面已有研究,而关于该属植物的繁殖生物学方面的研究相对较少(张万灵等,2013;Chen et al.,2015;Wu et al.,2018)。细柄野荞麦(Fagopyrum gracilipes)為荞麦属小粒组一年生草本植物,主要分布在我国西南较高海拔、较冷地区,常生长于草坡、湿山谷、田边或与玉米等作物混生(陈庆富,2012)。笔者调查发现在分布区,尤其是作物田中,细柄野荞麦很容易形成优势种群。细柄野荞麦没有营养生殖,那么其优势种群的形成是否与其有性生殖存在密切联系,尤其是哪些有性繁殖特性发挥重要作用有待探索。为此,本研究从花部基本特征、繁育系统、访花昆虫、种子生物学特性等方面对细柄野荞麦的繁殖生物学特性开展研究,从而为揭示其繁殖机制及形成优势种群的原因提供相关理论依据,同时也为荞麦育种提供理论参考。
1 材料与方法
1.1 样地基本情况
样地设在贵州省威宁彝族回族苗族自治县草海,样地总面积约200 m2。该地属亚热带季风气候,年均降雨量约为750 mm,年均气温为11.2 ℃,年均日照数为1 812 h,地理坐标介于104°12′—104°18′ E,26°49′—26°53′ N之间,生物多样性高,物种丰富,是细柄野荞麦的理想生存地。
1.2 研究方法
1.2.1 开花动态及花部基本特征观测 用记号笔标记待开放的细柄野荞麦花蕾,每天进行观察直至开放,且开放当天对开放的花朵进行连续观察,并对花被片、雄蕊、雌蕊各花部构件的变化过程及现象进行记录。随机标记待开放的花序,连续进行观察统计,直至最后1朵花凋谢,并计算其开放时间。随机选取20朵正盛开的花朵,用游标卡尺测量花柄长、花直径、花被片长和宽、花药与花柱高度等花部形态指标。
1.2.2 繁育系统测定
1.2.2.1 花粉胚珠比 随机选取20朵即将开放但未散粉的花,收集其所有花药于1.5 mL离心管中,加入0.5 mL 1.0 mol·L-1 HCl溶液去除花药壁,振荡形成悬浮液。用移液枪移取10 μL悬浮液于载玻片上,显微镜下统计全部花粉量,统计5次,取平均值,计算单花花粉量。用刀片横切所统计花的子房,显微镜下统计胚珠数。计算花粉胚珠比(P/O)。按Cruden(1977)的标准评价其繁育系统。
1.2.2.2 杂交指数估算 根据细柄野荞麦花的直径及雌雄蕊特性,参考Dafni(1992)的标准来评价其繁育系统。具体依据如下:(1)花朵或花序直径<1 mm、1~2 mm、2~6 mm及>6 mm时分别记为0、1、2、3;(2)花药开裂时间与柱头可授期同时或柱头可授期更早记为0,花药先开裂记为1;(3)柱头与花药在同一高度记为0,存在高度差记为1。三者之和为杂交指数(out crossing index, OCI)值。根据OCI值评价其繁育系统:OCI=0,闭花受精;OCI=1,专性自交;OCI=2,兼性自交;OCI=3,自交亲和,有时需要传粉者;OCI=4,部分自交亲和,异交,需要传粉者。
1.2.2.3 套袋实验 在细柄野荞麦花期,随机选择植株若干,每株选择发育一致的花做如下处理:(1)不做任何处理,对照;(2)不去雄,开花前套袋;(3)开花前去雄,不套袋;(4)开花前去雄,授异株异花花粉;(5)开花前去雄,不授粉,套袋。每株做处理2~4组,之后定期记录处理情况,统计结实率。
1.2.3 访花昆虫观测 在细柄野荞麦的盛花期对其访花昆虫进行观察,观察时间为晴天的每天8:30—18:00,不少于3 d。主要观察访花昆虫的访花行为,并进行拍照,同时用捕虫网捕捉相关访花昆虫,带回实验室做成标本,进行鉴定。
1.2.4 花期构件特性测定 于细柄野荞麦盛花期,在样地中随机选择30株植株,小心将整个植株挖取,统计其叶片数、花序数及不同级别的分枝数,测量植株高度和根长。
1.2.5 种子的基本特征及萌发特性测定 取50粒籽粒饱满的细柄野荞麦种子,测量其高度、宽度、竖长、短长等基本特征。随机挑选5组籽粒饱满的细柄野荞麦种子各100粒,称重,计算其千粒重。随机选择籽粒饱满的细柄野荞麦种子若干,放入水中浸泡5 h后,置于鋪有湿润滤纸的培养皿中进行萌发实验,每皿50粒,设4个重复。置于培养箱中进行培养[培养湿度为70%、温度为(25±1)℃],每天记录种子萌发情况,连续统计30 d,计算萌发率。
1.2.6 统计分析 所有数据采用Excel和SPSS19.0统计分析软件进行处理,采用Pearson指数双侧检验法对花部各构件的相关性进行分析。
2 结果与分析
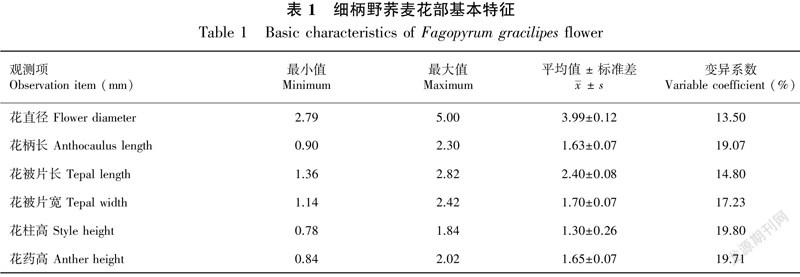
2.1 细柄野荞麦的开花动态及花部基本特征
2.1.1 开花动态 细柄野荞麦在贵州威宁的花果期常为每年的6—10月份,7—8月份为其盛花期。其花序为总状花序,单花序内有花10~40朵不等,每苞片内有花2~3朵,花较小。花被片5枚,深裂,淡红色到白色;雄蕊8枚,短于花被片,内3外5,花柱3,3心皮1心室。单花序的花期为13~21 d,单花花期一般为1~3 d,在花开放过程中,花序下面苞片中的其中1朵花先开,上面苞片中的花随后依次开放,而花序下端内其它的花也会同时开放;花被片由开始的包裹状态到微张再完全张开,颜色由淡红色变为白色最后变成褐色,花被片伸展的同时花丝和花柱也伸长,花药和柱头近等高,内轮花药先散粉,2 h后外轮花药开始散粉,雄蕊先于花被片枯萎;花无香味,开放时有少量分泌物产生。
2.1.2 花部基本特征 表1显示了细柄野荞麦花部各构件的基本特征,存在一定的差异。花柄长、花柱高和花药高的变异相对较大,平均值分别为1.63、1.30、1.65 mm,变异系数均大于19%。花直径和花被片长的变异相对较小,相应的变异系数均小于15%,平均值分别为3.99、2.40 mm。相关性分析表明,细柄野荞麦花部各构件形态指标间存在一定相关性。其中,花直径与花被片长(r=0.462)呈显著正相关,与花被片宽(r=0.555)呈极显著正相关;花柄长与花柱高(r=0.528)呈显著正相关,与花药高(r=0.637)呈极显著正相关;花被片长与花被片宽(r=0.447)呈显著正相关;花柱高与花药高(r=0.693)呈极显著正相关(表2)。
2.2 细柄野荞麦的繁育系统
2.2.1 花粉胚珠比 细柄野荞麦单花的花粉量为270~580粒,平均单花花粉量为(371 ± 16.40)粒,雌蕊为3心皮1心室,有胚珠1枚,可见其花粉胚珠比(P/O)为371 ± 16.40。根据Cruden(1977)的标准,细柄野荞麦的繁育系统偏向为兼性自交。
2.2.2 杂交指数 表3显示了细柄野荞麦的杂交指数测量结果。细柄野荞麦花朵直径为3.99 mm,记为2。柱头在花药散粉前即有可授性,即雌蕊先熟,记为0。柱头和花药近等高,记为0。统计可得细柄野荞麦杂交指数(OCI)为2,按照Dafni(1992)的评价标准可见其繁育系统为兼性自交。
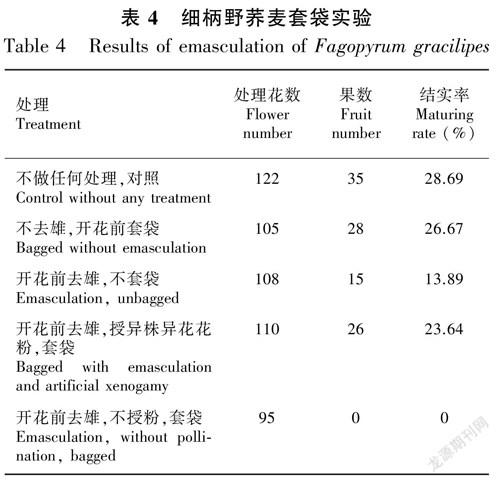
2.2.3 套袋实验 表4显示了细柄野荞套袋和人工授粉实验结果。开花前去雄,不授粉,套袋处理时细柄野荞麦的结实率为0,表明其不存在无融合生殖现象;自然条件下其结实率为28.69%,不去雄,开花前套袋的结实率为26.67%,两者显示了较近的结实率,表明细柄野荞具有较高的自交亲和性;开花前去雄,不套袋及开花前去雄,授异株异花花粉,套袋两个处理其结实率分别为13.89%和23.64%,表明细柄野荞也是异交亲和的。可见,细柄野荞的繁育系统是以自交为主的兼性自交。
2.3 细柄野荞麦的访花昆虫
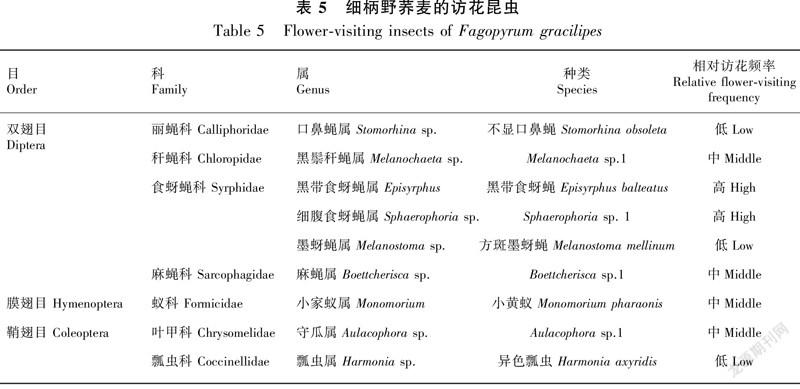
晴天的早晨8:30左右,在细柄野荞麦上开始有访花昆虫,但访花昆虫相对较少。经过观察发现,细柄野荞麦的访花昆虫包括双翅目(Diptera)、膜翅目(Hymenoptera)和鞘翅目(Coleoptera)昆虫近10种(表5和图1)。主要访花昆虫有食蚜蝇科(Syrphidae)的黑带食蚜蝇(Episyrphus balteatus)和细腹食蚜蝇(Sphaerophoriasp. 1)、麻蝇科(Sarcophagidae)的麻蝇(Boettcheriscasp.1)以及叶甲科(Chrysomelidae)的守瓜(Aulacophorasp.1)。
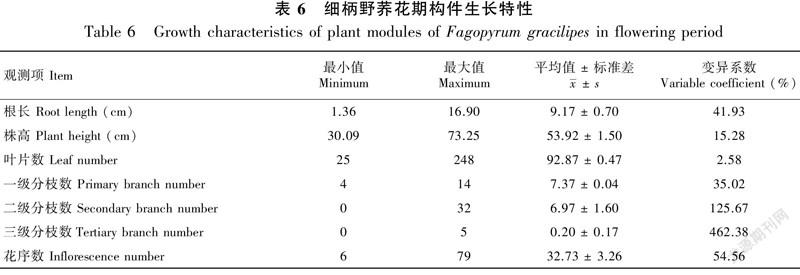
2.4 细柄野荞麦花期构件生长特性
表6显示了花期细柄野荞麦构件的生长特性。可见,细柄野荞麦构件特性在植株个体间存在一定的差异。其叶片数和株高相对稳定,叶片数为25~248片,株高为30.09~73.25 cm,平均值分别为92.87片和53.92 cm,变异系数分别为2.58%和15.28%;可形成三级分枝,一级分枝数相对稳定,三级分枝不稳定,两者的变异系数分别为35.02%和462.38%;其花序数为6~79个,平均为32.73个,变异系数为54.56%。
2.5 细柄野荞麦种子的基本特征及萌发特性
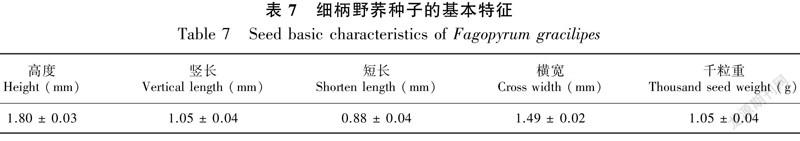
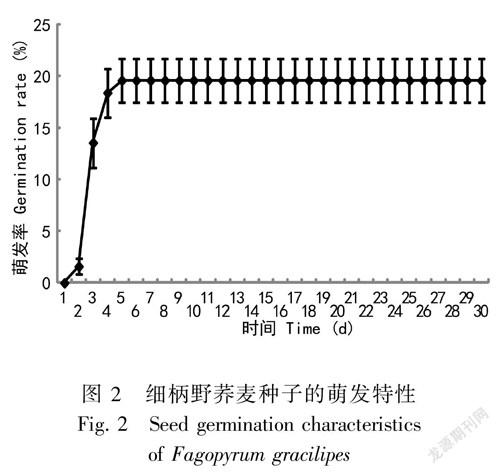
细柄野荞麦的种子较小,其高度、竖长、短长和横宽分别为1.80、1.05、0.88和1.49 mm,千粒重为1.05 g,相对较轻(表7)。由图2可知,在湿度70%,温度(25±1)℃条件下,细柄野荞麦种子的萌发率较低,30 d其累积萌发率仅为19.60%,但其萌发比较整齐,主要集中在第2天至第5天。
3 讨论
植物繁殖成功是植物繁育系统和传粉生物学的综合作用的结果(Harmon-Threatt et al.,2009)。Ramírez & Nassar(2017)认为被子植物繁育系统包括无融合生殖、自发自花授粉、辅助性自花授粉以及自花不亲和四种类型, 是遗传信息与环境因素共同作用的结果。植物的繁育系统并不是一成不变的,随着生存环境的变化可能会存在着繁育系统的地理变异性。比如任明迅等(2004)研究表明,虽然克隆繁殖是凤眼蓝(Eichhornia crassipes) 的主要更新方式,中国西南和海南岛种群表现出较多样化的繁育系统,而其它地理种群,由于部分花型的缺失,其异交系统解体,倾向于自交亲和类型。这种差异在酢酱草科(Oxalidaceae)植物高山酢酱草(Oxalis alpina)的不同地理种群间同样存在(Weller et al.,2016)。植物的繁育系统通常通过花粉胚珠比、杂交指数及套袋实验等来进行检测。本研究结果表明,细柄野荞麦花的花粉胚珠比为371,杂交指数为2,根据相关的评价标准其繁育系统为兼性自交,而套袋实验的结果表明其是自交异交亲和的。可见,细柄野荞麦的繁育系统为自交为主的兼性自交。植物繁育系统的类型通常与其花部性状密切相关,比如雌雄异花类型的植物只能是异交亲和的,而两性花的植物会表现出多样化的繁育系统类型(Castro et al.,2004;Costa et al.,2013)。在荞麦属植物中,部分植物存在二型花柱,这种花部性状使得其具有较严格的自交不亲和特性(Yasui et al.,1998;张万灵等,2013)。该属植物还存在同型花柱植物,这类植物尤其是小粒组的荞麦属植物更多的倾向于兼性自交(Yasui et al.,1998)。该属植物至少经历了3次从二型花柱到同型花柱的转变,Cymosum组两次,Urophyllum组一次即硬枝野荞麦(F. urophyllum)-细柄野荞麦(Yasui et al.,1998;Wu et al.,2018)。这种转变导致繁育系统由原来较为严格的异交转为自交亲和,虽然自交存在一定的负面影响,但自交亲和可有效保障合子的形成,产生大量种子,为植物种群的生存和扩散提供重要保障(Baker,1955)。细柄野荞麦花期构件生长特性表明其单株平均具有37个花序,而每个花序有花10~40朵,这为其产生大量籽实提供了保障,也使得其成为群落内优势种,占据更广阔的生境成为可能。
传粉生物学是植物繁殖生物学研究的另外一个重要方面。访花昆虫的多少尤其是传粉昆虫的多少及其传粉效率对植物的繁殖成功至关重要。Castro et al.(2013)研究表明远志科(Polygalaceae)植物Polygala vayredae的访花昆虫较多,有24种,但只有4种有效传粉昆虫一次访花后即能携带花粉使其沉降在柱头上。这是该物种繁殖产出较低的原因。张万灵等(2013)研究表明金荞麦有访花昆虫38种之多,但由于型内的不亲和性及有效传粉昆虫较少,其结实率为20%左右,而以自交为主的植物其访花昆虫可能更少。本研究表明,细柄野荞麦的访花昆虫较少,只有以膜翅目和双翅目为主的近10种访花昆虫。这种现象在繁育系统为兼性自交的美洲商陆(Phytolacca americana)中得到验证(周兵等,2013)。植物花的结构、颜色、气味、分泌物质类型及其产量等特征对传粉昆虫的吸引产生重要影响(Cerana,2004;Klahre et al.,2011)。获取花粉和花蜜等食物是大多数访花昆虫的访花目的(Heinrich & Raven,1972),而植物常通过产生一定的花部特征吸引或适应某些特定的传粉昆虫(龚燕兵和黄双全,2007)。如凤仙花属(Impatienssp.)植物常通过较长的花筒及距结构的存在使得其传粉昆虫常以具有较长口器的熊蜂及天蛾为主(毛志斌等,2011)。花的大小及花的对称性均使植物对昆虫的吸引能力产生影响。有研究表明,同一类型的花,花越大对昆虫的吸引力越强(Worley et al.,2000),两侧对称的花比辐射对称的花能更有效地传送视觉介导的花部信息,更有利于传粉昆虫的识别(Fenster et al., 2009; Ushimaru et al., 2009)。细柄野荞麦为较小的辐射对称的花,直径仅为3.99 mm,植株较矮小,单花平均花粉量为371粒,为昆虫能提供的食物有限,从而限制了其对昆虫的吸引,而该植物为自交亲和,传粉昆虫的存在与否对其结实影响较小,这也可能是其访花昆虫较少的原因。
植物籽实的性状会直接影响到植物种群的延续和扩散。为适应风力传播苏门白酒草(Conyza sumatrensis)等菊科植物常产生成具有冠毛的瘦果(Hao et al.,2009),为借助动物的携带传播大狼把草(Bidens frondosa)等植物常产生具有钩刺的果实(Yan et al.,2016);而北美车前(Plantago virginica)等植物通过产生具有粘液的种子适应旱生环境(罗辉等,2015)。细柄野荞麦果实含有有翅和无翅两种类型(陈庆富,2012),有翅的类型可借助水流进行传播,而无翅的类型则可以占据原有的生境。植物种子的萌发特性是影响其种群的另一重要方面。细柄野荞麦种子的萌发率较低,在湿度70%、温度(25±1)℃的条件下,其30 d内的累积萌发率不足20%,但其萌发较为整齐,主要集中在前5 d内。在种子量基数大的情况下,将会形成大量新的植株,占据更广阔的生境。美洲商陆种子萌发特性也表现出相类似的结果,其播种20 d后的累积萌发率仅为18.50%(周兵等,2013)。另外,细柄野荞麦千粒重相对較小,为1.05 g,属于小粒种子。虽然小粒种子因含有的有限营养物质而对种子的繁殖力产生负面效应(郭水良等,2006),但其使得该类植物具有更大的多度空间,占据更广阔的生境(刘志民等,2003),同时小粒种子更适应于风力和水力的传播。
可见,细柄野荞麦具有较长的花期、兼性自交,部分异交亲和的灵活繁育系统,种子萌发率较低但萌发整齐且种子量大,同时具有有效的果实传播方式,这些繁殖特性为其占据多样化的生境,成为群落内的优势种奠定了基础。对其繁殖生物学特性的了解,将对揭示其繁殖机制提供理论基础,也为该属栽培荞麦的育种提供理论参考。虽然,本研究对细柄野荞麦的繁殖生物学特性进行了相对较为详尽的研究,但某些方面还有待深入研究。比如细柄野荞麦的开花物候及其花粉粒与柱头的形态学特性等。另外,细柄野荞麦具有有翅和无翅两种类型的果实,那么这两种类型果实间的萌发特性存在什么样的差异?产生这种异型果的相关机制是什么?有待进一步研究。
参考文献:
BAKER HG, 1955. Self compatibility and establishment after“long distance” dispersal [J]. Evolution, 9(3): 1-24.
BARRETT SCH, 2002. The evolution of plant sexual diversity[J]. Nat Rev Genet, 3(4): 274-284.
CASTRO CC, OLIVEIRA PE, ALVES MC, 2004. Breeding system and floral morphometry of distylous PsychotriaL. species in the atlantic rain forest, SE Brazil [J]. Plant Biol, 6: 755-760.
CASTRO S, FERRERO V, COSTA J, et al., 2013. Reproductive strategy of the invasive Oxalis pes-caprae: Distribution patterns of floral morphs, ploidy levels and sexual reproduction[J]. Biol Inv, 15(8): 1863-1875.
CASTRO S, LOUREIRO J, FERRERO V, et al., 2013. So many visitors and so few pollinators: Variation in insect frequency and effectiveness governs the reproductive success of an endemic milkwort [J]. Plant Ecol, 214(10): 1233-1245.
CERANA MM, 2004. Flower morphology and pollination in Mikania(Asteraceae) [J]. Flora, 199: 168-177.
CHEN QF, 1999. Hybridization between Fagopyrum(Polygonaceae) species native to China[J]. Bot J Linn Soc, 131(2): 177-185.
CHEN QF, 2004. A study on structures of abnormal flowers of diploid common buckwheat (Fagopyrum esculentum)[J]. Guihaia, 24(4): 339-341.[陈庆富, 2004. 二倍体普通荞麦异常花朵及其结构的研究[J]. 广西植物, 24(4): 339-341.]
CHEN QF, 2012. Plant sciences on genus Fagopyrum [M]. Beijing: Science Press: 20-22.[陈庆富, 2012. 荞麦属植物科学 [M]. 北京: 科学出版社: 20-22.]
CHEN QF, HSAM SLK, ZELLER FJ, 2004. A study of isozyme, and interspecific hybridization on big-achene group of buckwheat species (Fagopyrum, Polygonaceae)[J]. Crop Sci, 44(5): 1511-1518.
CHEN ML, YOU YL, WEN HH, et al., 2015. The breeding system and reproductive ecology of the endangered plant Fagopyrum dibotrys(D. Don) Hara[J]. Bangladesh J Bot, 43(2): 197-205.
COSTA CBN, COSTA JAS, DE QUEIROZ LP, et al., 2013. Self-compatible sympatric Chamaecrista(Leguminosae-Caesalpinioideae) species present different interspecific isolation mechanisms depending on their phylogenetic proximity [J]. Plant Syst Evol, 299: 699-711.
CRUDEN RW, 1977. Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants[J]. Evolution, 31(1): 32-46.
CURSACH J, RITA J, 2012. Implications of the reproductive biology of the narrow endemic Naufraga balearica(Apiaceae) for its conservation status[J]. Plant Syst Evol, 298(3): 581-596.
DAFNI A, 1992 . Pollination ecology[M]. New York: Oxford University Press: 1-57.
FENSTER CB, ARMBRUSTER WS, WILSON P, et al., 2009. Specialization of flowers: Is floral orientation and overlooked first step? [J] New Phytol, 183: 502-506.
GONG YB, HUANG SQ, 2007. On methodology of foraging behavior of pollinating insects [J]. Biodivers Sci, 15(6): 576-583.[龔燕兵, 黄双全, 2007. 传粉昆虫行为的研究方法探讨[J]. 生物多样性, 15(6): 576-583.]
GUO SL, WANG Y, CAO T, 2006. The high diversity of weed reproduction and adaptation to the human-disturbed environment[J]. J Shanghai Norm Univ (Nat Sci Ed), 35(3): 103-110.[郭水良, 王勇, 曹同, 2006. 杂草繁殖方式的多样性及其对环境的适应[J]. 上海师范大学学报(自然科学版), 35(3): 103-110.]
HAO JH, QIANG S, LIU QQ, et al., 2009. Reproductive traits associated with invasiveness in Conyza sumatrensis [J]. J Syst Evol, 47(3): 245-254.
HARMON-THREATT AN, BURNS JH, SHEMYAKINA LA, et al., 2009. Breeding system and pollination ecology of introduced plants compared to their native relatives[J]. Am J Bot, 96(8): 1544-1550.
HEINRICH B, RAVEN PH, 1972. Energetics and pollination ecology [J]. Science, 176: 597-602.
HUANG SQ, 2006. Debates enrich our understanding of pollination biology[J]. Trends Ecol Evol, 21(5): 233-234.
KISHORE G, PANDEY A, DOBHAL R, et al., 2013. Population genetic study of Fagopyrum tataricumfrom western Himalaya using ISSR markers[J]. Biochem Genet, 51(9-10): 750-765.
KLAHRE U, GURBA A, HERMANN K, et al., 2011. Pollinator choice in Petuniadepends on two major genetic loci for floral scent production[J]. Curr Biol, 21: 730-739.
LI FL, ZELLER FJ, HUANG KF, et al., 2013. Improvement of fluorescent chromosome in situ PCR and its application in the phylogeny of the genus FagopyrumMill. using nuclear genes of chloroplast origin (cpDNA)[J]. Plant Syst Evol, 299(9): 1679-1691.
LIU ZM, JIANG DM, GAO HY, et al., 2003. Relationships between plant reproductive strategy and disturbance[J]. Chin J Appl Ecol, 14(3): 418-422.[劉志民, 蒋德明, 高红瑛, 等, 2003. 植物生活史繁殖对策与干扰关系的研究 [J]. 应用生态学报, 14(3): 418-422.]
LUO H, YAN XH, ZHOU B, et al., 2015. Water absorbance features of seed mucilage of an invasive plant, Plantago virginica, and its germination response to drought stress[J]. Chin J Ecol, 34(8): 2155-2160.[罗辉, 闫小红, 周兵, 等, 2015. 入侵植物北美车前种子粘液的吸水特性及其对干旱胁迫的萌发响应[J]. 生态学杂志, 34(8): 2155-2160.]
MAO ZB, BOEHLER C, GE XJ, 2011. Pollination ecology and breeding system of Impatiens lateristachys(Balsaminaceae) endemic to China[J]. Guihaia, 31(2): 160-166.[毛志斌, Cedric Boehler, 葛学军, 2011. 侧穗凤仙花的传粉生态和繁育系统[J]. 广西植物, 31(2): 160-166.]
MO QH, LI JW, GONG HJ, et al., 2016. Reproductive biology of endangered plant Actinidia chrysantha [J]. Guihaia, 36(6): 640-645.[莫权辉, 李洁维, 龚弘娟, 等, 2016. 濒危植物金花猕猴桃繁殖生物学初步研究[J]. 广西植物, 36(6): 640-645.]
PARK N, LI XH, THWE AA, et al., 2012. Enhancement of rutin in Fagopyrum esculentumhairy root cultures by the Arabidopsistranscription factor AtMYB12[J]. Biotechnol Lett, 34(3): 577-583.
PELLEGRINO G, NOCE ME, BELLUSCI F, et al., 2006. Reproductive biology and conservation genetics of Serapias vomeracea(Orchidaceae) [J]. Folia Geobot, 41(1): 21-32.
RAMREZ N, NASSAR JM, 2017. Breeding systems in angiosperms: Novel inferences from a new analytical approach[J]. Plant Syst Evol, 303(2): 1-19.
REN CJ, CHEN QF, 2008. The current status and prospect of the study on morphological anatomy and phylogeny of Fagopyrum [J]. J Guizhou Educ Inst (Nat Sci Ed), 19(3): 23-27.[任翠娟, 陈庆富, 2008. 荞麦(Fagopyrum)的形态解剖学及系统学研究[J]. 贵州教育学院(自然科学版), 19(3): 23-27.]
REN MX, JIANG XH, ZHANG DY, 2012. Some important questions in plant reproductive ecology[J]. Biodivers Sci, 20(3): 241-249.[任明迅, 姜新华, 张大勇, 2012. 植物繁殖生态学的若干重要问题 [J]. 生物多样性, 20(3): 241-249.]
REN MX, ZHANG QG, ZHANG DY, 2004. Geographical va-riation in the breeding systems of an invasive plant, Eichhornia crassipes, within China[J]. Acta Phytoecol Sin, 28(6): 753-760.[任明迅, 张全国, 张大勇, 2004. 入侵植物凤眼蓝繁育系统在中国境内的地理变异[J]. 植物生态学报, 28(6): 753-760.]
RUAN JJ, CHEN H, 2008. Buchwheat protein: Study progress and prospective application[J]. J Chin Cereal Oils Assoc, 23(3): 209-213.[阮景军, 陈惠, 2008. 荞麦蛋白的研究进展与展望[J]. 中国粮油学报, 23(3): 209-213.]
SUETSUGU K, 2015. Autonomous self-pollination in the nectarless orchid Pogonia minor [J]. Plant Species Biol, 30(1): 37-41.
TANG W, ZHOU XL, WU Y, et al., 2010. Analysis of mineral elements in buckwheat seeds of 24 species[J]. J Chin Cereal Oils Assoc, 25(5): 39-41.[唐文, 周小理, 吴颖, 等, 2010. 24种荞麦种矿物元素含量的比较分析[J]. 中国粮油学报, 25(5): 39-41.]
TANG Y, ZHOU ML, BAI DQ, et al., 2010. Fagopyrum pugense(Polygonaceae), a new species from Sichuan, China[J]. Novon, 20(2): 239-242.
TSAI H, DENG H, TSAI S, et al., 2012. Bioactivity comparison of extracts from various parts of common and tartary buckwheats: Evaluation of the antioxidant-and angiotensin-converting enzyme inhibitory activities[J]. Chem Cent J, 6(1): 78-82.
USHIMARU A, DOHZONO I, TAKAMI Y, et al., 2009. Flower orientation enhances pollen transfer in bilaterally symmetrical flowers [J]. Oecologia, 160: 667-674.
WANG AH, CAI GZ, CHEN B, et al., 2011. Investigation and research of buckwheat resources in Sichuan Province and Southeast Tibet[J]. SW Chin J Agric Sci, 24(6): 2057-2061.[王安虎, 蔡光澤, 陈波, 等, 2011. 四川及藏东南荞麦资源的调查研究[J]. 西南农业学报, 24(6): 2057-2061.]
WANG XP, YU WB, SUN SG, et al., 2016. Pollen size strongly correlates with stigma depth among Pedicularisspecies[J]. J Integr Plant Biol, 58(10): 818-821.
WELLER SG, SAKAI AK, GRAY T, 2016. Variation in hete-rostylous breeding systems in neighbouring populations of Oxalis alpina(Oxalidaceae)[J]. Plant Biol, 18(1): 104-110.
WEN HH, CHEN ML, ZHANG ZS, 2015. Reproductive biology in the distylous species of Polygonum orientale [J]. Acta Pratac Sin, 24(11): 155-162.[文慧慧, 陈明林, 张自生, 2015. 二型花柱植物红蓼的繁殖生物学研究[J]. 草业学报, 24(11): 155-162.]
WORLEY AC, BAKER AM, THOMPSON JD, et al., 2000. Floral display in Narcissus: Variation in flower size and number at the species, population, and individual levels[J]. Int J Plant Sci, 161(1): 69-79.
WU LY, CHANG FF, LIU SJ, et al., 2018. Heterostyly promotes compatible pollination in buckwheats: Comparisons of intraflower, intraplant, and interplant pollen flow in distylous and homostylous Fagopyrum [J]. Am J Bot, 105(1): 108-116.
WU ZH, LUO XH, CHEN X, et al., 2010. Investigation of resources of Fagopyrum dibotrysfrom Qinling-bashan mountains in Shaanxi[J]. N Hortic, 34(14): 192-194.[吴振海, 罗小华, 陈西, 等, 2010. 陕西秦巴山区金荞麦资源调查[J]. 北方园艺, 34(14): 192-194.]
YAN XH, ZHOU B, YIN ZF, et al., 2016. Reproductive biological characteristics potentially contributed to invasiveness in an alien invasive plant Bidens frondosa [J]. Plant Species Biol, 31(2): 107-116.
YASUI Y, OHSAKO T, OHNISHI O, 1998. Evolutionary processes of Fagopyruminferred from the molecular phylogenetic analyses [C]// Advances in Buckwheat Research. Proceedings of the 7th International Symposium on Buckwheat at Winnipeg, Canada. Winnipeg: Manitoba University Press: 50-60.
ZHANG DY, JIANG XH, 2001. Mating system evolution, resource allocation, and genetic diversity in plants[J]. Acta Phytoecol Sin, 25(2): 130-143.[张大勇, 姜新华, 2001. 植物交配系统的进化、资源分配对策与遗传多样性[J]. 植物生态学报, 25(2): 130-143.]
ZHANG WL, ZHOU B, XIAO YA, et al., 2013. Reproductive ecology of distylous Fagopyrum dibotrys [J]. Acta Bot Boreal-Occident Sin, 33(3): 483-493.[张万灵, 周兵, 肖宜安, 等, 2013. 二型花柱植物金荞麦的繁殖生态学研究[J]. 西北植物学报, 33(3): 483-493.]
ZHOU B, YAN XH, XIAO YA, et al., 2013. Traits of reproductive biology associated with invasiveness in alien invasive plant Phytolacca americana [J]. Ecol Environ Sci, 22(4): 567-574.[周兵, 閆小红, 肖宜安, 等, 2013. 外来入侵植物美洲商陆的繁殖生物学特性及其与入侵性的关系 [J]. 生态环境学报, 22(4): 567-574.]
ZHOU ML, BAI DQ, TANG Y, et al., 2012. Genetic diversity of four new species related to southwestern Sichuan buckwheats as revealed by karyotype, ISSR and allozyme characterization[J]. Plant Syst Evol, 298(4): 751-759.
