东海中部浮游生态系统季节变化的数值模拟
2019-09-10陈建忠葛建忠BELLERBYRichard
陈建忠 葛建忠 BELLERBY Richard






摘要:利用一维物理一生物耦合模型(GOTM-FABM-ERSEM)对东中国海中部站位浮游生态系统要素的季节变化进行模拟,较好地刻画并分析了其物理、生化要素之间的相互作用.模拟结果表明浮游生态系统的季节性变化的物理控制因子主要为光照、温度及其引起的垂向层化;生化控制因子主要为营养盐水平,其夏季集中分布在跃层以下深度,并在9月达到最大值.模型较好地呈现了春秋季浮游植物的双峰结构,浮游植物在夏季次表层(约20 m)出现最大值,并在潮汐混合影响下呈周期性斑块状生长,峰值为5.3 mg C.m-3.浮游动物和细菌的分布与浮游植物类似,均在春季达到最大值,并滞后3d左右,细菌在夏季表层受浮游植物和温度影响.
关键词:东海;海洋数值模拟;浮游生态系统;季节变化
中图分类号:Q735
文献标志码:A
DOI: 10.3969/j.issn.1000-5641.2019.06.015
0 引言
海洋生态系统是生物圈的重要组成部分,海洋生态系统是指在一定的时间和海域中生物和非生物成分通过物质循环和能量流动相互作用而形成的统一体,它的功能主要体现在系统内物质循环、能量流动及其稳态调节机制[1].
东中国海(东海)是我国一个重要的大陆架边缘海,其主要受到黑潮、台湾暖流、浙闽沿岸流、苏北沿岸流、黄海环流等流系的影响,同时还受到长江径流影响,且不同季节长江径流的影响范围有所不同.河流是东海营养盐的重要来源,长江每年向东海输入的溶解无机氮(DIN)、磷酸盐(Poi一)、硅酸盐(Si03-)的总量分别达到7.84x10、1.5lx10、2.22x10 t[2].主要营养盐(硝酸盐、硅酸盐和磷酸盐)的总体分布趋势为由近岸到远海逐渐减少,垂向上一般随深度增加,并在夏季出现稳定的层化,此时底层达到一年中的最高值,冬季混合较为充分.外海的黑潮、局部地区上升流、生物活动和底栖过程也将对营养盐供给产生影响[3-4].
东海浮游植物的变化过程较复杂,是光照、温度、营养盐和被捕食压力等共同作用的结果,浮游植物生物量大致呈现出一年双峰的变化规律,即在春秋两季出现两次规模较大的藻华[5].浮游植物的垂向分布具有明显的季节性,冬季總体生物量小,春秋季藻华时期,维持较高的生物量水平,上下层差异明显,夏季浮游植物易在次表层出现最大值,生物量随深度先增加后减少[6].
东海浮游生态系统要素的季节性变化是物理与生物地球化学过程相互耦合作用的体现,采用数值模型是进行物理一生化耦合的重要方法.在物理过程的模拟方面,前人采用诸多数值模型对长江冲淡水在东海的季节性扩展[7]、夏季风暴潮[剐、黑潮跨陆架入侵东海[9]、泥沙输运[10]等物理过程进行模拟研究.基于这些三维海洋动力学模型,较好地刻画了东海区域的物理过程的季节变化,包括温度、盐度、环流及悬浮泥沙等.
而对于生物模型而言,准确的物理场是其进行生态过程模拟的前提和基础,孙科等采用一个三维生物一物理耦合模型ROMS-NEMURO对东海赤潮的典型藻种进行动态的数值模拟,并成功再现了优势藻种之间的竞争关系[11];闫庆等将三维ECOM-si与一个N2P2ZD模型相耦合,并对长江口外锋区浮游植物生物量及其影响因子进行数值模拟和分析[12];为研究长江冲淡水对东海生态环境的影响,贾守伟使用三维POM-NORWECOM耦合模型,对研究区域生态系统要素的季节变化和年际变化进行研究[13].
以上所述前人成果均为在三维模型的基础上进行,需要较为复杂的三维模型系统的支持,其建立合理模型涉及的过程较为复杂,难度较大.垂向一维数值模型也是进行生态动力学模拟和研究的有效工具[14-16],并在世界及中国海域已有诸多成功的应用[17-18],但在东海却鲜有报道,这与东海复杂的物理和生物地球化学环境有关.同时,垂向一维模型的合理模拟也是将其扩展到三维模型的前提.本文基于垂向湍流模型GOTM和生物地球化学模型ERSEM,采用FABM框架进行物理和生化过程的耦合,对东海的中部站位的温盐、营养盐、浮游植物、浮游动物和细菌等进行垂向的动态模拟,利用实测数据和卫星数据进行数据对比,重点分析控制东海季节性变化的物理和生物地球化学因子,并分析了生态系统各要素之间的相互作用关系,及季节和垂向的变化.
1 1D GOTM-FABM-ERSEM
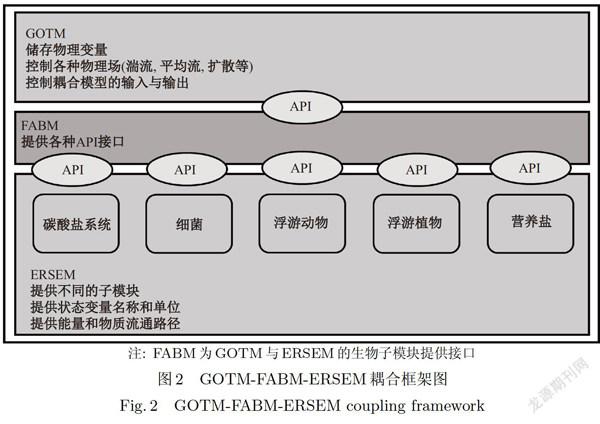
本文中所采用的垂向一维模型由物理和生物地球化学模型耦合而成,物理模型为通用海洋湍流模型General Ocean Turbulence Model (GOTM)[19],物理模型主要计算水动力过程,并为生物模型提供温度、盐度、水位、太阳辐射、外部压力梯度和湍流等必要的物理驱动,生物模型为European Regional Seas Ecosystem Model (ERSEM)[20],由于GOTM和ERSEM模型都是较为独立的模型,且各自输入、输出和计算都较为复杂,特别是ERSEM涉及的状态变量和生物过程众多,因此引入一个耦合框架模型Framework forAquatic Biogeochemical Models (FABM)[21]来对以上两个模型进行耦合,下面对这三个模型进行简单介绍.
1.1 物理模型GOTM
GOTM是比较通用的一维湍流模型,主要优势为采用结构模块化处理,即可独立运行,也可以与主流三维模型如FVCOM,ROMS等模型进行耦合.GTOM集合了大部分主要的湍流解决方案,如统计模型k-kl、k一ε和经验模型KKP等[22].
1.2 生物模型ERSEM
本文所使用的生物模型为ERSEM,在已出版的文献和研究中,ERSEM是最为成熟的低营养级海洋食物网模型之一[20],其模型结构图如图1所示,最新的版本包含了海洋生态系统水层(pelagic)和底栖层(benthic)所有的重要组成成分(如微生物食物网,碳酸盐系统,钙化过程等).
ERSEM垂直分层依次为大气、水层和底栖层.海洋和大气的界面中存在着C02,02和光等气体和能量交换,同时在此界面上大气也给海洋施加风、潮汐和太阳輻射等外界强迫和热量辐射,这些过程通常由与ERSEM耦合物理模型完成.在水层部分,模型主要包含了硅藻(20~200μm)、小型浮游植物(20~200 μm)、微型浮游植物(2~20 μm)和微微型浮游植物(<2 μm)四种浮游植物,纳米鞭毛虫(<20 μm)、微型浮游动物(20~200μm)和中型浮游动物(>200 μm)三种浮游动物,一种细菌、溶解有机物物质(DOM)和颗粒有机物质(POM)等.底栖部分主要包含两种底栖细菌(需氧细菌和厌氧细菌),三种底栖动物(悬浮摄食动物、小型底栖动物和沉积摄食动物),DOM和POM等.考虑的营养盐有硅酸盐、磷酸盐、铵盐和硝酸盐,在水层和底栖层之间通过湍流、底部再悬浮、捕食与被捕食、吸收、沉降和矿化作用等过程完成生物地球化学的物质循环和能量流动.
从图1可知,在ERSEM浮游生态系统框架下,浮游植物作为系统的生产者通过光合作用将光能转化为化学能,同时在生长过程中吸收营养盐以维持新陈代谢,浮游动物通过捕食,将浮游植物的初级生产力进行同化,细菌则通过分解和矿化作用将水中的DOM和POM转化为营养盐,同时细菌本身也需要消耗一定的营养盐,DOM和POM主要来源于生命有机体死亡和排泄等过程.
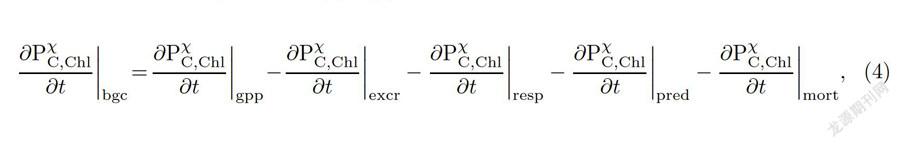
浮游植物动态生长过程是以每种浮游植物类型的吸收和损失来定义的,浮游植物碳和叶绿素控制方程如式(4)所示,浮游植物营养元素控制方程如式(5)所示,与浮游植物类似,浮游动物和细菌的控制方程如式(6)-(9)所示[20]:在方程(4)-(9)中,P代表浮游植物,Z代表浮游动物,B代表细菌,X代表不同浮游植/动物类型,C和Chl代表生物体的碳含量和浮游植物叶绿素含量,左边bgc (biogeochemical)为各种元素(或者叶绿素)生物地球化学过程总的变化率,gpp(gross primary production)为总初级生产力变化项,excr(excretion)为排泄变化项,resp(respiration)为呼吸变化项,pred(predation)为被捕食变化项,mort (mortality)为死亡变化项,upt(uptake)为吸收变化项,rel(release)为释放(如溶解有机质的释放)变化项,N,P,F,[,S]分别代表生物体的氮、磷、铁和硅元素,硅只在硅藻考虑.以方程(4)为例,左边第一项表示浮游植物碳或者叶绿素总的变化率,它等于右边由光合作用产生的总生产力变化率减去排泄、呼吸、被捕食和死亡的变化率.
1.3 耦合连接模型FABM
在生物模型和水动力物理模型的耦合过程中通常面临很多的挑战,除模型自身编写语言不同的原因,这些模型一般涉及很多专业性很强的理论知识,这就需要模型的使用者在具有较强的编程基础上进行耦合,难度相对较大.传统的生物一物理耦合过程都是通过一系列专用的代码或者接口把两个模型封装在一起,这些接口一般移植性差,且不容易维护.基于上述问题,本文在耦合过程中引进Jorn Bruggeman等人开发的耦合框架模型FABM[21],其主要原理是在保持原始模型没有改动的前提下,提供一个通用的、易于使用的高性能耦合层,该耦合层将物理模型和生物模型连接起来,并通过运用程序界面接口(API)在模型间进行信息传递和数据交换.GOTM-FABM-ERSEM的耦合框架如图2所示.
2 模型设置与数据来源
2.1 研究位置
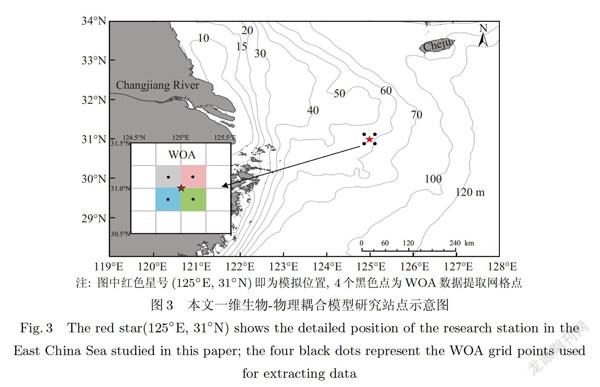
本文研究位置如图3所示,其具体位置为东经1250、北纬310,水深大约为60 m.该研究站点位于离岸较远的陆架浅水海区,处于长江冲淡水和台湾暖流等流系影响的边缘区域,不存在明显的水文要素锋面[23-24],同时所收集的资料可以提供该位置的主要生态系统要素的垂向剖面季节变化数据.
2.2 模型配置
2.2.1 GOTM设置
本文所用的耦合模型在Linux集群上运行,采用Intel C/Fortran混合编译.GOTM的运行时间长度为10年,从2005年1月1日开始,到2015年1月1日结束.模型水深采用研究海区典型水深,设置为60 m,垂向设置均匀分布的100层,时间步长300 s,每天输出一个数据,密度由温盐的状态方程计算而来.物理模型关键参数如表l所示,
2.2.2 ERSEM设置
由于ERSEM在中国海域运用得较少,且模型状态变量众多,所以模型参数选取以ERSEM常用参数为主,在此基础上结合东海具体生物种群结构和优势种情况,浮游生物主要参数取值如表2和表3所示.模型营养盐的初始值使用《中国海洋图集》海洋化学分册C16-3(125°E,31°N)站位的冬季平均值,硝酸盐、铵盐、磷酸盐和硅酸盐的初始值分别为7.56 μmol/L、0.14 μmol/L、0.55 μmol/L、8.44 μmol/L[25].
2.3 数据来源
2.3.1 温盐及大气驱动数据
根据GOTM的设计原理,为使温度方程和盐度方程求解,模型需要用户提供研究站位的温盐垂直剖面数据,这些温盐剖面可以来自实地测量,也可以由三维海洋动力学数值模型提供.本文所用的温盐剖面、水位和外部压力梯度数据均取自葛建忠等人的东海三维FVCOM模拟结果[26].提取数据的时间间隔均为lh一次,温盐剖面垂向采用s分层.本文所使用的海表强迫数据包含海表风速和风向(10 m)、大气云量、大气辐射、大气温度(2 m)、大气相对湿度和海表压力,海表强迫数据均来自European Centre for Medium-Range Weather Forecasts(ECWMF).空间分辨率为0.125°,时间间隔为3h一次[27].此外,由于在FVCOM中使用的湍流模块为GOTM,所以本文直接用耦合之后的GOTM-FABM-ERSEM模型计算湍流相关的物理参数,这将与耦合的生物模型连接得更紧密,并有利于文章后面的结果分析.
2.3.2 营养盐、温盐观测数据
营养盐(硝酸盐,磷酸盐和硅酸盐)垂向剖面数据来自由国家海洋局出版的《中国近海海洋图集》海洋化学分册的C16-3(125°E,31°N)站位數据,该站位包含春(4月)、夏(7月)、秋(10月)、冬(12月)四个季节的垂直剖面数据[25].营养盐剖面数据观测时间在2006 2007年,本文采用与观测数据同一年同一月份15日的模拟数据与观测数据对比,并用2006 2007年的模拟结果分析其季节和深度变化.温盐垂直剖面数据来自WOA13(World Ocean Atlas 2013)的统计月平均数据(2005-2012年),空间分辨率为0.25° [28-29],因研究站位不在WOA的数据网格点上,所以本文使用的数据为与研究站位相邻四个网格点的平均值(如图3所示).
2.3.3 叶绿素数据
叶绿素垂直剖面数据收集自研究站点(1250E,3IoN)附近0.50范围内的World OceanDataset(WOD)数据[30].卫星表层叶绿素数据,取自由英国普利茅斯海洋实验室的再分析表层叶绿素遥感数据(https://www.oceancolour.org/),时间分辨率为l天1次,空间分辨率为9 krrl[31].
3 结果和分析
3.1 物理过程季节变化
3.1.1 光照和混合层
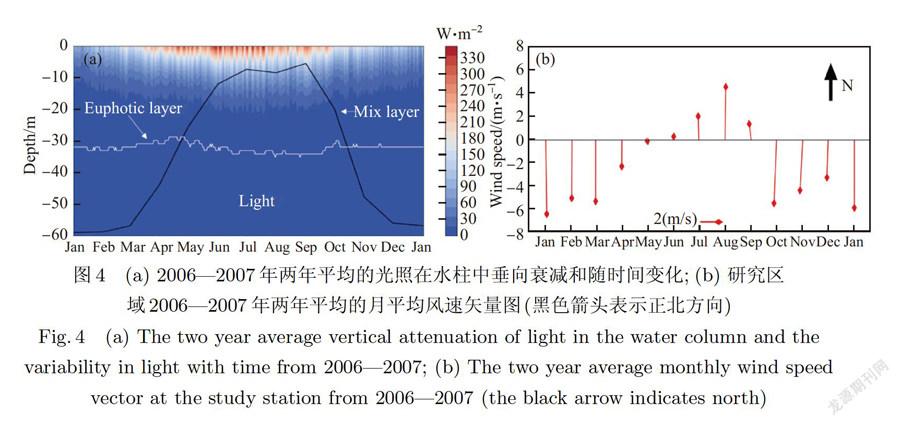
季节性的太阳辐射及水一气界面的热量交换在水体表层形成加热和冷却,从而造成的水体的升温和降温,以及垂向混合层的季节性变化,这也是水体季节性变化的主要驱动物理过程.光照(light)和混合层(mix layer)的深度如图4(a)所示,海表所吸收的太阳辐射在2月开始增强并在6-9月达到高值,其日平均峰值达到330W.m左右,9月底开始减少,并在1月左右达到低值.光在垂向上由于水体、颗粒物、叶绿素等物质的吸收和散射作用随深度迅速衰减,上层水体吸收了大量的太阳辐射.一般而言,真光层(Euphotic layer)深度有两种定义方法,一是将真光层深度定义为海面光合有效辐射的1%深度;二是把真光层深度定义为恒定值14 μmol·m.s,其是对光子流强度的测量,而在模型中光的单位是W·m,其是对光能量流的直接测量,它们之间并不是线性的换算关系[32- 33].在东海海域,本文使用“1%”方案,该区域受长江口泥沙输运的影响较弱,模拟的真光层深度稳定在3lm左右,季节变化不大.
海水混合程度与表面风应力和海水温度有关,夏季海表吸收热量最多,导致海水温度升高,造成垂向分层加强,且由于夏季前后平均风速较弱,导致湍流很难向深层发展,其深度一般小于10 m,冬季由于表层降温,向下混合加强,海水基本没有层化,同时受到冬季季风较强(图4(b)),湍流作用明显,其深度几乎可以到达底层.同时,为了与营养盐实测数据同步分析,在图4中描述的物理量皆是2006 2007年两年的平均数据.
3.1.2 温度和盐度
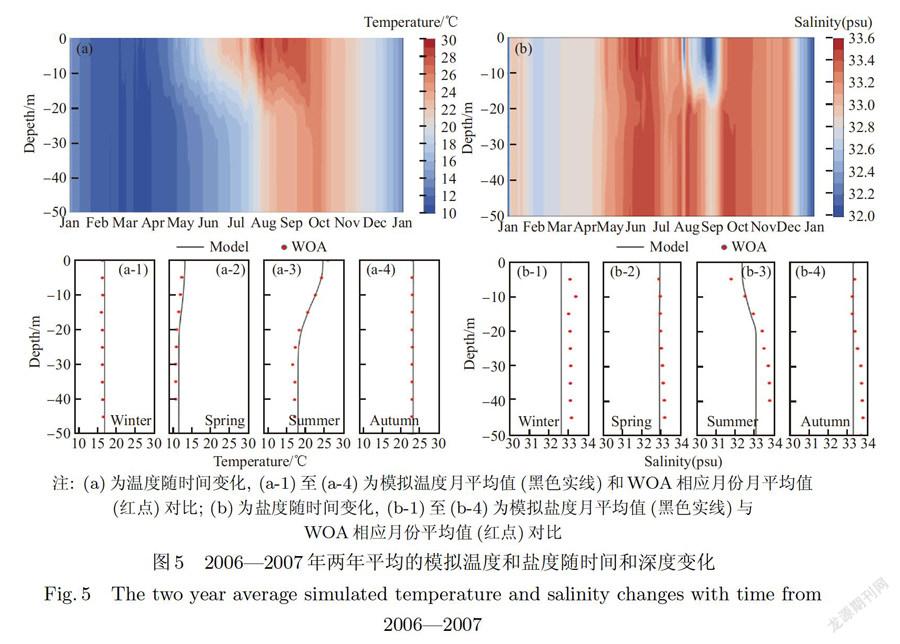
模拟得出的温盐垂直结构和季节变化以及与WOA数据对比结果如图5所示,由对比结果可以看出,模型较好地模拟了研究站位的温盐场.1月,由于太阳辐射角度较小且光照时间短,气温较低,水一气界面的能量交换主要以向空气的热量散失为主,水体降温(如图5(a)),海水上下混合充分,温盐垂向分布均匀.大约3月之后,季风开始转弱,太阳辐射增强,温度开始回升,水层分层逐渐形成,4月的盐度在32.8~33.0之间,水柱混合均匀.6月由于处在冬夏季风转换期.风速减弱到全年最小(月平均1 m/s),混合层深度浅,且温度开始进一步升高,水柱分层显著,盐度垂向出现波动,并且达到最高值33.6,这可能与高盐的台湾暖流入侵有关.在8-9月,由于海水温度达到一年中最高290,且夏季季风开始向冬季季风过渡,导致海水层化最强,表层混合主要由风驱动,混合层深度仅为5m左右,同时盐度也达到一年中的最低值32.0,且存在强烈分层,这和安琰等人的研究结果一致[34].从10月开始逐渐转入冬季季风期,混合层深度加深,温盐均有下降趋势,重新趋于均匀分布.总体而言,在温盐与WOA数据的对比中,温盐对比结果良好,模拟结果在垂向都分布得比较均匀,这可能是由水深较浅(60 m),离岸较远,温盐锋面较弱导致的.
3.2 营养盐季节变化
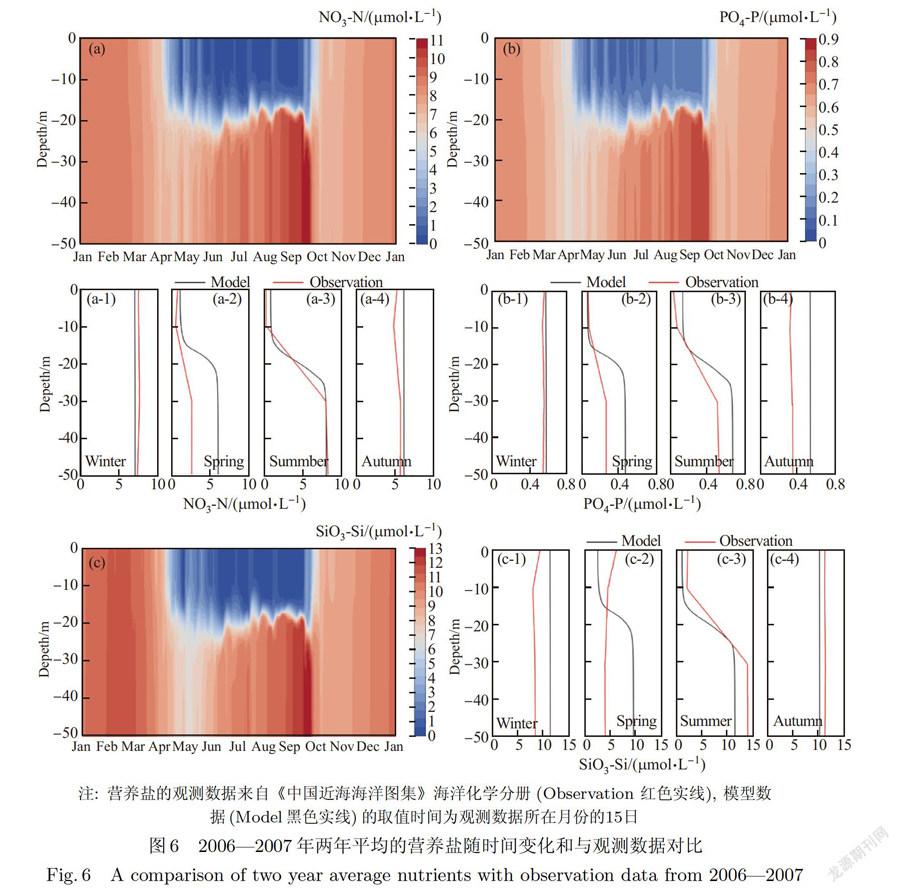
在物理过程模拟良好的基础上对生态过程进行模拟分析,本文营养盐考虑硝酸盐(N03),磷酸盐cP03-)和硅酸盐(Si02-),其模拟结果,以及与《中国近海海洋图集》海洋化学分册数据对比如图6所示.营养盐的垂向分布与很多物理和生物化学过程息息相关,从该站位模拟营养盐数据和海洋图集数据对比可以看出,耦合模型较好地模拟了东海中部站位的营养盐垂直和季节变化.值得注意的是,《中国近海海洋图集》数据是取自当月的某一天的数据,具体日期不详,而模型的对比数据的时间取自当月的15日,所以在数据对比的过程中模型在数值上可能存在误差,但不影响反映营养盐垂向变化总体趋势.
冬季海水混合充分,温度较低,光照时间短,生物新陈代谢缓慢,营养盐垂向分布均匀且浓度较大,3月末,温度回升,海表吸收太阳辐射增加,温度升高,浮游植物开始大量生长,导致上层营养盐减少,海洋生物f浮游动物,浮游植物和细菌)在经过排泄和死亡等生化过程之后,大部分分解成有机颗粒物质(POM)和溶解有机物(DOM),POM经过一系列沉降、分解、再悬浮和矿化等过程之后,重新以海洋生物可利用的形式返回到水体中,而这些过程将导致水体的中下层的营养盐浓度升高.6月开始,海水上层温度迅速上升,海表所受风应力较小,混合层深度维持在10 m以内,层化阻碍底部营养盐对上层补给,营养盐聚集在真光层深度以下,在9月左右,由于季风转向且温度最高,混合层深度达到最小5m,营养盐在底部出现峰值(硝酸盐、磷酸盐和硅酸盐的最大值分别为10.7 μmol/L、0.85μmol/L和12.3 μmol/L),此时的海水的层化最稳定.10月以后,北方季风加强,混合层开始向底部发展,营养盐跃层弱化,温度适宜,浮游植物等海洋生物迎来新一轮生长,但与春季相比,营养盐垂向分布较均匀.
3.3 浮游植物季节变化
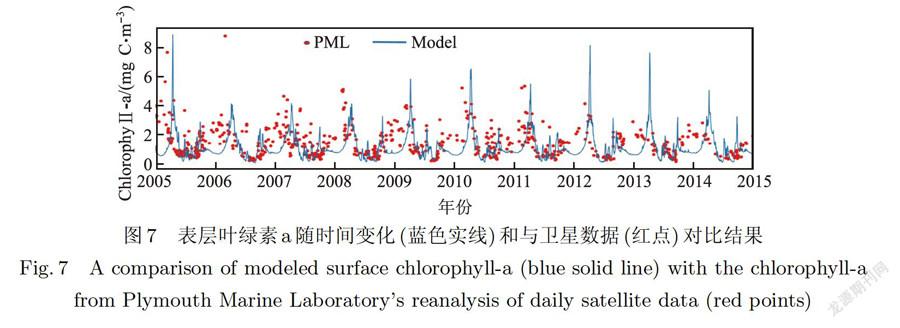
浮游植物生长状态也是物理一生态过程共同作用结果,如营养盐的供给、光照、温度等,由海洋生态系统的食物网关系可知,浮游植物作为极为重要的生产者,影响着生态系统的物质和能量的流动.在ERSEM中细胞碳与细胞叶绿素的比值(C:Chl-a)是动态的,这意味着叶绿素可以动态地反映植物的生长状态.为研究表面强迫和光照对浮游植物季节变化的影响,本文为模型提供10年相同的温盐输入,其模拟的10年(20052015年)日均表层叶绿素数据与遥感叶绿素数据进行对比,其结果如图7所示.除冬季时段外,模型可以较好地再现浮游植物的生长趋势,春秋(10年平均均方根误差(RMSE)为1.56 mg C.m和0.76 mg C.m)分别出现一次叶绿素的峰值,春季峰值一般比秋季大,并在夏季(10年平均RMSE为0.81 mg C.m)达到全年的最低水平,冬季(10年平均RMSE为1.83 mg C.m)模拟结果不准确可能是由于该时段较为强烈的台湾暖流及黑潮的跨陆架输运中的外源输入叶绿素所造成的,
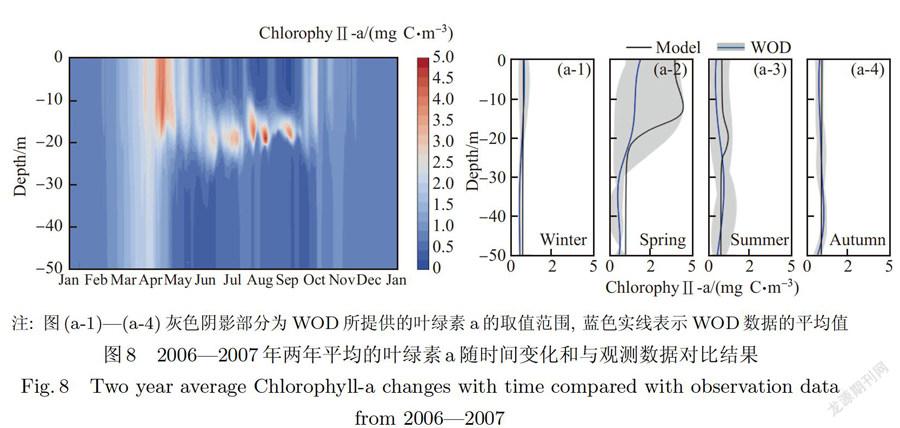
浮游植物随时间变化、垂向分布及与观测数据对比结果如图8所示.在叶绿素模拟数据与观测数据对比过程中,观测数据采用的是世界海洋图集(WOD)中研究站位附近的數据(经度125°±0.5°,纬度31°±0.5°).因WOD数据库中,研究区域的数据量较少,所以本文选取从1990年开始的垂向叶绿素数据,为模拟叶绿素结果提供一个参考范围(图8(a-l)到(a-4)阴影区域1.
总体而言,真光层以上的浮游植物的分布状况和营养盐的上层分布趋势类似,但不同季节主要控制因素不一,真光层以下水层由于受光限制,浮游植物生物量全年保持较低水平.冬季海表吸收太阳辐射少,海水温度低,浮游植物生长受到光和温度的限制,生物量小,混合层深度大,垂向分布均匀.3月下旬,温度上升,光照充足,浮游植物开始迅速生长,峰值达到4.O mg C.m左右.6月开始,温度迅速上升,季风向偏南季风转换,风力减弱,混合层变浅,分层稳定,营养盐向上输运困难,上层营养盐匮乏,且此时太阳辐射强度在四季中最强,除营养盐外,温度和光照对上层浮游植物的生长也有抑制作用,叶绿素在水柱上层的含量达到一年中的低值(小于0.5 mg C.m),并一直持续到9月.在海水次表层(约20 m),温盐跃层和营养盐跃层现象有所减弱,温度维持在18--.22℃,营养盐充足,并处在真光层之内,浮游植物生长条件均满足并开始生长,如图8所示,浮游植物在此阶段经历3~4次不同程度的峰值,并在期间达到最大值5.3 mg C.m,这主要是由于在天文大潮期间,海水垂向湍流动能增大,将底部营养盐向上输运到跃层下部,促进浮游植物的生长.10月.海面开始盛行北风,混合层加深,温度相比夏季开始降低,营养盐向上输运增强,此时温度、光照和营养盐都比较适合植物生长,表层浮游植物开始第二次水华,此时次表层叶绿素浓度最大现象消失,但表层水华规模与春季相比较小,其峰值约为2.6 mg C.m. 11月之后,虽然水体混合充分,营养盐充足,但此时温度降低,且可利用的光照通量减少,浮游植物生长开始变缓,并一直持续到来年春季.
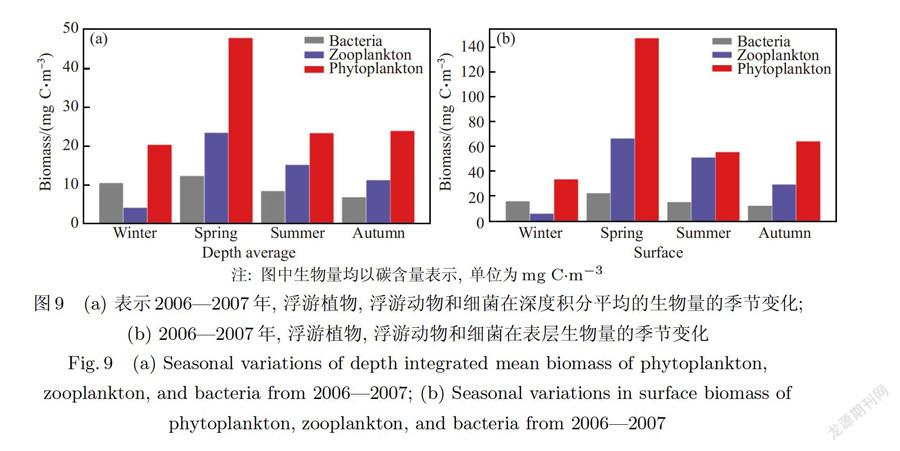
浮游植物由于受到诸多因子的共同影响,其生长具有明显的季节变化趋势(见图8和图9),且其在表层和深度平均(表征整个水柱变化趋势)生物量的季节变化存在不同.表层表现为春季>秋季>夏季>冬季,而在深度平均表现为春季最大,夏秋大致相等,冬季最小.造成浮游植物表层和深度平均的生物量在夏秋季节变化差异的原因是浮游植物在夏季表层由于受到营养盐和温度限制生物量较低,而在秋季表层存在秋季赤潮;但在夏季次表层存在生物量峰值且其大于秋季赤潮,经过深度平均之后将出现较高值,同时秋季浮游植物在中下层生物量也处于较低水平,所以夏秋深度平均大致相当.
3.4 浮游动物和细菌的季节变化
浮游动物和浮游细菌的模拟结果如图10所示,浮游动物和细菌的变化趋势与浮游植物具有很强的相关性,在浮游植物达到峰值之后,浮游动物和细菌相继达到峰值.浮游动物能量来源主要为浮游植物,所以其季节变化趋势与浮游植物很相似,在表层和水柱平均生物量均表现为春季>夏季>秋季>冬季(见图9),这与李晓慧等人的研究结果一致[35].浮游动物和细菌的最大值均出现在春季,分别为150mg C.m和39 mg C.m.夏季两者在垂直方向上都出现明显的跃层,在真光层以下浮游动物与浮游植物分布类似,含量较低.在9月,上层浮游动物和细菌分别出现了低值25 mg C.m和9 mg C.m,主要与营养盐匮乏和浮游植物生物量在此期间较低有关,这与杨位迪等人的调查结果相符[36].秋季因浮游植物增加浮游动物生长迅速,冬季浮游动物随着浮游植物的减少生物量保持一年最低水平(约为8 mg C.m),且因水体混合强烈,其垂向上分布均匀.
细菌作为生态系统的分解者,影响其生物量的因素较多,包括浮游动物、浮游植物、溶解氧、营养盐和温度等,具有明显季节变化和垂向变化,水柱深度平均生物量为春季>冬季>夏季>秋季(图9(a)),而在表层生物量为:春季最高,冬季和夏季大致相等,秋季最低(图9(b).造成这种夏、冬生物量季节变化差距的主要原因是夏季海水垂向分层稳定,表底生物量差距较大,且底部较低,所以导致其深度平均生物量比冬季低.细菌生物量在冬季比秋季高的可能原因是冬季水体中有充足的营养盐可以利用,且与其存在竞争关系的浮游植物受到光和温度的限制生长缓慢.春季,浮游植物和浮游动物迅速繁殖之后产生的大量POM和DOM给细菌提供了充足的食物潜力,由于浮游植物和浮游动物的生长繁殖需要一定的时间,所以浮游细菌生物量的峰值滞后3d左右.除春季外,整体而言营养盐对细菌生物量的影响不是很大,李云等人的研究认为夏季浮游植物和温度是影响研究区域细菌生物量的主要因素[37],而模型的模拟结果反映了这个趋势.在夏季的中下层,细菌在真光层以下可以维持16 mg C.m左右的生物量,主要原因可能是细菌可以吸收营养盐和通过分解溶解有机质(DOM)与颗粒有机物质(POM)来维持新陈代谢,这些DOM和POM会在湍流和重力(针对POM)作用下向底部输送.细菌和浮游动物的最低值出现在9月的中下层,其生物量分别小于9 mg C.m和15 mg C.m,一个重要原因可能是水体溶解氧(文中未列出数据)的减少,秋季由于细菌生长所需的条件都比较满足,细菌垂向分布均匀,平均保持8 mg C.m左右的生物量(图9(a)),虽然冬季海水温度较低,但细菌依然保持一定的生物量(10 mg C.m以上).
4 讨论
本文利用一维垂向GOTM-FABM-ERSEM模型,较好地模拟了东海中部站位的浮游生态系统各种要素的垂向和季节分布.与传统的三维耦合模型相比,本文的一维耦合模型对特定水柱的生物地球化学过程的刻画保持较高的精度,可以更深入地理解海洋生态系统各要素的相互关系.同时,一维模型还有计算简单、计算效率高、考虑物理因素(开边界条件及河流输入等)少等优点.但不可否认一维模型与三维模型相比固然存在部分局限性,如只考虑海水的垂向交换,难以考虑水平输运,但实际上研究海区可能受到长江冲淡水的营养盐输入、台湾暖流和黑潮等海洋水平环流的影响.同时本文在进行模型结果与观测结果对比时存在数据较少和数据时间较久远等问题,这主要是因为研究区域相对于传统东海考察作业区距离较远,可收集数据较少.
在模型结果与卫星数据对比的过程中,冬季模拟结果与卫星数据的观测误差较大,可能原因如下:研究区域在冬季可能会受到台湾暖流和黑潮分支的影响,这将为该区域带来外源的营养盐、浮游植物及叶绿素;在一维模型中难以刻画这一外源输入过程;在模拟过程中,模型10年输入所使用的是相同温盐剖面,所以冬季低值也有可能是温度输入过低造成的结果.此外,冬季云量较多,卫星数据本身也存在较大误差.
在夏季次表层叶绿素最大值深度附近,浮游生物的分布存在明显的半月周期性震荡(见图8(a)和图9),这主要是受到潮汐混合的周期性影响.在天文大潮时,潮流引起的湍流能量最强,将水层下层的营养盐输运到跃层下方,在有利的光照影响下,此时浮游植物开始迅速增长,浮游动物和细菌同步生长,在浮游植物的上方3~5 m出现细菌的较大值.
5 结论
本文利用一维物理一生物耦合模型GOTM-FABM-ERSEM对东海浮游生态系统的温盐、营养盐(硝酸盐、磷酸盐和硅酸盐)、浮游植物、浮游动物和细菌等要素进行模拟,并对大部分要素进行了观测数据的对比验证.并结合现有的理论成果,对控制季节性变化的物理和生物地球化学因子及其相互影响进行模拟分析.
从模拟结果可知,营养盐在秋冬季节垂直混合比较均匀,在春季逐渐出现跃层,并在69月出现稳定的跃层,在9月次表层叶绿素浓度以下,各种营养盐均达到峰值,硝酸盐、磷酸盐和硅酸盐的最大值分别为10.7μmol/L、0.85 μmol/L和12.3 μmol/L.模拟结果与观测数据显示了良好的一致性.浮游植物在一年中呈现出典型的双峰结构,分别在春季和秋季各有一次水华现象,夏季在20 m左右呈现次表层叶绿素浓度最大的现象,并在8月达到一年的最大值5.3 mg C.m.浮游植物的垂向和季节分布状态主要取决于温度、光照、摄食压力和营养盐可得性.浮游动物和细菌的分布趋势与浮游植物类似,并在春季存在3d左右的滞后期,这与它们在生态系统中所处的营养级有关.浮游动物和细菌在春季达到最大值150 mg C.m和39 mg C.m,两者主要分布在叶绿素浓度最大值附近,浮游动物一年也呈现出双峰态势,相比春季,秋季峰值较小,只有60 mg C.m左右.细菌的分布与多种因素相关,其分布除了9月在中下层之外,在水柱中均可保持一定的生物量.夏季浮游生物的周期性斑块状分布与潮汐和光照变化有关.
总体而言,一维耦合模型可以较为合理地模拟东海中部陆架区域生物地球化学过程及其季节性变化,为下一步的三维物理一生物地球化学耦合模型的建立打下了良好的基础.
[参考文献]
[1] 沈国英,黄凌风.海洋生态学[Ml.2版.北京:科学出版社,2002.
[2] 姜加虎,王苏民.长江流域水资源、灾害及水环境状况初步分析[J].第四纪研究,2004, 24(5): 512-517.
[3] 白涛,杨德周,尹宝树.夏季长江口外海区域上升流现象的数值研究[J].海洋科学,2009, 33(11): 65-72.
[4] 董书航.东海营养盐分布特征及跨陆架交换研究[D].山东青岛:中国海洋大学,2015.
[5] 孙百晔.长江口及邻近海域浮游植物生长的光照效应研究[D].山东青岛:中国海洋大学,2008.
[6] 文斐,孙晓霞,郑珊,等.2011年春、夏季黄、东海叶绿素a和初级生产力的时空变化特征[J].海洋与湖沼,2012, 43(3):438-444.
[7] 朱建荣,肖成猷,沈焕庭.夏季长江冲淡水扩展的数值模拟[J].海洋学报:中文版,1998, 20(5): 13-22.
[8] 葛建忠,胡克林,丁平兴风暴潮集成预报可视化系统设计和应用[J].华东师范大学学报(自然科学版),2007(4): 20-25.
[9] 杨德周,许灵静,尹宝树,等.黑潮跨陆架入侵东海年际变化的数值模拟[J].海洋与湖沼,2017(6): 1318-1327.
[10] 杨雪飞.基于GOCI和数值模拟的东海近岸悬浮泥沙浓度逐时变化研究[D].上海:中国科学院研究生院(上海技術物理研究所),2016.
[11] 孙科.东海典型赤潮藻种群动态的数值模拟[D].山东青岛:中国科学院研究生院(海洋研究所),2013.
[12] 闫庆.长江口外锋区浮游植物生物量及其影响因子的观测与数值模拟[D].上海:上海海洋大学,2016.
[13] 贾守伟.长江冲淡水对长江口附近海域生态环境影响的数值研究[D].山东青岛:中国海洋大学,2014.
[14]BLACKFORD J C, BURKILL P H. Planktonic community structure and carbon cycling in the Arabian Sea asa result of monsoonal forcing: The application of a generic model [J]. Journal of Marine Systems, 2002, 36(3):239-267
[15]VICHI M, PINARDI N, ZAVATARELLI M, et al. One-dimensional ecosystem model tests in the Po prodeltaarea [J]. Environmental Modelling& Software, 1998, 13(5): 471-4812.
[16]ALLEN J I,BLACKFORD J C, RADFORD P J.An l-D vertically resolved modelling study of the ecosystemdynamics of the middle and southern Adriatic Sea [J]. Journal of Marine Systems, 1998, 18(1/2/3): 265-286.
[17]PAN s,SHI J, GAO H, et al. Contributions of physical and biogeochemical processes to phytoplankton biomassenhancement in the surface and subsurface layers during the passage of Typhoon Damrey [J]. Journal of Geo-physical Research: Biogeosciences, 2017, 122(1): 212-229.
[18] 夏潔,高会旺.南黄海东部海域浮游生态系统要素季节变化的模拟研究[J].安全与环境学报,2006, 6(4): 59-65.
[19]BURCHARD H, BOLDING K, VILLARREAL M R. GOTM,a general ocean turbulence model: Theory, imple- mentation and test cases [R]. Space Applications Institute, 1999.
[20]BUTENSCHON M, CLARK J,ALDRIDGE J N, et al. ERSEM 15.06: Ageneric model for marine biogeochem-istry and the ecosystem dynamics of the lower trophic levels [J]. Geoscientific Model Development, 2016, 9(4):1293-1339.
[21] BRUGGEMAN J,BOLDING K.A general framework for aquatic biogeochemical models [J]. Environmental[elling& Software, 2014, 61(C): 249-265.
[22] MELLOR G L,YAMADA T. Development of a turbulent closure model for geophysical fluid problems [J].Reviews of Geophysics&Space Physics, 1982, 20(4): 851-875.
[23] 翁学传,王从敏,台湾暖流水的研究[J].海洋科学,1985, 9(1):7-10.
[24] 张文静.长江冲淡水扩展区域低盐水团的数值模拟及动力机制分析[D].山东青岛:中国海洋大学,2010.
[25] 暨卫东.中国近海海洋图集.海洋化学[M].北京:海洋出版社,2012.
[26]GEJZ, DING P X, CHEN c s,et al. An integrated East China Sea-Changjiang Estuary model system with aimatresolving multi-scale regional-shelf-estuarine dynamics [J]. Ocean Dynamics, 2013, 63(8): 881-900.
[27]DEE D P,UPPALA s M, SIMMONS A J,et al. The ERA-Interim reanalysis: Configuration and performance ofthe data assimilation system [J]. Quarterly Journal of the Royal Meteorological Society, 2011, 137(656): 553-597.
[28] LOCARNINI R A, MISHONOV A V, ANTONOV J I,et al. World Ocean Atlas 2005 Volume l:Temperature[R]. NOAA Atlas NESDIS, 2006.
[29] ANTONOV J,SEIDOV D, BOYER T, et al. World Ocean Atlas 2009, Volume 2:Salinity [Rl. Ocean ClimateLaboratory, National Oceanographic Data Center, 2010.
[30]BOYER T P,ANTONOV J I,BARANOVA 0, et al. World ocean database 2013 [Jl. Data Science Journal, 2013,90(49): 153-173.
[31] BROTAS V, GRANT M, CHUPRIN A, et al. In-situ databases and comparison of ESA Ocean Colour ClimateChange Initiative (OC-CCI) products with precursor data, towards an integrated approach for ocean colourvalidation and climate studies[C],/EGU General Assembly, 2014.
[32]ERM A, ARST H: TREI T, et al. Optical and biological properties of Lake Ulemiste,a water reservoir of the cityof Tallinn I: Water transparency and optically active substances in the water [J]. Lakes&Reservoirs Research& Management, 2010, 6(1): 63-74
[33] REINART A, ARST H: ERM A, et al. Optical and biological properties of Lake Ulemiste,a water reservoir ofthe city of Tallinn II: Light climate in Lake Ulemiste [J]. Lakes&Reservoirs Research&Management, 2010,6(1): 75-84.
[34] 安琰.東海PN断面及邻近海域温盐及化学要素月季变化特征研究[D].上海:上海师范大学,2009.
[35] 李晓慧,刘镇盛.长江口及邻近海域浮游动物生物量分布及季节变化[J].海洋学研究,2017(4): 94-101
[36] 杨位迪,郑连明,李伟巍,等.长江口邻近海域夏季大中型浮游动物物种多样性、年际变化及其影响因素[J].厦门大学学报(自然科学版),2018(4): 517-525.
[37] 李云.长江口及其邻近海域浮游异养细菌、寡营养细菌、光合细菌的分离鉴定、分布规律及与生态环境因子关系[D].上海:华东师范大学,2005.
(责任编辑:李万会)
收稿日期:2018-09-06
基金项目:国家重点研发计划(2016YFA0600903);国家自然科学基金(41776104,41476076)
第一作者:陈建忠,男,硕士研究生,研究方向为海洋数值模拟.
E-mail: chenjianzhong1003@outlook.com.
通信作者:葛建忠,男,副研究员,硕士生导师,研究方向为海洋环境动力学数值模拟.
E-mail: jzge@sklec.ecnu.edu.cn.
