环境因子对互花米草定居潮滩的影响分析
2019-09-10朱晓泾袁琳赵志远张利权李伟魏伟潘家琳陈雅慧
朱晓泾 袁琳 赵志远 张利权 李伟 魏伟 潘家琳 陈雅慧




摘要:外来物种互花米草入侵给滨海湿地生态系统带来一系列生态危害.为了探究潮滩环境因子对互花米草入侵的影响,在上海南汇东滩选择了砂质与粉砂质两种不同类型的潮滩,由陆向海移栽互花米草根茎苗,同步监测不同样点高程、水动力、沉积物特性等环境因子及互花米草存活情况.通过冗余分析(Redundancy Analysis,RDA),研究影响互花米草根茎苗定植的主导环境因子.结果表明:(1)临界剪切应力、中值粒径、高程和最大流速对互花米草根茎苗定植有主导作用(P<0.05).(2)当潮滩类型不同时,影响互花米草根茎苗定植的主要环境因子也有所不同,具体表现为在北部砂质潮滩上,高程和中值粒径是影响互花米草定植的主导因素(P<0.05),高程越高、中值粒径越小,互花米草根茎苗的存活率越高;在南部粉砂质潮滩上,临界剪切应力是互花米草根茎苗定植的主导因素(P<0.05),并与互花米草根茎苗定植存在正相关关系.上述结果对互花米草入侵的机理研究与防控管理具有理论价值与指导意义.
关键词:互花米草;根茎苗;入侵;潮滩; 临界剪切应力
中图分类号:Q948.1
文献标志码:A
DOI: 10.3969/j.issn.1000-5641. 2019.06.014
0 引言
互花米草(Spartina alterniflora)隶属禾本科米草属,原产于大西洋西海岸及墨西哥湾,根系发达、植株稠密,主要分布在平均海平面至平均高潮位之间的广阔滩面及河口湾[1].由于人类有意引种或无意带入,现已成为全球海岸盐沼生态系统中危害最严重的入侵植物之一[2—3].中国于1979年从美国引入互花米草,1982年起向全国沿海地区推广[4].目前互花米草已遍布我国南至广东雷州,北至辽宁葫芦岛的沿海地区[5].互花米草被引入后,凭借其强大的繁殖能力、适应能力和种间竞争力[1-2,6],迅速占领潮间带空生态位而导致光滩面积减少和生态环境改变,给滨海湿地生态系统带来了诸多负面影响[7-9].
互花米草主要通过种子、实生苗或断落的根茎苗进行跳跃式传播[2],定植在潮滩的繁殖体会受潮水频繁的淹没、波浪的拍打拖拽及泥沙侵蚀或掩埋的威胁,只有当繁殖体能够抵御环境胁迫与干扰时,才能成功入侵[10-11].这一过程一方面取决于繁殖体本身的生物因素(如繁殖体大小、繁殖体类型)[12-13],另一方面则取决于高程[14-15]、水动力条件[16-18]和沉积物特性[11,13]等环境因素.李蕙等[12]和Schwarz等[13]在野外的移栽实验均表明,繁殖体单元越大,植物存活率越高,高初始密度的植丛可以保持较高的存活率.在环境因素方面,动态的潮滩环境对互花米草入侵具有重要的影响作用[16,19-20].已有研究表明,高程[21]、波浪[22]、潮流[23]、沉积物特性(如中值粒径[13]和沉积物组成[11])和临界剪切应力[24]会影响盐沼植物的定居过程.但综合分析多个环境因子对互花米草入侵的影响及比较各环境因子重要性的研究,仍有待进一步深入.
为此,本研究在上海南汇东滩的自然潮滩,沿环境梯度移栽互花米草根茎苗,模拟互花米草植株入侵潮滩的情形.通过野外监测与室内实验,研究潮滩高程、水动力条件及沉积物特性等环境因子的变化及互花米草繁殖体的存活动态,采用冗余分析(Redundancy Analysis,RDA)的方法分析每一个环境因子对互花米草根茎苗定植的影响,辨识影响互花米草根茎苗入侵潮滩生境的关键环境因子,研究结果将为互花米草的入侵机理研究、风险评估与防控管理提供理论依据.
1 研究区域
上海南汇东滩湿地(30°50′04″~3l°Ol′00″N,121°50′50″~122°01′00″E)位于長江口和杭州湾海岸交汇处,受非正规半日潮影响,多年平均潮差2.67m左右,属于北亚热带海洋性气候,温和湿润,年均温15~16℃[25-26].在南汇东滩北部和南部选择两处不同类型潮滩的光滩(无植被覆盖)作为实验地点,其位置如图1所示,北部潮滩实验地点为砂质滩涂,沉积物黏着性较强.南部潮滩实验地点为沉积物粒径较粗的粉砂质潮滩,是典型的“铁板砂”滩涂.
2 研究方法
2.1 互花米草移栽样点布设
在南汇东滩的北部潮滩和南部潮滩,由陆向海方向各布设5个移栽样点,其中1个设置在消浪堤内,4个设置在消浪堤外,由此代表由陆向海的5种环境梯度(见图1和图2a).2017年5月4-5日(小潮期),在南汇东滩就近带土挖取高度一致(18±2) cm、长势健康的互花米草根茎苗(根系土壤厚度>20 cm),并按1株/m2、3株/m2、5株/m2、10株/m2和20株/m2的源株密度随机移栽到每个样点的固定样方内(1 mxl m),以此代表不同繁殖体大小的根茎苗入侵潮滩生境.每个密度设3个重复,样方与样方之间间隔2m(见图2a和图2b).
2.2 野外监测及数据处理
(1)高程监测2017年4月17日,利用三维激光扫描仪(RIEGL Laser Measurement Sys-tems,Riegl V2-4000)和实时动态定位系统(Real-Time Kinematic Global Position System,RTK-GPS)获取移栽样地的地形高程,高程基准面为吴淞高程.在ArcGIS中生成两处移栽样地的数字高程模型(Digital Elevation Model,DEM).
(2)水动力监测2017年4月21 - 23日小潮期,在每个移栽样点架设电磁式流速计(Electromagnetic Current Meter,ALEC)和RBR波潮仪(Tide Wave Recorder-2050,TWR-2050)监测水体流速和波能密度,观测时将仪器固定在木桩上,保证传感器距离滩面均为5 cm.
(3)植物监测分别在移栽后第1天、第7天、第14天及第30天监测并记录互花米草根茎苗的定植及存活状况,在第14天互花米草根茎苗存活率趋于稳定,第30天结束监测.
2.3 野外采样及样品处理
(1)沉积物粒度测定2017年5月11-12日在每个移栽样方中使用100 cm3环刀采集表层沉积物样品,带回实验室通过激光粒度仪(MS-2000)获取沉积物的粒度信息和泥沙组分,采用谢帕德(Shepard)方法命名沉积物[27-28],依据砂、粉砂和粘土的百分比含量,绘制沉积物的谢帕德分类三角图.
(2)沉积物临界剪切应力测定参照Xu等[29]的方法,使用10 cm直径玻璃管采集表层20 cm沉积物样品,上下两侧加保鲜膜和PVC管帽密封,带回实验室使用U-GEMS微观侵蚀系统测量北部潮滩和南部潮滩各移栽样点沉积物的临界剪切应力.
2.4 统计分析
在SPSS软件中运用独立样本t检验(Independent-Samples T Test)和单因素方差分析(One-Ways ANOVA)比较北部潮滩和南部潮滩及N1-N5、S1-S5各样点间环境特征的差异,以及不同环境梯度(由陆向海的5个移栽样点)对互花米草繁殖体存活率的影响.
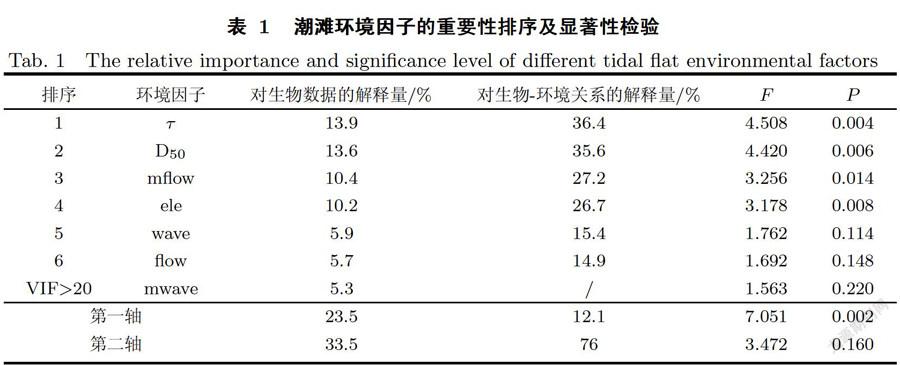
利用Canoc04.5软件RDA分析方法研究潮滩环境因子对互花米草根茎苗定植的相对重要性[30-31].以互花米草繁殖体的存活率指标作为生物变量,以高程(elevation,ele)、平均流速(mean fiow velocity, flow)、最大流速(maximum flow velocity, mflow)、平均波能密度(mean wave energy,wave)、最大波能密度(maximum wave energy,mwave)、临界剪切应力(critical shear stress,丁)和中值粒径(median diameter,Dso)作为环境变量.所有环境变量均经过log(l+x)转换以消除量纲的影响.分析过程中,剔除解释量低且膨胀因子(VarianceInflation Factor,VIF)大于20的环境变量,使用Manual Selection分析环境因子对生物变量的影响,并通过蒙特卡罗检验得到环境因子的重要性排序.
3 结果分析
3.1 南汇东滩自然潮滩环境特征
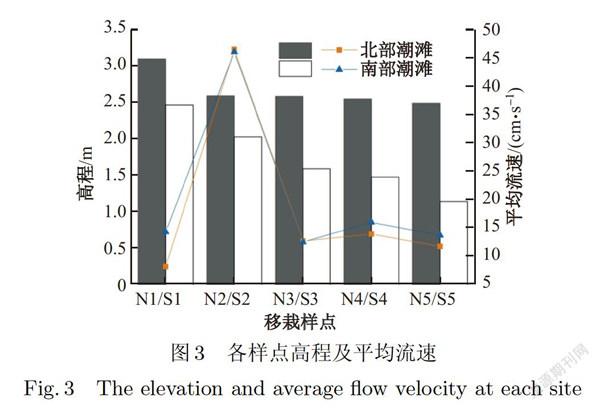
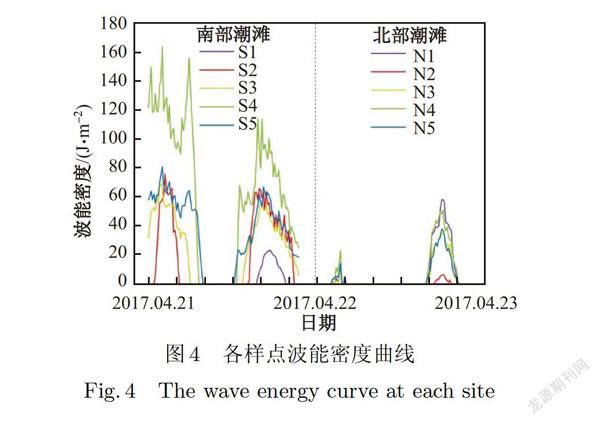
南汇北部潮滩及南部潮滩的地形高程均呈现由陆向海逐渐降低的趋势,但南部潮滩的高程显著低于北部潮滩(见图3,P<0.05).南北部潮滩平均流速无显著差异(P>0.05)(见图3),消浪堤外第一个样点(N2和S2)的流速均显著高于其他样点(P<0.05).此外,由图4可知,南部潮滩的波能密度大于北部潮滩(P<0.05).
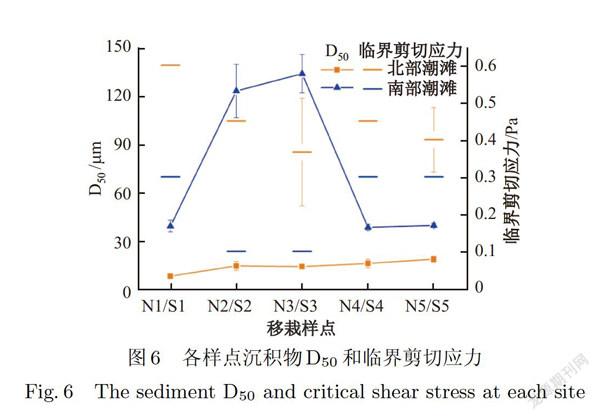
由图5和图6可知,北部潮滩沉积物中值粒径(median diameter,Dso)变化范围为6.5~20.3 μm,底质为砂和粘土质砂,属于砂质潮滩;南部潮滩沉积物Dso变化范围为36.2~147.O μm,底质为粉砂质砂和砂质粉砂,属于粉砂质潮滩.南部潮滩所有样点的沉积物Dso均显著高于北部潮滩(见图6,P<0.05).在北部潮滩,消浪堤内(NI)的沉积物Dso与消浪堤外的样点(N2N5)有显著差异(P<0.05),在分类上消浪堤内的沉积物属于粘土质砂,而消浪堤外的属于砂(见图5).在南部潮滩上,S2、S3处的沉积物D50高于其他样点(见图6,P<0.05),在分类上这两个点的沉积物属于砂质粉砂,而南部潮滩其他样点的沉积物属于粉砂质砂(见图5).南部潮滩的临界剪切应力显著低于北部潮滩(见图6,P<0.05),表明北部潮滩滩面的抗侵蚀能力高于南部潮滩.综合来看,南汇东滩的南部潮滩沉积物异质性较强.更易受到侵蚀,
3.2 互花米草繁殖体存活特征
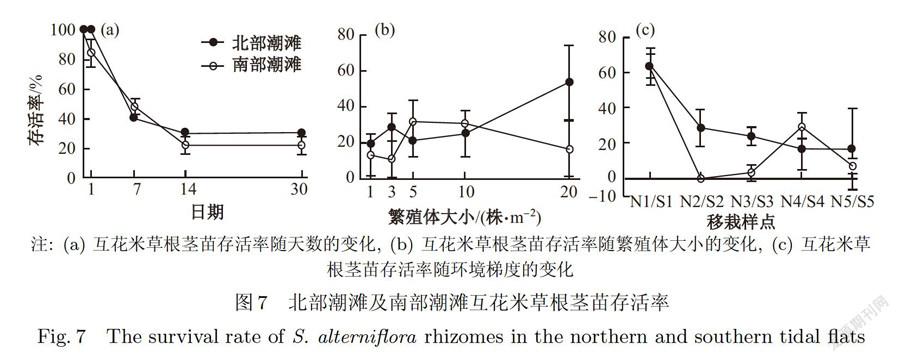
从图7a可以看出,两个实验地点的互花米草繁殖体在移栽后的两周内存活率均急剧下降,到第14天开始趋于稳定,北部互花米草存活率为(29.8±1.5)%,略高于南部的(21.7±5.6)% (P>0.05).第30天,北部潮滩和南部潮滩分别仍保持(29.8±1.5)%和(21.7±5.6)%的存活率,但差异不显著(P>0.05).
在南北两处潮滩上,互花米草根茎苗的存活率都随繁殖体大小和环境梯度(樣点距大堤距离)的变化而变化(见图7b和图7c)在北部砂质潮滩上,互花米草存活率整体上随繁殖体株数的增加而增大,但3株/m2互花米草斑块的存活率也较高,为(28.9±7.7)%,仅次于20株/m2繁殖体(53.7±20.8)%的存活率;在南部粉砂质潮滩上,互花米草存活率随繁殖体株数的增加整体上呈现先增大后减小的趋势,但3株/m2互花米草斑块的存活率最小,为(11.1±10.2)%,5株/m2互花米草斑块的存活率最大,为(32.0±12.0) %.环境梯度对互花米草的存活率存在显著影响(P<0.05).在北部砂质潮滩上,由陆向海繁殖体的存活率依次递减,存活率最大为(63.4±10.6)% (N1,消浪堤内),最小为(16.4±23.3)% (N5,消浪堤外);在南部砂质潮滩上,消浪堤内(SI)互花米草存活率最大,为(64.0±6.9)%,消浪堤外仅在S4处能保持(29.8±7.1)%存活率,其余各点接近或等于0.
3.3 影响互花米草繁殖体存活的关键环境因子分析
3.3.1 环境因子对互花米草定居的RDA分析
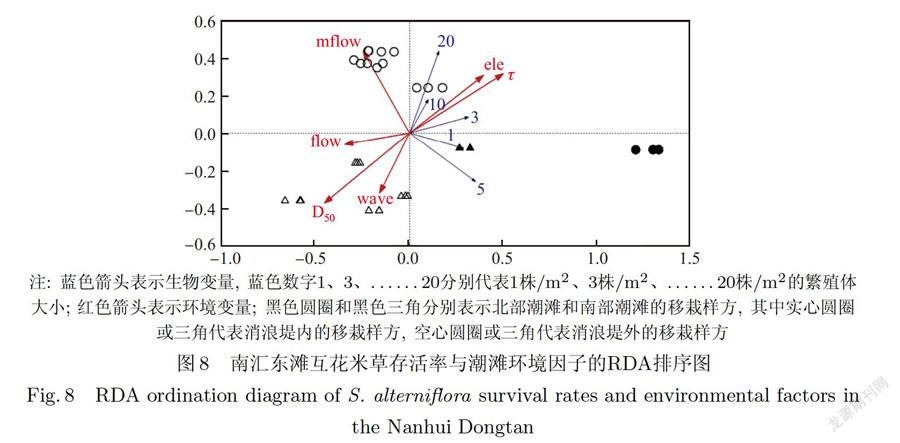
当不考虑南北两个潮滩差异,对整个南汇东滩生物一环境数据进行RDA分析时,最大波能密度的膨胀因子大于20且解释量较低,因而被剔除.剩余6个潮滩环境因子对互花米草繁殖体存活的影响程度由大到小为临界剪切应力、中值粒径、最大流速、高程、平均波能密度和平均流速(见表1).其中临界剪切应力、中值粒径和高程对互花米草繁殖体存活的影响达到极显著水平(P<0.01),是影响互花米草根茎苗定植的主导环境因子.临界剪切应力和高程与不同大小繁殖体的存活率呈正相关关系,中值粒径与不同大小繁殖体的存活率呈负相关关系(见图8).滩面临界剪切应力越大说明沉积物的抗侵蚀性能越强,互花米草繁殖体入侵光滩后越难以被冲走;高程越高的地点淹水深度、时间等环境胁迫与干扰越小,互花米草繁殖体的存活率越高;沉积物中值粒径与临界剪切应力存在负相关关系[32],中值粒径越大,土壤结构越松散,越不利于互花米草定居.最大流速对互花米草繁殖体存活的影响也达到了显著水平(P<0.05),也是影响互花米草根茎苗定植的主导环境因子,它对互花米草繁殖体存活率的影响与繁殖体的大小有关,当互花米草植丛较大时(20株/m2和10株/m2)可以抵御更大的流速.平均流速和平均波能对互花米草繁殖体存活的影响并不显著(P>0.05).此外,RDA排序图(见图8)能很好地区分潮滩消浪堤内、外的样方,其中消浪堤内的样方主要分布在与存活率有正相关关系环境变量箭头的正方向上,这说明消浪堤对互花米草根茎苗的定植有一定保护作用.
3.3.2 北部潮滩环境因子RDA分析结果
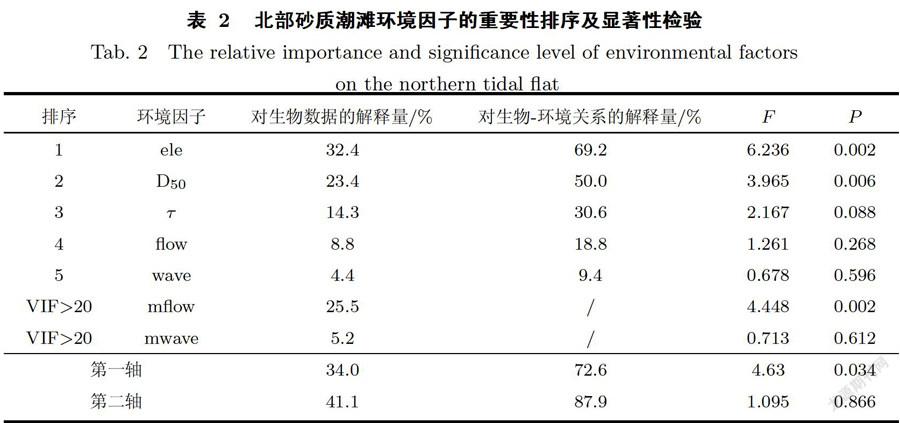
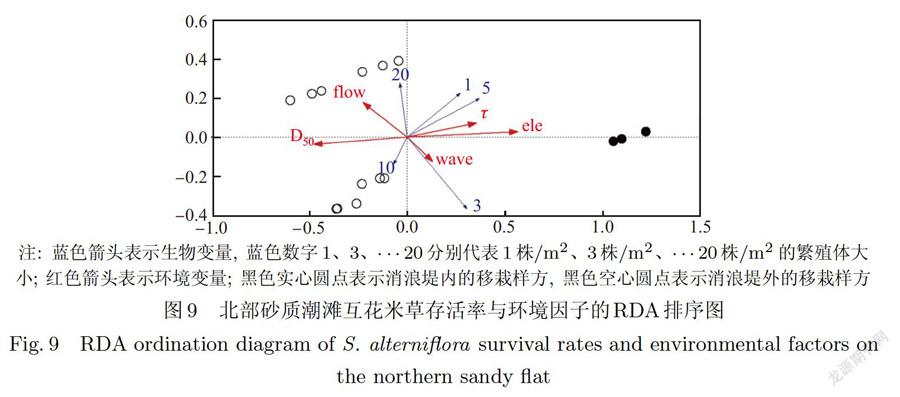
北部潮滩为砂质潮滩,对它的生物一环境数据进行RDA分析时,最大流速和最大波能密度的膨胀因子大于20,故剔除.结果显示,在北部砂质潮滩上,影响互花米草繁殖体存活的环境因子由大到小依次为高程、中值粒径、临界剪切应力、平均流速和平均波能密度(见表2).其中只有高程和中值粒径达到显著水平(P<0.01),是互花米草根茎苗定植的主导环境因子.当互花米草繁殖体密度较低时(1株/m2、3株/m2和5株/m2),繁殖体存活率与高程呈正相关,与中值粒径呈负相关;当繁殖体较大(10株/m2和20株/m2)时,繁殖体存活率与高程呈负相关,与中值粒径呈正相关(见图9).
RDA排序结果表明(见图9),中值粒径的箭头方向与高程相反,说明在北部砂质潮滩,沿高程降低的方向中值粒径逐渐变大,沉积物颗粒逐渐变粗.高程高的地点对低密度的互花米草斑块(1株/m2、3株/m2和5株/m2)有相对强的保护作用;在高程低的地点环境干扰与胁迫相对更为强烈,较大的互花米草植丛(10株/m2和20株/m2)存活能力相对较强;当植丛为20株/m2时受高程和中值粒径的影响较小,体现了其对环境干扰具有较强的抵御能力.
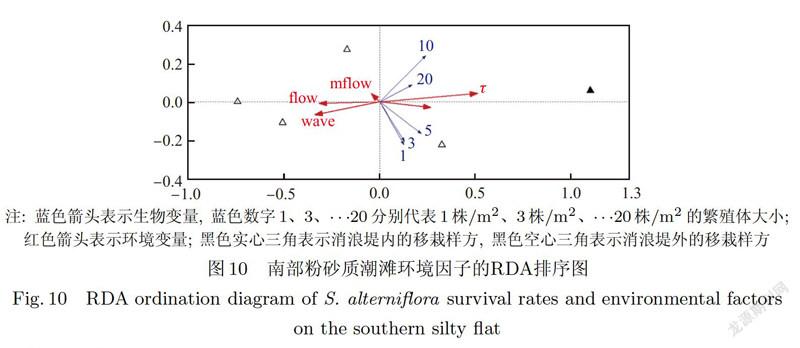
3.3.3 南部潮滩环境因子RDA分析结果
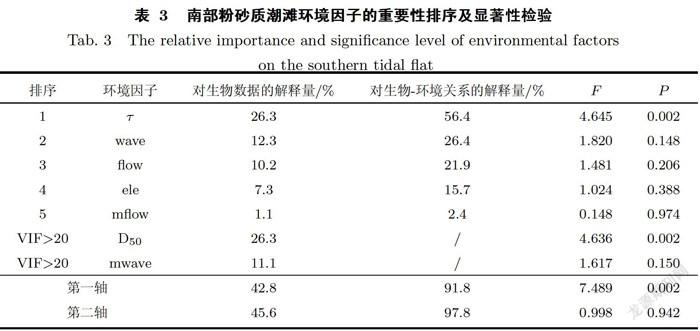
南部潮滩为粉砂质潮滩,对其生物一环境数据进行RDA分析时,剔除中值粒径和最大波能密度两个膨胀因子大于20的变量.RDA结果显示,环境因子的重要性由大到小依次为临界剪切应力、平均波能密度、平均流速、高程和最大流速(见表31.其中仅临界剪切应力对互花米草根茎苗定植的影响达到显著水平(P
4 讨论
4.1 潮滩环境因子對互花米草定植的影响
在本研究中,滩面临界剪切应力、中值粒径、最大流速和高程都是影响互花米草定植的重要环境因子.滩面临界剪切应力表征了沉积物的抗侵蚀能力,当临界剪切应力小于波、流产生的联合剪切应力时,滩面将会发生侵蚀[32],定居植物面临冲刷的威胁[24,33],这将不利于潮滩植物的定居和植被的保留.中值粒径反映了地貌的基本特征[34]和水动力环境[35],潮滩沉积物中值粒径越大,土壤结构更松散,使得入侵植物越易被冲刷走,不利于其定居.最大流速表征了潮水流速的极值情况,代表了潮水可达到的最强动力,在河口复杂的水文环境下,频繁的潮水干扰会使得互花米草植株在尚未形成足够强大的根系网络时就被冲走,造成定居的失败[12-13].高程对互花米草入侵的影响已有大量研究[36-38],高程因子关系着淹水程度(淹水深度、淹水时间、淹水频率)、风浪大小、土壤条件和地形冲淤条件等[33],环境因子沿高程梯度也呈一定梯度变化,通常高程越高,环境条件越好,越有利于互花米草根茎苗定植.
研究结果还表明,当潮滩类型不同时,影响互花米草成功定居的主导影响因子也不同.本研究中,在北部砂质潮滩,对互花米草根茎苗定植起主导作用的环境因子为高程和中值粒径(P<0.05).高程较高的区域水动力相对较弱,有利于低密度的互花米草斑块(1株/m2、3株/m2和5株/m2)存活,但随着高程降低、中值粒径增大,较大的互花米草植丛(10株/m2和20株/m2)更易定居存活,这主要是因为随着高程降低,环境胁迫增大,大斑块的互花米草植丛可通过“抱团”效应以抵抗环境干扰与胁迫,使其更利于在较强水动力条件下定居[39].在南部粉砂质潮滩,对互花米草根茎苗定植有主导影响的环境因子为临界剪切应力(P<0.05).这主要是因为粉砂质类型的潮滩沉积物粒径较大,通常该区域水动力过程也较强,潮滩的临界剪切应力较低,抗冲刷的能力相对更弱[40],不利于互花米草定植,因此临界剪切应力对互花米草定居发挥着最重要的作用.本研究中南部潮滩互花米草移栽样点的临界剪切应力均值为0.22 Pa,而朱琴[40]在南汇的野外观测及模型研究发现南汇嘴南滩波流联合的床底剪切应力均值约为0.25 Pa,表明南部潮滩实验区临界剪切应力小于波流联合剪切应力,潮滩主要处于侵蚀状态,不利于互花米草定居.
本研究中,北部潮滩和南部潮滩互花米草存活率曲线存在明显不同,这主要是因为南北两条断面在地貌类型、水动力过程等环境特征方面存在明显差异(见图7c)首先,两处潮滩的地貌类型显著不同.北部潮滩属于砂质潮滩,南部潮滩属于粉砂质潮滩,其所有样点的沉积物中值粒径均显著高于北部潮滩(见图6,P<0.05),这意味着南部潮滩的沉积物更松散,互花米草不易在此扎根,所以存活率相对更低.其次,南部潮滩的水动力(例如波能密度)显著大于北部潮滩(见图4,P<0.05),波浪的运动会对互花米草植株形成拖曳,冲走互花米草根茎苗.第三,南部潮滩的临界剪切应力显著低于北部潮滩(见图6,P<0.05),表明南部潮滩滩面的抗侵蚀能力低于北部潮滩,同时观测数据也表明南部潮滩整体处于冲刷状态,互花米草更难以保留.
4.2 互花米草入侵的管控建议
互花米草是世界广布的入侵植物,入侵范围包括中国沿海、北美西海岸、法国、英国、西班牙、澳大利亚、新西兰和南非[2-3].近年来,互花米草的治理工作在各地开展[3,41-42],但由于互花米草强大的生长繁殖能力、环境特征的异质性及治理措施的局限性,彻底根除互花米草很难实现,一些地区甚至在治理后出现了二次入侵问题[3,43],加剧了互花米草的入侵风险和管控难度.
本研究中,北部砂质潮滩和南部粉砂质潮滩的主导环境因子有所区别,这意味着在控制互花米草扩散和评估互花米草入侵风险时,需要充分考虑潮滩类型的差异.在北部砂质潮滩上影响互花米草入侵的主导环境因子是高程和中僮粒径.高程较高的地点有利于互花米草入侵,这与已有研究结果一致[37].此外,滩面沉积物的性质也会影响盐沼植物的定居能力[11,13].因此,砂质光滩上高程较高、土壤中值粒径较小的区域应被视为互花米草管控的重点区域,需加强观测、管控与风险评估,以防互花米草肆意蔓延.在南部粉砂质潮滩上,主导环境因子为临界剪切应力.临界剪切应力与潮滩冲淤密切相关,临界剪切应力越大,滩面越不容易受到侵蚀,滩面植物越易于保留[24].目前,滩面临界剪切应力与互花米草定植的关系研究较少,多数研究关注于波流联合的剪切应力[10,18],而不是滩面的抗侵蚀能力.潮滩侵蚀会使光滩上的植物难以保留,不利于互花米草入侵[24];而一定程度的淤积为盐沼植被提供了新生生境,有利于互花米草入侵[20].因此,粉砂质光滩的抗侵蚀能力及侵蚀淤积状况需要得到关注,以便对互花米草入侵进行管控与预警.
在我国海岸带,潮滩类型多样[44],对盐沼植物产生扰动作用的环境因子也多样[13,17],综合研究不同类型潮滩、不同环境因素对互花米草入侵的影响,厘清影响互花米草入侵的主导环境因子,将有助于有效开展互花米草防控及准确评估互花米草的入侵风险,提高互花米草入侵的管控效率,对滨海湿地生态系统稳定及可持续发展具有重要意义.
[参考文献]
[1] DAEHLER c c,STRONG D R.Status, prediction and prevention of introduced cordgrass Spartina spp. invasins in Pacific estuaries, USA [J]. Biological Conservation, 1996, 78(1/2): 51-58.
[2] 王卿,安树青,马志军,等.入侵植物互花米草——生物学、生态学及管理[J].植物分类学报,2006, 44(5): 559-588.
[3]RIDDIN T, VAN WYK E, ADAMS J The rise and fall of an invasive estuarine grass [J]. South African Journalof Botany, 2016, 107: 74-79.
[4] 赵彩云,李俊生,柳晓燕.中国主要外来入侵物种风险预警与管理[M].北京:中国环境科学出版社,2016.
[5] 宫璐,李俊生,柳晓燕,等中国沿海互花米草遗传多样性及其遗传結构[J].草业科学,2014, 31(7): 1290-1297.
[6] 邓自发,安树青,智颖飙,等.外来种互花米草入侵模式与爆发机制[J].生态学报,2006, 26(8): 2678-2686.
[7] 陈中义,李博,陈家宽.米草属植物入侵的生态后果及管理对策[J]生物多样性,2004, 12(2): 280-289.
[8] 李郑杰.漳江口红树林区互花米草入侵及扩散机制研究[D].福建厦门:厦门大学,2014.
[9] 赵彩云,柳晓燕,白加德,等,广西北海西村港互花米草对红树林湿地大型底栖动物群落的影响[J].生物多样性,2014, 22(5):630-639
[10]BALKE T, BOUMA T J,HORSTMAN E M, et al. Windows of opportunity: thresholds to mangrove seedlingestablishment on tidal flats [Jl. Marine Ecology Progress Series, 2011, 440(1): 1-9.
[11]SCHWARZ C, BOUMA T J,ZHANG L Q, et al. Interactions between plant traits and sediment characteristicsinfluencing species establishment and scale-dependent feedbacks in salt marsh ecosystems [J]. Geomorphology,2015, 250: 298-307.
[12] 李蕙,袁琳,张利权,等.长江口滨海湿地潮间带生态系统的多稳态特征[J].应用生态学报,2017, 28(1): 327-336.
[13]SCHWARZ C: YSEBAERT T, ZHU z,et al. Abiotic factors governing the establishment and expansion of twosalt marsh plants in the Yangtze Estuary, China [J]. Wetlands, 2011, 31(6): 1011-1021.
[14] 王卿,汪承焕,黄沈发,等.盐沼植物群落研究进展:分布、演替及影响因子[J].生态环境学报,2012, 21(2): 375-388.
[15] 闫芊,陆健健,何文珊.崇明东滩湿地高等植被演替特征[J].应用生态学报,2007, 18(5): 1099-1103
[16] ZHU z,ZHANG L,WANG N, et al. Interactions between the range expansion of saltmarsh vegetation andhydrodynamic regimes in the Yangtze Estuary, China [Jl. Estuarine, Coastal and Shelf Science, 2012, 96(1):273-279.
[17]CAO H, ZHU z,BALKE T, et al. Effects of sediment disturbance regimes on Spartina seedling establishment:Implications for salt marsh creation and restoration [J]. Limnology and Oceanography, 2018, 63(2): 647-659.
[18]HU z,VAN BELZEN J, VAN DER WAL D, et al. Windows of opportunity for salt marsh vegetation establish-ment on bare tidal flats: The importance of temporal and spatial variability in hydrodynamic forcing [J]. Journalof Geophysical Research Biogeosciences, 2015, 120(7): 1450-1469.
[19] BALKE T, HERMAN P M J,BOUMA T J.Critical transitions in disturbance-driven ecosystems: Identifyingwindows of opportunity for recovery [J]. Journal of Ecology, 2014, 102(3): 700-708
[20] 曹浩冰,葛振鸣,祝振昌,等.崇明东滩盐沼植被扩散格局及其形成机制[J].生态学报,2014, 34(14): 3944-3952.
[21] 沙晨燕,李玲,唐浩,等.典型灘涂环境因子对植物群落空间分布的影响[J].人民长江,2016, 47(22): 10-15.
[22] CALLAGHAN D P,BOUMA T J,KLAASSEN P,et al. Hydrodynamic forcing on salt-marsh development:Distinguishing the relative importance of waves and tidal flows [Jl. Estuarine Coastal and Shelf Science, 2010,89(1): 73-88.
[23] VANDENBRUWAENE W, TEMMERMAN s,BOUMA T J,et al. Flow interaction with dynamic vegetationpatches: Implications for biogeomorphic evolution of a tidal landscape [J]. Journal of Geophysical Research:Earth Surface, 2011, 116(F1): 155-170.
[24]FAGHERAZZI s,CARNIELLO L,ALPAOS L D, et al. Critical bifurcation of shallow microtidal landforms intidal flats and salt marshes [J]. Proceedings of the National Academy of Sciences of the United States of America,2006, 103(22): 8337-8341.
[25] 刘曙光,郁微微,匡翠萍,等.三峡工程对长江口南汇边滩近期演变影响初步预测[J].同济大学学报(自然科学版),2010, 38(5):679-684.
[26] 陈万逸,张利权,袁琳.上海南汇东滩鸟类栖息地营造工程的生境评价[J].海洋环境科学,2012, 31(4): 561-566.
[27] 蒋丰佩,何青,张国安,等.异质潮滩波浪衰减特性研究——以长江口崇明东滩为例[J].泥沙研究,2013(1): 45-52.
[28] 薛春汀.两种碎屑沉积物分类的比较[J].海洋地质与第四纪地质,1994, 14(2):109-113.
[29] xu K, CORBETT D R, WALSH J P,et al. Seabed erodibility variations on the Louisiana continental shelfbefore and after the 2011 Mississippi River flood [J]. Estuarine, Coastal and Shelf Science, 2014, 149: 283-293.
[30] 肖石红,张中瑞,覃德华,等.天宝岩国家级自然保护区4种类型泥炭藓沼泽植被组成及其与环境因子的关系[J]应用与环境生物学报,2016, 22(4): 631-638.
[31] 黄小荣,何峰,庞世龙,等.广西石山人工林灌草多样性与环境因子的关系[J].生态学杂志,2015, 34(11): 3024-3033.
[32] 郑杰文,贾永刚,刘晓磊,等.现代黄河三角洲沉积物临界剪切应力研究[J]海洋学报,2015, 37(3): 86-98.
[33] 张利权,袁琳.基于生态系统的海岸带管理[M].北京:海洋出版社,2012.
[34] 刘红,何青,吉晓强,等.波流共同作用下潮滩剖面沉积物和地貌分异规律——以长江口崇明东滩为例[J].沉积学报,2008,26(5): 833-843.
[35] 傅开道,杨文辉,苏斌,等.流域环境变化的河流沉积物粒度响应——澜沧江案例[J].地理科学进展,2015, 34(9): 1148-1155.
[36]GE z M. WANG H, CAO H B. et al. Responses of eastern Chinese coastal salt marshes to sea-level rise combinedwith vegetative and sedimentary processes [J]. Scientific Reports, 2016, 6(1): 28466-28475.
[37] 陈中义,李博,陈家宽.长江口崇明东滩土壤盐度和潮间带高程对外来种互花米草生长的影响[J].长江大学学报(自然科学版),2005, 2(2): 6-9.
[38] 刘会玉,林振山,齐相贞,等.基于个体的空间显性模型和遥感技术模拟入侵植物扩张机制[J].生态学报,2015, 35(23):7794-7802.
[39] 赵志远,袁琳,李伟,等生境异质性及源株密度对互花米草入侵力的影响[J]生态学报,2018, 38(18): 6632-6641
[40]ZHU Q, YANG s, MA Y. Intra-tidal sedimentary processes associated with combined wavecurrent action on anexposed, erosional mudflat. southeastern Yangtze River Delta. China [J]. Marine Geology, 2014: 347(2): 95-106.
[41]YUAN L, ZHANG L, XIAO D, et al. The application of cutting plus waterlogging to control Spartina alternifZoraon saltmarshes in the Yangtze Estuary, China [J]. Estuarine, Coastal and Shelf Science, 2011, 92(1): 103-110.
[42] 赵相健,柳晓燕,宫璐,等.刈割加遮荫综合治理互花米草(Spartina alterniflora) [J].生态学杂志,2014: 33(10): 2714-2719.
[43] 肖德荣,祝振昌,袁琳,等.上海崇明东滩外来物种互花米草二次入侵过程[J]应用生态学报,2012, 23(11): 2997-3002.
[44] 邸向红,侯西勇,吴莉中国海岸带土地利用遥感分类系统研究[J].资源科学,2014, 36(3): 463-472.
(责任编辑:李万会)
收稿日期:2018-09-14
基金项目:国家自然科学基金项目(41876093);上海市科委科研计划项目(17D21201902,
18D21204802,18D21206506);国家重点研发计划项目(2016YFE0133700)
第一作者:朱晓泾,女,硕士研究生,研究方向为湿地生态学.
通信作者:袁琳,女,副研究员,研究方向为湿地生态学.E-mail: lyuan@sklec.ecnu.edu.cn.
