温度变化对南极假山毛榉光合系统的影响
2019-09-05郝向春韩丽君
郝向春,周 帅,翟 瑜,韩丽君
(山西省林业科学研究院,山西 太原 030012)
环境温度对植物的生理生化活动具有深刻影响,是决定植物生长发育和分布范围的关键因素。适宜的温度能使植物的光合效率最大化,加快有机物的合成与运输,更高效地吸收水肥;低温胁迫则会导致细胞器受损、细胞组织脱水、膜脂相变、活性氧(Reactiveoxygen species;ROS)大量产生、以及多糖等大分子物质降解或变性[1];高温胁迫则对细胞膜、细胞骨架、蛋白质、酶和RNA的结构与功能产生负面影响[2]。研究表明,光合作用是对温度变化最敏感的生理进程之一[2],高低温胁迫能加速光合色素降解速率[1,3],对光反应和碳代谢均能造成较大伤害[2-5]。高低温胁迫产生的大量ROS会造成光系统II(PhotosystemII;PSII)蛋白功能紊乱和脂质过氧化[6],这使得PSII成为光合系统中对胁迫最敏感的组分[2,7]。近年来,利用非损伤、检测时间短、灵敏度高的叶绿素荧光分析技术[8],研究发现冻害胁迫(零下低温)主要导致类囊体膜的光合反应能力受损,主要作用部位位于放氧复合体(Oxygen-releasing complex;OEC)、PSII和光系统I(PhotosystemI;PSI);冷害胁迫(零上低温)主要降低PSII反应速率,并抑制D1蛋白合成[5,7];高温则导致Rubisco酶、OEC、CF0-CF1 ATP酶的活性降低[9]。快速光响应曲线(Rapid light curve;RLC)常被用来研究环境变化对光合作用光化学反应的影响[10-11],也可作为一种快速检测手段对胁迫下植株的光合能力进行评估[12],已经在植物非生物胁迫分析中广泛使用[10],相关研究发现高低温胁迫均能导致经RLC推导出的多项参数显著下降[13-14]。作为光合系统的物质基础——叶绿素对高低温胁迫较为敏感,多数植物的叶绿素合成最适温度是20~30 ℃,低于2 ℃或高于40 ℃均会导致叶绿素的降解速率高于合成速率[3]。同时对高低温胁迫下植株的叶绿素荧光、快速光响应曲线和叶绿素含量进行观测,既可以对胁迫下光合机构的结构性损伤进行评估,也可以对胁迫导致的光反应能量分配的功能性改变进行分析,并能综合评估光合系统结构和功能的改变对叶片光合能力的影响。
南极假山毛榉Nothofagus antarctica为壳斗科Fagaceae假山毛榉属Nothofagus落叶乔木或灌木,主要分布在南美洲南部的温带森林[15]。当前国际上对林草复合系统中南极假山毛榉的生存状态、南极假山毛榉的自交不亲和与种间杂交现象,以及其在不同环境中表现的形态变化和采取的生存适应策略等方面进行了大量研究[16],发现南极假山毛榉具有环境适应性强[17-19]、观赏价值高[20]、经济价值大[21]、基因库丰富[22-23]等优势,因此对其国际间引种也日益增多[15]。但现有研究尚缺乏对南极假山毛榉温度耐受性的研究,仅见对其能耐受的极端低温的报道[24],也没有针对不同温度下南极假山毛榉光合系统变化的系统研究。本研究采用人工模拟高低温胁迫的方法,观测不同温度处理下南极假山毛榉叶片的叶绿素荧光、RLC和相对叶绿素含量(Relative chlorophyll content;SPAD)的变化,以期为南极假山毛榉培育温度的设置提供科学依据,完善南极假山毛榉的引种驯化技术体系。
1 材料与方法
1.1 试验材料与处理方法
南极假山毛榉实验种子于2016年采自阿根廷内乌肯省(38°52′S,71°14′W,海拔1 240 m),试验植株由山西省林业科学研究院繁育。选取长势良好且一致的10株盆栽1年生苗,于2018年7月连盆土置于25℃的人工气候箱(光照时间,6:00—18:00,12 h·d-1;光照强度,100 μmol·m-2s-1;相对湿度,60%±3%)中适应培养3 d。利用人工气候箱和装有照明设备的冰柜模拟不同环境温度,设置-5、5、15、25、30、35和40 ℃共7个温度处理,每个温度处理适应24 h后开始各项测试,处理步骤如下。
降温处理:选择5株植株放入预先调至25 ℃的人工气候箱,然后以5 ℃/h的速度依次降到各处理温度,直至-5 ℃。-5 ℃处理在冰柜中完成。
升温处理:选择另外5株植株放入预先调至25 ℃的人工气候箱,然后以5 ℃/h的速度依次升到各处理温度,直至40 ℃。
每项测试在各处理环境中进行,均选择每株的同一片成熟叶片测试,每项测试每株测量1次,重复5次。测试按照快速叶绿素荧光诱导动力学测定、稳态叶绿素荧光动力学测定、快速光响应曲线测定、叶绿素含量测定的步骤开展,以25 ℃的温度处理为对照(CK)。
1.2 指标调查与分析方法
1.2.1 快速叶绿素荧光诱导动力学的测定
将各温度处理的叶片暗适应20 min后,采用叶绿素荧光仪(Yaxin-116G,雅欣理仪公司,中国),选取叶肉部分,用 3 000 μmol·m-2s-1饱和蓝闪光照射1 s,以10 μs(300 μs之前)、100 μs(300 μs ~3 s之间)和1 ms(3 ms之后)的间隔记录荧光信号,测得叶绿素荧光动力学参数。暗适应后,PSII最大光化学效率Fv/Fm、单位面积反应中心数量RC/CS、单位反应中心复合体吸收的能量ABS/RC、单位反应中心复合体中被核心捕获的能量TRo/RC、单位反应中心捕获的用于电子传递的能量ETo/RC、捕获的激子能导致电子传递的效率ψo、电子传递的量子产额ΦEo、叶片性能指数PIABS等参数计算见文献[9]。
1.2.2 稳态叶绿素荧光动力学的测定
将各温度处理的叶片暗适应20min后,采用前述叶绿素荧光仪对各温度处理的叶片进行稳态荧光测量,选取叶肉部分,按照饱和脉冲光(4 000 μmol·m-2s-1,持 续1 s)—远 红 光(10 W·m-2,持续6 s)—黑暗(持续4 s)—光 化 光(1 500 μmol·m-2s-1,持 续9 s)的 顺序,连续诱导7个周期,以10 μs(300 μs之前)、100 μs(300 μs ~3 ms之间)、1 ms(3 ms~300 ms之间)、100 ms(300 ms之后)的间隔记录荧光信号,相关叶绿素荧光参数由仪器自动给出。光化学猝灭系数qP、非光化学猝灭系数qN等参数计算见文献[25]。PSII用于天线色素热耗散的能量比例D、用于光化学反应的能量比例P、非光化学反应耗散能量比例E等参数计算见文献[26]。PSI荧光强度F1计算见文献[27],总荧光强度中PSI荧光强度的占比C=F1/F0,F0表示暗处理后的最小荧光产量。
1.2.3 快速光响应曲线的测定
使用便携式调制式叶绿素荧光仪(PAM-2500,Walz,德国)对光适应1 h的各温度处理叶片的叶肉部分进行光响应曲线测量,光强梯度设定为0、20、50、100、300、600、900、1 200、1 500、1 800、2 000 μmol·m-2s-1,每个梯度光照持续时间20 s,之后利用仪器配套软件PamWin_3的曲线拟合功能[28]获得相关参数。α、ETRmax、IK分别表示光能利用效率、最大电子传递速率和半饱和光强。
1.2.4 叶绿素含量的测定
采用叶绿素测定仪(SPAD-502Plus,KONICA MINOLTA,日本)测定各温度处理叶片的叶肉部分的SPAD。
1.2.5 数据分析统计
数据统计分析采用Excel 2010和SPSS 22.0,非线性回归拟合和绘图采用Origin2018。
2 结果与分析
2.1 温度对快速叶绿素荧光诱导动力学曲线的影响
经过暗适应后转入饱和脉冲光强照射,南极假山毛榉叶片的叶绿素荧光迅速上升,经过O、J、I、P各点后逐渐平稳,其中-5 ℃处理的荧光强度最高,15 ℃和30 ℃处理的荧光强度明显偏低(图1-Ⅰ)。经过标准化处理,可见各条曲线在J点有明显分散,低温胁迫(-5、5和15 ℃等处理,下同)的荧光强度高于高温胁迫(30、35和40 ℃等处理,下同),CK的荧光强度最低(图1-Ⅱ)。将标准化的各处理曲线与CK相减,可见高温胁迫下,J、I点的荧光强度随着温度升高逐渐增加;低温胁迫诱导J、I点的荧光强度由大到小依次是-5、15、5 ℃(图1-Ⅲ)。高温胁迫下,K点的荧光强度随温度上升而逐渐明显;低温胁迫下,只有-5 ℃和5 ℃的K点较明显(图1-Ⅳ)。高低温胁迫下均能观察到L点出现,并且随着低温胁迫的加剧,L点愈加明显(图1-Ⅴ)。
2.2 温度对叶绿素荧光动力学参数的影响
温度变化导致多项南极假山毛榉叶片的叶绿素荧光动力学参数在25 ℃处理时达到最大值,-5 ℃或40 ℃处理时降低到最小值。
高低温胁迫对PSⅡ反应中心复合体能量的吸收和传递均构成负面影响。高温胁迫下,PSII单位反应中心复合体吸收的能量ABS/RC和单位反应中心复合体中被核心捕获的能量TRo/RC以及单位反应中心捕获的用于电子传递的能量ETo/RC均呈现下降趋势,40 ℃胁迫时分别比CK下降了8.6%、9.1%(P< 0.05)和17.1%(P< 0.05)。低温胁迫导致ETo/RC持续显著下降,-5 ℃胁迫时仅为CK的80.6%(P< 0.01)。低温胁迫对量子产额或流量比的抑制比高温胁迫明显,-5 ℃胁迫时,电子传递的量子产额ΦEo和捕获的激子将电子传递到电子传递链中QA-下游的其他电子受体的概率ψo分别比CK下降了18.9%(P< 0.01)和16.9%(P< 0.01),而40 ℃胁迫时两者仅比CK下降了9.4%和7.7%(表1)。
高低温胁迫使PSII单位面积反应中心数量RC/CS增加,-5 ℃和40 ℃胁迫时分别比CK增加了12.8%和6.9%。性能参数PIABS对某些胁迫比PSII最大量子产率Fv/Fm能更好地反映胁迫对光合机构的影响[4],-5 ℃和40 ℃胁迫时PIABS分别比CK下降了37.9%(P< 0.05)和18.0%,而Fv/Fm都只下降了1.3%(表1)。
2.3 温度对稳态叶绿素荧光参数的影响
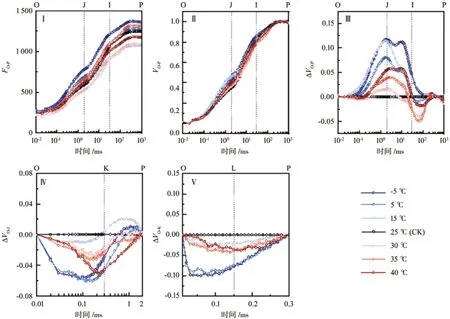
图1 不同温度处理对叶片叶绿素荧光动力学曲线的影响Fig.1 Effects of different temperature treatments on the chlorophyll fluorescence kinetics curve of leaves
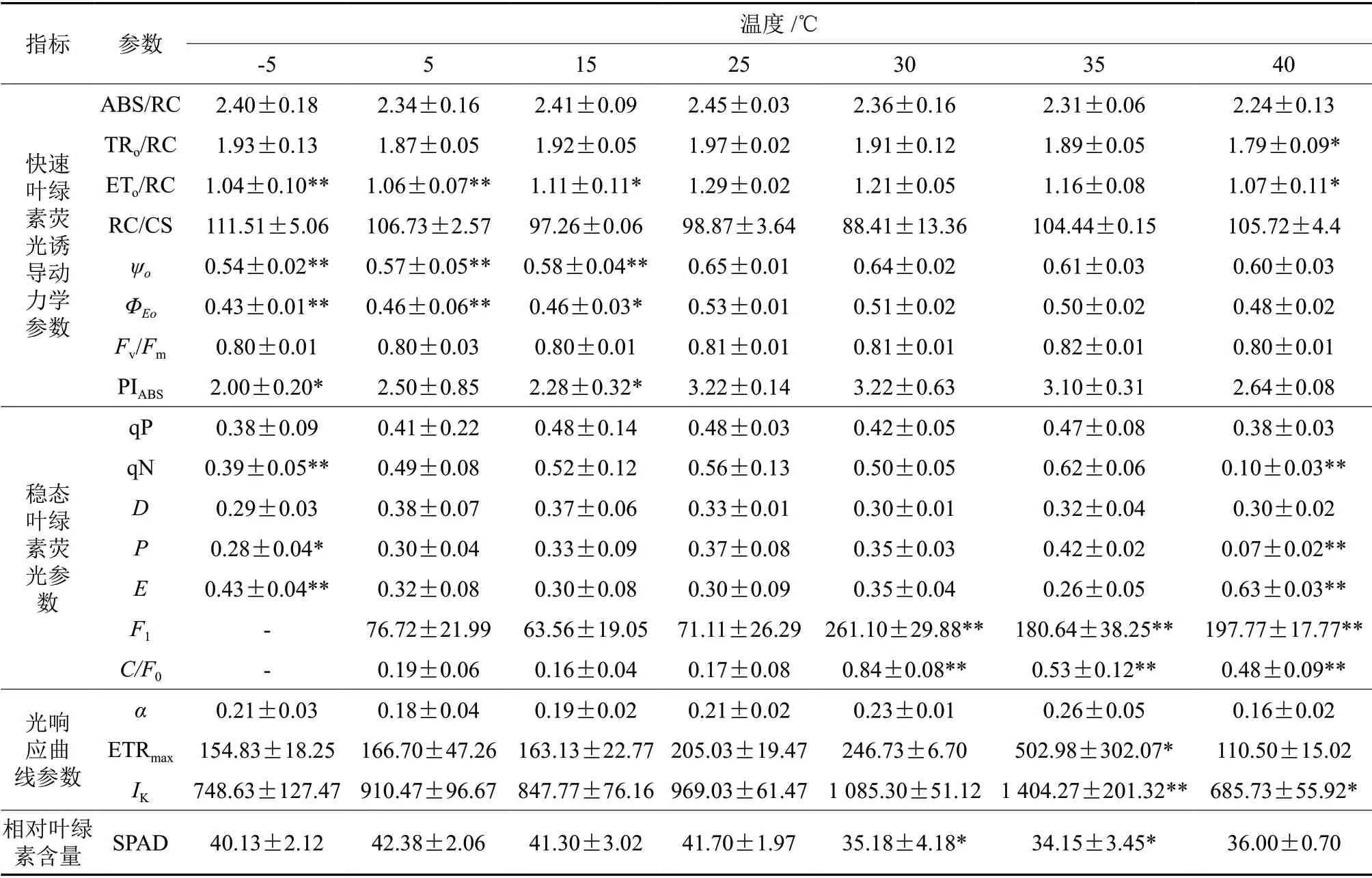
表1 不同温度处理下的叶绿素荧光参数†Table 1 Chlorophyll (Chl) a fluorescence variables in different temperature treatments
高低温胁迫对光化学猝灭系数qP无显著影响,显著抑制非光化学猝灭系数qN;-5 ℃和40 ℃胁迫时,qN与CK相比分别极显著下降了30.4%(P< 0.01)和82.1%(P< 0.01)。高低温胁迫下,PSII用于天线色素热耗散的能量比例D与CK相比无显著差异,但PSII非光化学反应耗散能量比例E显著上升,40 ℃和-5 ℃胁迫时分别比CK上升了110.0%(P< 0.01)和43.3%(P< 0.01)。高温胁迫比低温胁迫对PSII用于光化学反应的能量比例P的负面影响显著,40 ℃和-5 ℃时分别比CK下降了81.1%(P< 0.01)和24.3%(P< 0.05)。与CK相比,高温胁迫诱导PSI荧光强度F1和总荧光强度中PSI荧光强度的占比C/F0极显著上升,40 ℃时分别是CK的178.1%(P< 0.01)和182.4%(P< 0.01),低温胁迫对两项参数的影响不显著(表1)。
2.4 温度对光响应曲线的影响
各低温胁迫处理的RLC均低于CK,并且差异不明显;高温胁迫处理的RLC从高到低依次是35、30、40 ℃(图2),反映植物的光能利用能力参数α在35 ℃时达到最大值(0.26),在45 ℃时达到最小值(0.16)。ETRmax表示无光抑制时的最大电子传递速率,伴随温度的升高,在35 ℃时极显著升高到CK的245.3%(P< 0.05),但40 ℃时又回降到CK的53.9%;随着温度的降低,ETRmax也持续下降,但与CK无显著差异。Ik反映植物对强光照的耐受性,随着温度的上升,35 ℃时Ik极显著上升到CK的144.9%(P< 0.01),40 ℃时又快速回落到CK的70.8%(P< 0.05);低温胁迫导致Ik持续降低,但与CK无显著差异(表1)。
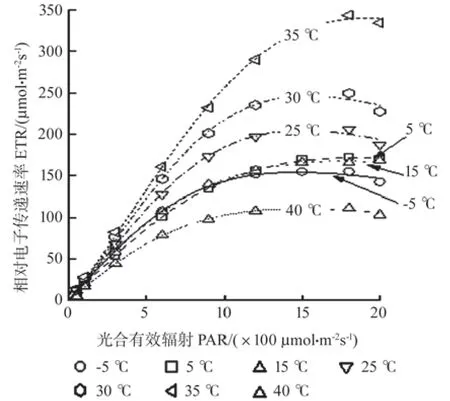
图2 不同温度处理对叶片光响应曲线的影响Fig.2 Effects of different temperatures on the RLC of leaves
2.5 温度对叶绿素含量的影响
温度胁迫下SPAD的变化可以反映光化学反应基础物质的变动情况。高温导致SPAD显著下降,30 ℃和35 ℃胁迫时分别比CK降低了15.6%(P< 0.05)和18.1%(P< 0.05),40 ℃胁迫时略有上升,但仍然比CK低13.7%;低温胁迫对SPAD无显著影响,15、5、-5 ℃胁迫时分别比CK降低了1.0%、1.6%、3.8%(表1)。
2.6 南极假山毛榉光合作用的最适温度预测
由表1可知,ETo /RC在多个温度处理下均与CK形成显著差异,说明温度变化极易通过干扰PSⅡ单位反应中心捕获的用于电子传递的能量,导致南极假山毛榉的光合速率发生改变。对不同温度处理所得的ETo/RC进行非线性拟合,得到5个潜在拟合函数(表2),其中Edgeworth-Cramer函数具有最高的AIC、adjR和SEE,所以确定为最优拟合函数,函数的极大值为23.83℃(图3)。
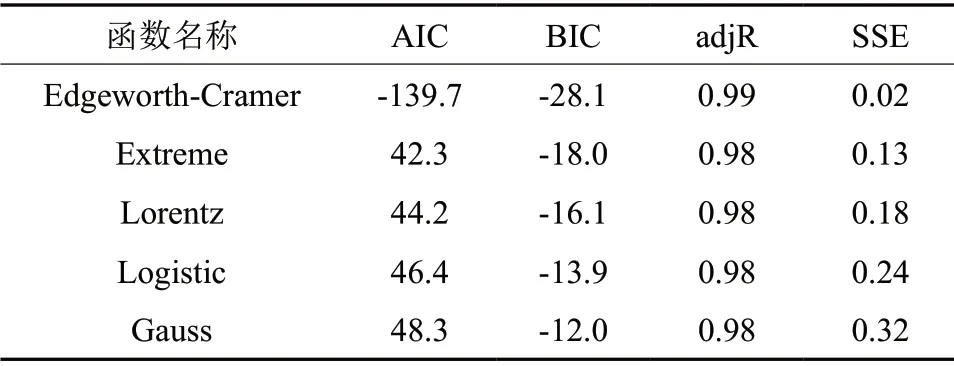
表2 潜在拟合函数与相应的AIC,BIC,adjR和SSETable 2 The latent function and the corresponding AIC,BIC,adjR and SSE

图3 不同温度处理下ETo/RC的非线性拟合Fig.3 Non-linear fitting of ETo/RC under different temperatures
3 讨论与结论
通过快速叶绿素荧光诱导动力学曲线和JIP-的测定,可以发现高低温胁迫主要引起南极假山毛榉叶片PSII反应复合体的电子传递受阻以及PSII放氧复合体受损,其主要表现是胁迫下ETo/RC持续显著下降,这表明光合作用Z链中脱镁叶绿素(Pheo)传递电子至醌(QA、QB)的路径不畅,以及叶绿素荧光动力学曲线中出现K点和L点,这预示了放氧复合体受损[29-30]和PSII各单位之间的能量流通不畅[31]。这一现象与已有研究结果相符[9],其机理可能源于PSII捕光复合体内的植物光合色素的光吸收能力较稳定,短期环境胁迫较难改变其性能[32],但环境胁迫会降低光合速率,这必然导致叶片吸收的光能不能及时被固定而出现光能过剩[4]。在本研究中,低温胁迫对SPAD无显著影响,高温胁迫虽然诱导SPAD显著下降,但降幅最高仅为CK的18.1%;同时,高低温胁迫下,反应光合效率的α、ETRmax和IK等参数均不同程度下降,而qN则持续上升,表明南极假山毛榉出现了光能过剩现象。
研究显示,胁迫下植物叶片通过增加荧光散射量以减轻光能过剩引起的光伤害[33]。为应对高低温胁迫导致的光能过剩,南极假山毛榉也采取了类似策略,即增加叶片的荧光散射强度,以耗散掉多余的吸收光能。例如快速叶绿素荧光诱导动力学曲线的J和I点荧光强度上升,40 ℃和-5 ℃胁迫时E分别比CK显著上升了110.0%(P< 0.01)和43.3%(P< 0.01),以及高温胁迫下F1和C/F0极显著上升。在高低温胁迫下,南极假山毛榉PSII的RC/CS在-5℃和40℃胁迫时分别比CK增加了12.8%和6.9%,在以往类似的温度胁迫研究中,也观察到RC/CS有上升[34]和下降2种变化趋势[35-36],推测这是南极假山毛榉抵御胁迫的另一种方式,即通过增加PSII单位面积光合机构含有的反应中心数量,以加大光能吸收量。但因为已有的反应中心数量有限,合成新的反应中心需要时间与原料,导致第一种策略的效果不理想,所以胁迫下RC/CS与CK相比没有达到显著差异。已有研究发现,为维持光合能量传递的稳定性,PSII和PSI之间可以通过能量溢出机制,将PSII多余的能量传递到PSI,帮助其进行能量耗散[33]。本研究也发现,随着高温胁迫程度的加深,PSI的荧光散射强度也随之增强,这可能是减缓PSII光伤害的第3项措施。
低温胁迫对南极假山毛榉RLC的α、ETRmax和IK没有显著影响,说明其实际和潜在光合能力对低温有较好的耐受性,与文献报道南极假山毛榉耐寒性强的结论相符[24]。本研究得出南极假山毛榉光合作用的最适温度是23.83 ℃,这一温度低于通常C3植物的最适温度(25 ℃左右)[3],预示南极假山毛榉耐寒性强的重要生理基础可能是其光合系统能在较低温度下运行。关于南极假山毛榉能耐受的最高环境温度还未见报道,但有研究发现,接近30 ℃的环境温度对南极假山毛榉的生长没有显著影响[19],本研究也发现高温35 ℃胁迫时才导致多项参数显著下降。研究中还发现,高温胁迫对南极假山毛榉光合系统的损伤明显大于低温胁迫,在高温40 ℃胁迫下,多项参数数值低于-5 ℃的数值,其机理还需进一步探究。
