鄂西南川陕鹅耳枥种群结构与动态
2019-09-05艾训儒吴漫玲洪建峰
朱 强 ,艾训儒 ,姚 兰 ,朱 江 ,吴漫玲 ,黄 小 ,王 进 ,洪建峰
(1.湖北民族大学 林学园艺学院,湖北 恩施 445000;2.湖北星斗山国家级自然保护区管理局,湖北 恩施 445000)
植物种群结构和动态是植物个体生存力与环境相互作用的结果,研究森林优势种群的结构与动态,对阐明森林生态系统的形成与维持、群落的稳定性与演替规律,种群的生态特征和更新具有极为重要的意义[1]。种群结构主要包括年龄结构、径级结构、冠幅结构和高度结构等,可以反映种群内不同个体数量的分布状况[2],年龄结构分析、静态生命表、存活曲线和生存分析等是进行种群结构和动态分析的重要工具与手段[2-4]。
川陕鹅耳枥Carpinus fargesiana,落叶乔木,喜光,属桦木科Betulaceae鹅耳枥属Carpinus,是鄂西南海拔1 000 m以上山地常绿落叶阔叶混交林中较典型的森林群落优势树种。目前,对鄂西南常绿落叶阔叶混交林群落方面的研究主要集中在植物区系分析、群落学一般特征、优势树种生态位及植被生态恢复等方面[5-7];近几年,又开始基于大样地对鄂西南山地常绿落叶阔叶混交林群落结构、动态及物种多样性特征等方面的研究[8-10],然而尚未见对鄂西南不同地段优势种群结构与动态的研究报道。本研究基于鄂西南4个样地典型地段植被调查数据,分析种群年龄结构特征,用线性回归模型拟合种群胸径和树高相关性,编制静态生命表,绘制存活率曲线、死亡率曲线、消失率曲线,同时用生存分析函数量化种群动态,旨在为川陕鹅耳枥种质资源的有效保护和科学管理提供理论依据,同时对鄂西南常绿落叶阔叶混交林群落生态系统结构和功能的维持有着重要意义。
1 研究区概况与研究方法
1.1 研究区概况
本研究选取的4个川陕鹅耳枥种群分别位于湖北七姊妹山、湖北星斗山、湖北木林子等3个国家级自然保护区和湖北利川金子山国有林场的常绿落叶阔叶混交林典型区域,其分属恩施土家族苗族自治州的宣恩县、利川市-恩施市-咸丰县交界区域、鹤峰县及利川市,研究区域属亚热带山地季风湿润气候区,伴生乔木树种主要为多脉青冈Cyclobalanopsis multinervis、亮叶桦Betula luminifera、锥栗Castanea henryi、白栎Quercus fabri、大叶杨Populus lasiocarpa等,以山茶科、壳斗科、蔷薇科等树种为主。研究区概况如表1所示。

表1 研究区概况Table1 Survey of study area
1.2 研究方法
1.2.1 样地调查
2014年7月至2017年10月,分别在星斗山自然保护区、木林子自然保护区、七姊妹山自然保护区和金子山国有林场选择生境相似的亚热带常绿落叶阔叶混交林中的川陕鹅耳枥为优势树种的区域布设样地。按照CTFS(Centre for tropical forest science)样地监测与技术规范标准设置20 m×20 m的固定样地共40个,星斗山、木林子、七姊妹山自然保护区及金子山国有林场各10个。对胸径(DBH)在1.0 cm以上的所有木本植物进行每木检尺,分别对种名、胸径、树高、萌生等指标进行调查,并测定所有个体在样地内相对坐标和其它立地因子。
1.2.2 种群结构划分
采用上限排外法按步长4.0 cm由低到高将川陕鹅耳枥种群划分8个径级:Ⅰ级,1~5 cm(木本植物起测径阶为1 cm);Ⅱ级,5~10 cm;Ⅲ级,10~15 cm;Ⅳ级,15~20 cm;Ⅴ级,20~25 cm;Ⅵ级,25~30 cm;Ⅶ级,30~35 cm;Ⅷ级,35 cm以上。根据川陕鹅耳枥树高特征和姚兰等对木林子树种高度级划分[9],由低到高将种群划分为6个高度层:a层<5 m,b层5~10 m,c层10~15 m,d层15~20 m,e层20~25 m,f层25 m以上。分级统计4个地段川陕鹅耳枥种群数量,用4个地段各指标的平均值以代表鄂西南地区,分析川陕鹅耳枥种群径级和高度级结构,同时探讨川陕鹅耳枥种群树高与胸径相关生长关系。
1.2.3 种群静态生命表和存活曲线
生命表和存活曲线是研究种群结构与动态特征的重要工具[11-12]。本研究采用径级结构代替年龄结构的方法分析种群动态。根据4个地段各种群的径级结构编制静态生命表,绘制存活曲线、死亡率曲线和消失率曲线[2]。参考苏建荣等[13]的方法,采用匀滑技术对各龄级内的实际个体数进行匀滑修正,以避免生命表中死亡率可能出现负值的情况。
1.2.4 生存分析
参考李艳丽等的方法[14],采用生存分析的4个函数,即生存率函数S(i)、积累死亡率函数F(i)、死亡密度函数f(i)和危险率函数λ(i)[14],揭示川陕鹅耳枥种群生存规律。各函数计算公式:
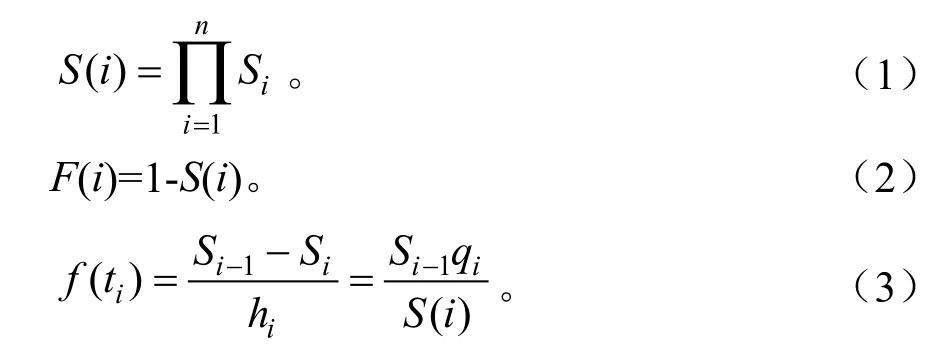

式(1)~式(4)中:Si为存活率;hi为龄级宽度;qi为死亡率。
1.2.5 数据处理与分析
选用R软件plot和boxplot函数对种群DBH与H关系进行分析,用abline函数和lowess函数添加最佳拟合的线性曲线和基于加权多项回归的平滑曲线,用数学模型进行拟合、检验并作图。其他作图在Origin2018软件中完成。
2 结果与分析
2.1 种群结构
2.1.1 种群径级结构
由川陕鹅耳枥种群径级结构(图1)可知,鄂西南川陕鹅耳枥种群属典型增长型种群,其种群数量峰值均出现在第Ⅰ级,木林子种群尤为明显。其中Ⅰ、Ⅱ级种群数量所占比例最高,平均占种群总量的73.21%;占比最大的是金子山种群,为86.24%;较少的是七姊妹山种群,为65.17%;木林子和星斗山种群在70.0%左右,相对而言,金子山川陕鹅耳枥种群其更新能力更强。胸径较大的Ⅵ~Ⅷ径级种群数量均较少,平均在1.66%左右;以金子山种群最少,仅占0.28%;七姊妹山种群相对较多,为2.37%,星斗山和木林子种群均在2.0%左右。第Ⅵ~Ⅷ径级个体在星斗山、木林子、金子山均有不同程度的个体数缺失,金子山种群缺失严重,未出现Ⅵ及以上的成龄树或老龄树。
2.1.2 种群树高结构

图1 各地段及鄂西南川陕鹅耳枥种群径级结构Fig.1 Diameter class structure of Carpinus fargesiana population in different sample plots
由鄂西南川陕鹅耳枥种群树高结构(图2)可知,不同地段高度级峰值均处在b层,居于该层的种群数量平均占种群总量的45.0%,金子山种群占比最高,为52.0%;木林子种群最低,为36.0%;七姊妹山和星斗山种群相当,在45%左右。其次是川陕鹅耳枥种群个体分布在a、c两层,居于a层的个体数占比平均为28.7%,c层平均为20.3%,a层和c层的个体数分布在不同地段存在一定差异,木林子和金子山种群,居于a层的个体数量相比c层要多,而星斗山和七姊妹山种群,a层的个体数量相比c层略少。鄂西南川陕鹅耳枥种群个体分布在20 m以上的e层和f层数量少,均不到1.0%,七姊妹山保护区为0.8%,星斗山保护区为0.3%,木林子保护区为0.2%,而金子山林场在该层次没有分布,可见三个自然保护区川陕鹅耳枥种群树高层次结构相比金子山林场更高。
2.1.3 胸径与树高的相关分析

图2 各地段及鄂西南川陕鹅耳枥种群高度结构Fig.2 Height class structure of Carpinus fargesiana population in different sample plots
川陕鹅耳枥种群树高(H)与胸径(DBH)的相关分析(图3)表明,鄂西南川陕鹅耳枥种群胸径(DBH)与树高(H)呈显著正相关(P<0.05),其线性回归方程为y=0.547x+3.991(R2=0.686),数学模型为y=3.061 9x0.5243(R2=0.756),拟合结果的可信度均较高。拟合模型表明,川陕鹅耳枥种群胸径(DBH)在增长初期,abline函数拟合的线性直线的斜率小于lowess函数拟合的平滑曲线;且种群DBH和H分别增长至3 cm、6 m及18 cm、15 m时,abline函数和lowess函数拟合的函数线交叉,说明种群前期树高生长显著,胸径生长趋缓;后期胸径生长相对较快,树高生长趋缓。

图3 川陕鹅耳枥种群胸径与树高的关系Fig.3 Relationship between DBH and height of Carpinus fargesiana population
2.2 种群静态生命表和存活曲线
2.2.1 川陕鹅耳枥静态生命表
鄂西南川陕鹅耳枥种群静态生命表(表2)表明,川陕鹅耳枥种群不同龄级的存活量差别较大,存活量随着龄级的增大而减少。死亡率和消失率最高均为Ⅴ龄级,其次为Ⅳ,Ⅱ龄级。随着龄级增加,Ⅰ~Ⅴ龄级川陕鹅耳枥的死亡率和消失率逐渐增加,这与川陕鹅耳枥生长前期的环境筛选相关,其后死亡率和消失率逐渐下降,表明川陕鹅耳枥大树的适应能力增强。种群期望寿命最高的是Ⅰ龄级,其次是Ⅲ、Ⅱ和Ⅵ龄级。Ⅰ~Ⅲ龄级的期望寿命高,说明该期间川陕鹅耳枥的生理活动相对旺盛,Ⅵ龄级的期望寿命高,可能由于川陕鹅耳枥进入中龄阶段后受到的环境筛选和竞争压力相对减弱。
2.2.2 川陕鹅耳枥各种群存活曲线
川陕鹅耳枥存活曲线(图4)表明,鄂西南川陕鹅耳枥种群存活曲线总体趋于Deevey-Ⅲ型,幼树死亡率较高,存活数量下降趋势较明显,随着树龄的增加死亡率较低而平缓。但各地段川陕鹅耳枥存活曲线在不同阶段呈现不同类型;图4表明,七姊妹山和金子山种群,在Ⅰ~Ⅲ龄级为Deevey-Ⅰ型,低龄级个体存活率相对较高,在Ⅲ~Ⅵ龄级大体呈Deevey-Ⅲ型,以后趋于平缓;木林子和星斗山种群,在Ⅰ~Ⅲ龄级为Deevey-Ⅲ型,低龄级个体存活率相对较低,Ⅲ~Ⅴ龄级为Deevey-Ⅰ型,以后趋于平缓。
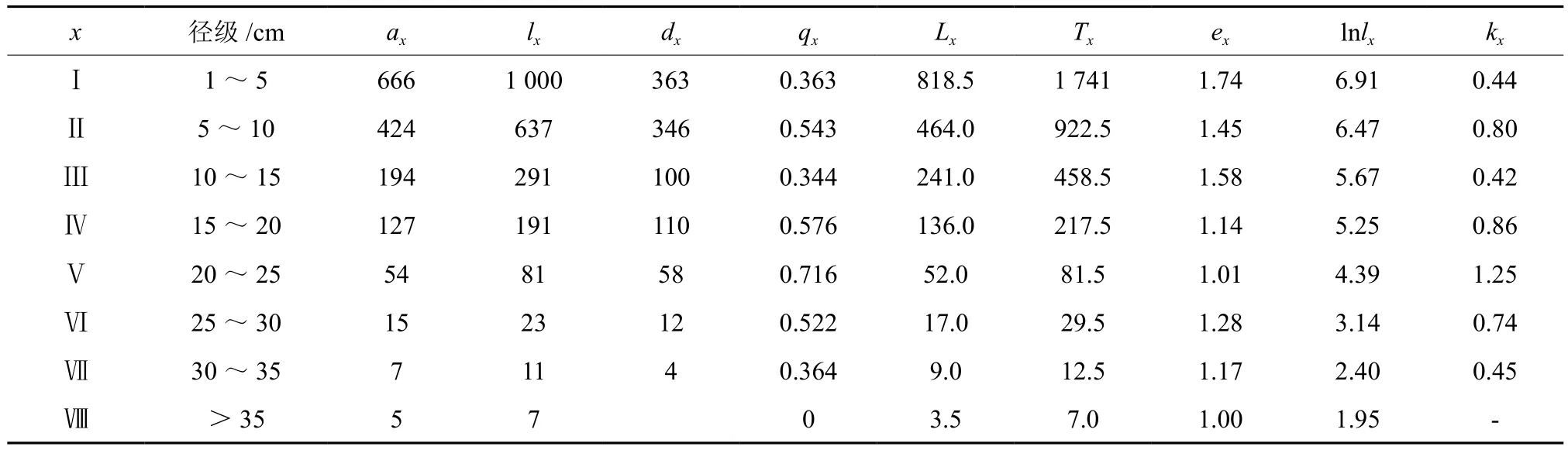
表2 川陕鹅耳枥种群静态生命表†Table2 Life table of Carpinus fargesian populations

图4 各地段及鄂西南川陕鹅耳枥种群的存活曲线Fig.4 Survival curves of Carpinus fargesian population in different sample plots
2.3 死亡率与消失率曲线
川陕鹅耳枥死亡率与消失率(图5)表明,鄂西南川陕鹅耳枥种群的死亡率和消失率曲线变化趋势保持一致。4个地段中,星斗山、木林子种群死亡率和消失率曲线均在Ⅶ龄级出现差异,种群死亡率在Ⅶ龄级达到最大值(1.00),金子山种群死亡率和消失率曲线在Ⅴ处出现差异,种群死亡率在Ⅴ龄级达到最大值(1.00),七姊妹山种群死亡率和消失率变化基本趋于一致;星斗山种群死亡率和消失率曲线在Ⅱ、Ⅳ达到高峰,消失率在Ⅳ龄级处达到最高峰(1.39);木林子种群死亡率和消失率曲线在Ⅰ、Ⅴ龄级为高峰,消失率在Ⅴ龄级处达到最高峰(1.29);金子山种群死亡率和消失率曲线在Ⅱ龄级达到峰值,Ⅱ-Ⅲ龄级消失率极高,Ⅱ为最高峰(1.36)。七姊妹山种群死亡率和消失率在Ⅴ、Ⅶ达到峰值,在Ⅴ龄级为最高峰(死亡率0.64;消失率1.02)。鄂西南川陕鹅耳枥种群死亡率和消失率曲线总体呈双峰型,在生长前期(Ⅱ级)和中期(Ⅴ级)达到高峰。
2.4 生存分析
川陕鹅耳枥生存分析(图6)表明,各地段川陕鹅耳枥种群随龄级增加呈现出生存率和积累死亡率曲线互补、死亡密度和危险率曲线基本一致的趋势。鄂西南及4个地段生存率曲线和累计死亡率曲线均在Ⅱ龄级处出现拐点,Ⅰ~Ⅱ龄级生存率锐减,累计死亡率骤增,在其它阶段种群生存率曲线和累计死亡率曲线的升、降相对较平缓;4个地段生存率与积累死亡率下降和增加的幅度均表现出前期高于后期的趋势,表明川陕鹅耳枥种群在幼龄时期死亡率较高。七姊妹山种群死亡密度先增后减,趋势较为平缓,在Ⅱ龄级达到峰值,该地段呈现前期锐减、中期稳定、后期衰退的趋势;星斗山种群生存率与积累死亡率在Ⅱ~Ⅲ龄级处出现短暂稳定期,死亡密度和危险率呈双峰型,在Ⅱ、Ⅳ处达到峰值,说明种群在前期短暂的高死亡率后又出现一次更高死亡率而后趋于稳定,具有前期锐减,中期波动,后期衰退的特征。木林子种群危险率函数先减后增,Ⅲ处为拐点;在Ⅴ处达到峰值,该地段具有前期略减、中期波动、后期衰退的特征。金子山种群死亡密度和危险率先增后减,表现较为剧烈在Ⅱ龄级处达到峰值,该地段种群在幼龄死亡率较高,种群呈现前期锐减,后期衰退的特征。鄂西南川陕鹅耳枥种群4个生存函数曲线具有前期锐减、中期稳定、后期衰退的特点。
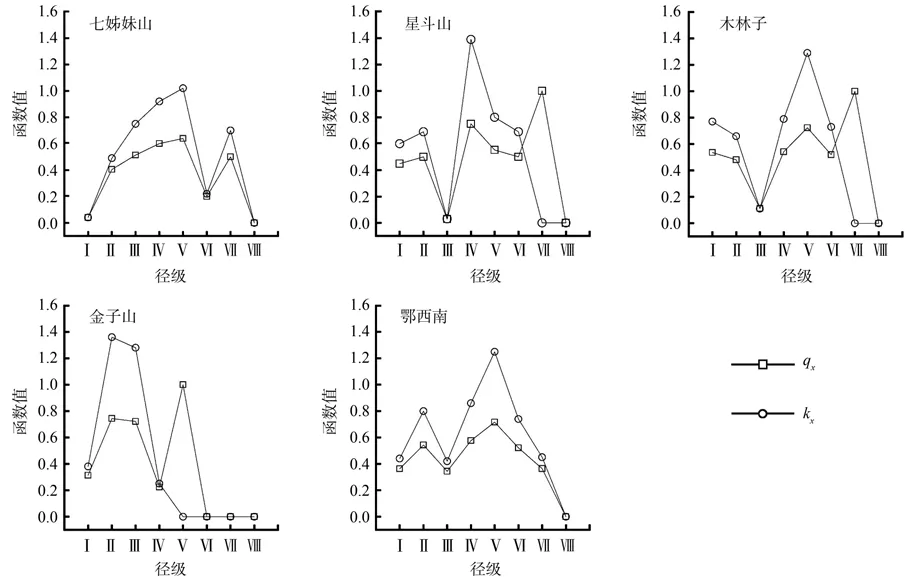
图5 各地段及鄂西南川陕鹅耳枥种群死亡率和消失率曲线Fig.5 Mortality and killing power curves of Carpinus fargesian population in different sample plots
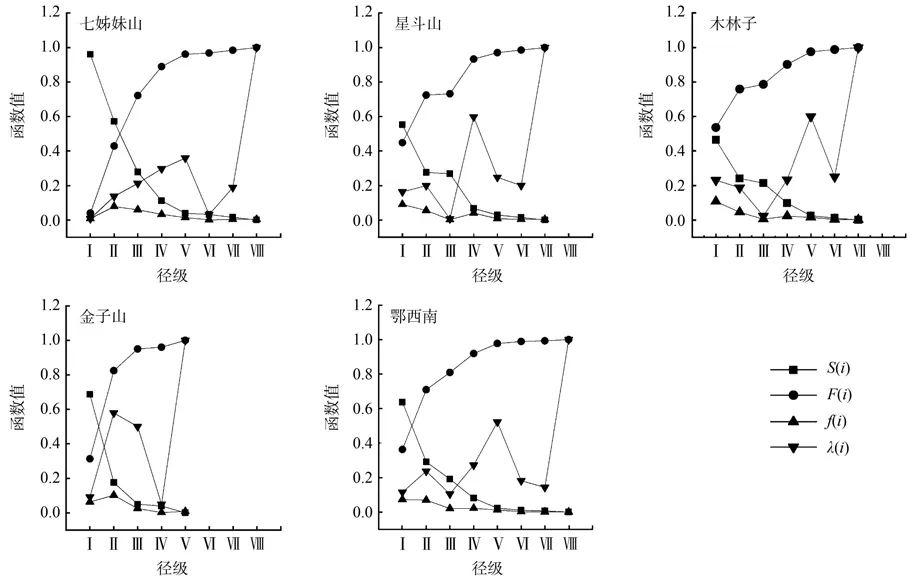
图6 各地段及鄂西南川陕鹅耳枥种群生存率、积累死亡率、死亡密度和危险率曲线Fig.6 Survival rate, accumulated mortality rate, mortality density rate, and hazard rate functional curves of Carpinus fargesian population in different sample plots
3 讨 论
3.1 种群结构
种群的径级结构可以代替年龄结构反映种群的生存状态,种群高度是描述种群垂直结构的重要指标之一,其个体高度分布状况能直观显示不同高度的种群个体在群落垂直结构中的地位[15-16]。种群结构分析表明,鄂西南4个地段的川陕鹅耳枥种群径级和高度级均在高龄级阶段有不同程度的缺失。鄂西南4个地段川陕鹅耳枥种群的年龄结构均属于倒“J”形,低龄级个体数量较多,幼苗植株储备量表现出一定的更新发展潜力。这与蒋迎红等[17]对极小种群847株海南吹风楠Horsfieldia hainanensis种群结构研究结果相似。金子山川陕鹅耳枥种群结构相比其他3个地段,小径级丰富、中大径级缺失的特征更为明显,说明了此地段种群较为年轻;星斗山与七姊妹山种群中,中高径级个体的占比较木林子和金子山高,种群较为成熟。鄂西南4个地段由于地理位置较近,气候相似等原因,4个地段川陕鹅耳枥年龄结构差异不大,树木径级结构表现为胸径普遍较小,仅有少数大径级个体,且川陕鹅耳枥种群所处群落样地密度较高,个体间竞争强度大,反映种群处于群落恢复的中期阶段[7,18]。
川陕鹅耳枥胸径与树高的数量关系可用于描述林分结构,分析生物多样性的固定监测样地的林分生长,对于准确估算森林蓄积量、碳储量有着重要意义[19-21]。川陕鹅耳枥胸径与树高呈显著正相关,且种群前期树高生长显著,胸径生长趋缓;后期胸径生长相对较快,树高生长趋缓;这是由于川陕鹅耳枥种群幼龄阶段处于林冠下层,为了能使自身生存,树高与胸径异速生长明显,树高生长较快;随着年龄的增长,川陕鹅耳枥渐渐处于林层的上部,资源利用性竞争减小,因此树高生长速度减慢。研究结果与林勇对金子山6个主要优势种群胸径与树高关系研究[12]及与程玉娜等对木荷种群胸径与树高生长模型及胡波等对刨花楠树高与胸径异速生长关系的研究相似[22-23]。
3.2 种群静态生命表与存活曲线
根据静态生命表可以分析种群过去生长状态、更新策略以及预测将来的种群动态;存活曲线是用各龄级存活个体数量相对时间来描述特定年龄的死亡率,表达种群动态的一种手段[24]。因此本研究在对鄂西南4个地段川陕鹅耳枥种群数量统计的基础上,编制静态生命表,绘制并分析存活曲线、死亡率曲线等参数来分析种群生存状态;鄂西南川陕鹅耳枥种群存活曲线趋于Deevey- Ⅲ曲线,但各地段川陕鹅耳枥存活曲线在不同阶段呈现不同类型,反映了不同地段种群对环境的适应及种间和种内竞争。其中,七姊妹山较其他3个地段低龄级个体存活率高;而中期存活曲线下降快,存活量低可能是因为七姊妹山种群中等径级的个体周围有较多的灯台树、四照花、多脉青冈等优势种,可能是由于种间竞争导致种群中期死亡率高;金子山低龄级个体存活量较星斗山、木林子低龄级存活量稍高,但下降速率极快,中龄级存活量极低,存活曲线前期趋于Deevey-Ⅱ型;而后期相对稳定,呈Deevey-Ⅲ型,这是由于中龄阶段后受到的环境筛选和竞争压力减弱所致,且幼年消失率明显较高,可能是由种群密度引起[12]。同时,星斗山、木林子存活曲线前期下降较快,到中龄级阶段趋缓,存活量较高,环境适应力较强,存活曲线趋于Deevey-Ⅲ型,且星斗山、木林子死亡率和消失率前期和中期较高,这是由于前期环境筛选和中期竞争所致。星斗山、木林子死亡率在Ⅶ龄级达到最大值金子山死亡率在Ⅴ龄级达到最大值,这与种群缺乏大龄植株相关,且金子山较为明显;这与张亚芳对黄土高原涿鹿和浑源地区杜松Juniperus rigida死亡率研究结果相似[2]。
从鄂西南川陕鹅耳枥种群生活史来看,中前期保持较高的死亡率和消失率,qx和Kx曲线整体呈双峰型,前期幼树死亡率较高,与其生物生态学特性及环境因素有关;原因可能是川陕鹅耳枥生长与分化速度较快,空间资源需求量大,从幼树到成树的生长过程中受到了相当程度的环境筛选和种内及种间竞争;同时,植物体的生长不仅受生物环境的影响, 还受到气候条件的影响,天气异常现象, 也会导致植株的死亡, 亦即种群的死亡和生存率是种群内在变化与极端环境条件相互作用的结果[25]。而消失率较低,可能是幼龄植株多,更新力强的原因。同时,中前期较高的生命期望值也反应出种群较强的发展潜力。
3.3 种群生存力
川陕鹅耳枥种群生存函数与种群存活曲线、死亡率曲线和消失率曲线分析结果一致,生存力分析进一步反映了种群的生存动态;4个地段的生存率曲线和累计死亡率曲线均在Ⅱ龄级处出现拐点,反映出这个阶段是川陕鹅耳枥种群生活史的脆弱环节,种群生存力低。此外,4个生存函数曲线反映出七姊妹山呈现前期锐减,中期稳定,后期衰退的趋势,种群的波动性较大。星斗山种群生存率与积累死亡率具有前期锐减,中期波动,后期衰退的特征;木林子地段具有前期略减、中期波动、后期衰退的特征。金子山具有前期锐减,后期衰退的特点。鄂西南川陕鹅耳枥种群总体具有前期锐减、中期稳定、后期衰退的特点,种群具有一定的幼苗储备。但幼苗生长前期可能因为种群密度大、生存空间较小且养分竞争强烈的原因,死亡率普遍较高。
4 结 论
川陕鹅耳枥种群低龄级个体数量较多,发展潜力良好,4个地段均表现出种群缺乏大龄植株;种群受群落年龄、环境筛选、种间及种内竞争等影响中前期死亡率较高,反映出川陕鹅耳枥种群结构年轻化及种群处在群落演替的初级阶段。保证幼苗存活率是种群恢复更新的关键,实地调查发现,川陕鹅耳枥萌蘖苗较为稀少,因此,在促进川陕鹅耳枥种群恢复更新过程中要适当地对种群进行砍伐催生,伐去部分上层林木,保存母树种源,改善林内光照条件,同时,幼苗附近常伴有茂密的低矮竹林,对幼苗的生长具有较大影响,因此,适度清理部分竹林,减小种间竞争,除去凋落物层及管理林地土壤能增加新幼苗定居的机会。
