桂林岩溶石山青冈群落植物功能性状的尺度变化与关联
2019-09-04李月娟李娇凤梁士楚
李月娟,李娇凤,常 斌,姜 勇,*,梁士楚
1 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室,桂林 541006 2 广西师范大学生命科学学院,桂林 541006
植物功能性状是指植物在长期适应环境的过程中,通过内部不同功能之间的协同进化,形成能够影响植物生存繁殖、死亡和最终适合度,并对生态系统结构及功能有一定影响的生物属性[1- 3]。植物功能性状的差异不仅表征了植物个体的生理过程及对环境的适应策略等的差异,而且与各种生态系统过程密切相关[4]。基于功能性状的群落生态学研究方法能够将物种的适应策略与群落构建和植物生态系统过程等有机结合起来,揭示植物对环境的响应和生态策略,对科学预测植被分布以及植被对全球变化的响应等具有重要的理论意义[5- 7]。
研究表明,在全球和区域尺度上,植物功能性状与环境具有显著的相关性,植物通过环境的筛选作用塑造其性状,而在局域尺度上,生物间的相互作用(极限相似性)对物种性状的塑造起关键作用,主要通过竞争使局域空间的植物性状趋异达到资源的合理分配与利用[8- 9]。那么, 如何根据性状变异推断环境筛选和生物竞争两种动力的相对重要性始终是生态学领域一个重大的挑战。最近 Ackerly和Cornwell提出了一个性状梯度分析(trait-gradient analysis)的新方法[10]。该方法采用定量的分析方法,能够将植物性状在群落内的变异(α组分)与在环境梯度上(β组分)的变化区别开来。如果存在明显的环境梯度,可以预期不同性状的β组分应该有很强的协变关系;反之,如果竞争排斥起主导作用,则物种在不同策略轴上分割生态位,可以预期不同性状的α组分仅有很弱的协变关系。显而易见,检验物种的α、β组分如何变异与关联对深入理解不同尺度上群落构建的机制具有重要的理论意义。
桂林岩溶区位于我国南方喀斯特地区东南部,分布面积高达2452 km2,是世界岩溶区演化最具典型代表的地区[11- 12]。由于岩溶地区岩石裸露率高,土层浅薄,土被不连续,地表水缺乏,导致生态系统极其脆弱,随着石漠化程度的不断加剧,岩溶地区面临的生态危机日益严重[13- 15]。然而,植被作为喀斯特退化生态系统恢复的主体,在维护生态系统的稳定和物种多样性的保护等生态服务功能方面具有重要作用[16]。以青冈(Cyclobalanopsisglauca)为优势种或共优种的群落是岩溶生态系统顶极群落的建群种,对于维持岩溶生态系统的结构和功能具有重要的作用。近年来,国内外学者对于青冈的研究主要集中于群落学特征[17- 18]、空间分布格局[19- 20]、种内竞争[21- 22]及幼苗生理特性[23- 24],对青冈的树干液流特征[25- 26]和光合速率[27- 28]等也多有研究,然而关于其功能性状的尺度变化与关联研究却鲜见报道。鉴于此,本研究以桂林岩溶石山青冈群落植物的叶面积(leaf area, LA)、比叶面积(specific leaf area,SLA)和木材密度(wood density,WD)为研究对象(叶面积(LA)主要反映了叶片与外界环境接触面积的大小,例如,在干旱且开阔的生境条件下,植物多采取“保守”性的策略来适应干旱的环境,叶面积一般较小,进而通过增大叶组织密度,减少叶片水分的丧失[29]。比叶面积(SLA)主要反映的是植物捕获光照资源的能力。一般来讲,比叶面积较高的物种生长速率较高,养分利用效率较低,比较适应土壤比较肥沃的生境,反映了植物采取的“开放性”策略[30]。木材密度(WD)作为影响植物碳分配、碳储量、水运输、生物量等功能过程的重要指标[8],一般采取与SLA相反的策略适应生境模式,采用性状梯度法等方法,分析该区植物功能性状的变异和关联,以期探讨以下两个科学问题:(1)植物功能性状在青冈群落间和群落内如何变化?(2)植物功能性状具有怎样的尺度依赖与关联性? 这些科学问题的合理解答,不仅能够揭示植物功能性状与生态系统结构的作用关系,还能帮助我们理解环境筛选与生物相互作用在群落构建中的地位和差异,从而为岩溶区的物种选择、植被恢复与重建提供科学的理论依据。
1 材料与方法
1.1 研究区概况
研究区位于桂林岩溶区处于演替晚期的青冈群落内,地处广西壮族自治区东北部(110°14′—110°42′E,24°43′—25°20′N)。该区域属于中亚热带湿润季风气候,雨量充沛,气候温和,并具有显著的岩溶地区特性(土层浅薄,岩石裸露率高,pH值为4—8之间,植被不连续),年平均气温为18—19℃,最冷1月份平均气温为8℃,最热8月份平均气温为28℃,全年无霜期达309 d,年降雨量为1856.7 mm,降雨量全年分配不均匀,春夏湿润多雨,年平均蒸发量为1458.4 mm[31]。该区域乔木层主要以青冈(Cyclobalanopsisglauca)为建群种,伴生种主要有扁片海桐(Pittosporumplanilobum);灌木层主要以一叶萩(Flueggeasuffruticosa)、粗糠柴(Mallotusphilippensis)、红背山麻杆(Alchorneatrewioides)、干花豆(Fordiacauliflora)、檵木(Loropetalumchinense)等为主;木质藤本以龙须藤(Bauhiniachampionii)为主。
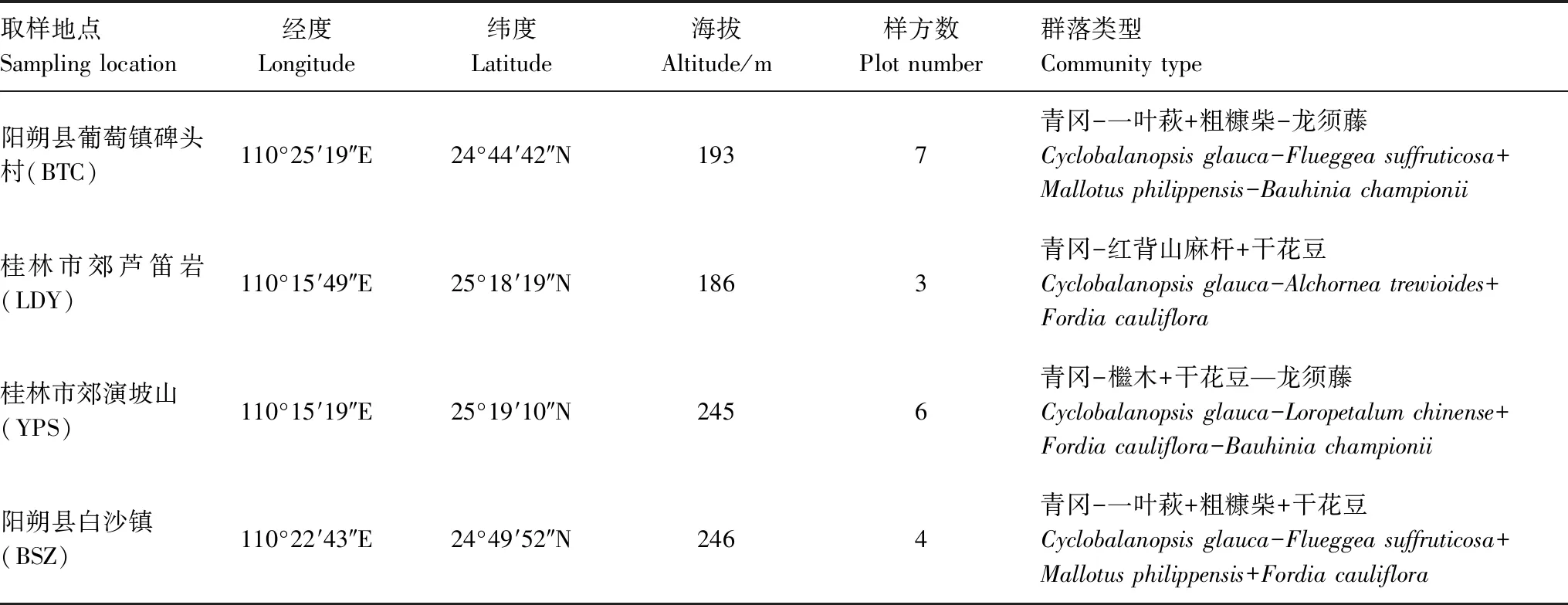
表1 样地基本概况
1.2 群落调查
在全面实地踏查的基础上,根据青冈群落的种类组成、结构、动态等特点。于2017年7—9月在桂林岩溶石山受人为干扰较轻且发育完好的青冈群落分布区,建立了20个20 m×20 m的典型样方。其中,芦笛岩3个、演坡山6个、碑头村7个、白沙镇4个。乔木样方大小为20 m × 20 m,在此基础上将每个样方划分成4个10 m×10 m的小样方进行灌木调查。对样方内所有乔木植株进行每木调查,测量并记录种名、高度、胸径、冠幅和盖度;对灌木层内所有灌木或木质藤本个体记录种名、株数、基径、高度及盖度。所有样方拥有4123株植物个体,隶属74个种。其中乔木、灌木和藤本的个体数分别为2310、1234和579。
1.3 功能性状测定
测定每个样方内胸径≥1 cm的所有木本植物个体的功能性状(包括乔、灌木和木质藤本)。对调查过程中出现的物种,每株植物个体选取3片完全展开的健康向阳叶片以及3根10 cm左右的非当年生枝条放入密封袋内,置于阴暗环境下,当天野外实验结束后带回实验室进行功能性状测定。对于叶性状,首先利用叶面积仪(Yaxin- 1241,Yaxin,China)测量每片叶片的叶面积(cm2),然后将叶片样品编号置于80℃烘箱中烘48 h至恒重,称量并记录叶干重。根据公式:比叶面积(cm2/g)=叶面积/叶干重获取比叶面积数据。枝条密度测定:取回的枝条削去表面的树皮后,使用电子天平(BSM- 220.4,Zhuojing,China)称量并记录鲜重,利用量筒排水法测其体积,然后将其编号并置于80℃烘箱中烘干至恒重后,称量并记录干重,根据枝条干重与体积的比值计算枝条密度(g/cm3)。根据枝条密度和木材密度是过原点的线性回归模型统计得出二者具有显著的线性关系 (P< 0.001)[32],进一步获取本研究中的木材密度数据。
1.4 性状梯度分析法
性状梯度分析法是由Ackerly和Cornwell[10]提出的,是将植物群落按照其群落间物种性状加权平均,然后基于群落加权性状均值大小进行排序,形成一维且具有潜在环境影响的性状梯度空间,进而将物种性状值累加降解为α和β两个组分。其中,β组分为物种在该性状梯度上所处的位置,表示物种在群落间沿着性状值的变化;而α组分是某一物种性状值与其所处群落平均性状值的差值,表示群落内部物种相对于共存种性状值的变化。性状梯度分析法主要计算公式如下:
(1)
(2)
(3)
αi=ti-βi
(4)
式中,pj为群落平均性状值,ti为物种平均性状值,aij为物种i在群落j中的多度或权重,tij为物种i在群落j中的性状值,n表示调查的群落总数,s表示调查出现的物种总数。本研究中n=20,s=74。
为使数据呈正态分布,首先对功能性状数据取自然对数。图1a为本研究中74个物种的平均比叶面积与群落平均比叶面积的散点图。图中灰色的点代表各群落中的物种,绿色实心方形代表青冈,黄色实心三角形代表一叶萩。x轴是群落平均比叶面积(pj),y轴为物种平均比叶面积(ti);矩形框起来的一列点代表在某一pj处群落j内所有共存物种。虚线y=x的斜率为一系列共存物种比叶面积均值与群落平均比叶面积值的比值。空心点代表物种i在群落梯度上的平均的β位置,空心点到虚线y=x的距离为αi,即群落中物种i比叶面积值与共存物种平均比叶面积值的差值。bi反映的是物种平均比叶面积值沿群落平均性状梯度在种内的变化大小。图1b为74个物种平均比叶面积与群落平均比叶面积的关系图,即群落中出现的所有物种的bi。突出显示的黑色实心点代表物种i所在各群落的群落平均比叶面积与群落j内物种个体比叶面积的交点。
图1a以青冈和一叶萩的比叶面积(SLA)为例来介绍其α和β两个组分的生态学意义。就比叶面积而言,参照上述的公式计算得到青冈的β组分为5.40,处于由群落平均比叶面积构建的性状梯度的下半部分,说明青冈常见于低SLA的群落中;αi值为-0.49,表示青冈在群落内相对其他物种有相对较低的SLA值,多处于群落演替后期,群落内部辐射较弱,湿度大,群落中常绿物种的比例较大,一般采取提高养分利用效率的保守策略。一叶萩的β组分为5.36,处于由群落平均比叶面积构建的性状梯度的上半部分,说明一叶萩多出现于高SLA的群落;αi值为0.81,表示一叶萩SLA略大于群落内其他植物,为了适应群落环境,采取提高生长速率的开放策略。
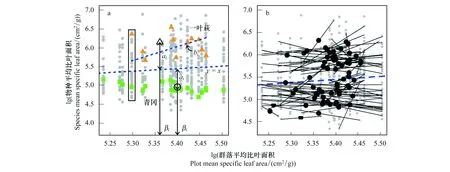
图1 物种比叶面积和群落平均比叶面积的散点图及其回归分析Fig.1 Scatterplot of species mean specific leaf area vs. plot mean specific leaf area and its regression analysis
2 结果与分析
2.1 植物功能性状在群落间和群落内的变化
如表2所示,叶面积α组分的取值范围为-1.87—1.87,β组分的取值范围为3.24—3.80,比叶面积α组分的取值范围为-0.74—1.07,β组分的取值范围为5.25—5.49,木材密度α组分的取值范围为-0.26—0.23,β组分的取值范围为0.60—0.67。综上所述,3个植物性状的α组分取值范围均大于β组分。叶面积的bi平均值为-0.08,比叶面积和木材密度的bi平均值分别为0.03,-0.06。
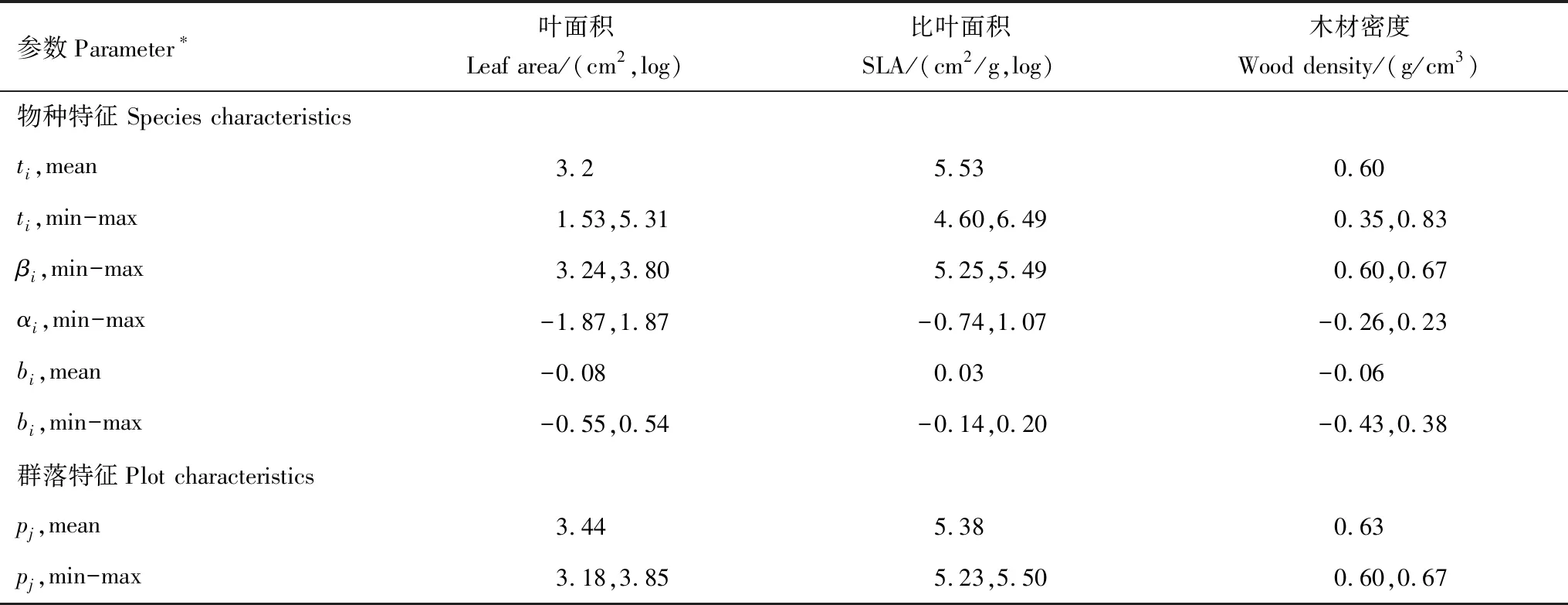
表2 植物功能性状参数的变化特征
SLA:specific leaf area;ti:物种平均性状值;βi:β组分值;αi:α组分值;bi:物种在种内的变化程度;Pj:样方平均性状值
2.2 样方平均性状值、物种功能性状及其α、β组分的相关性
如图2所示:物种水平,LA与SLA之间无显著的相关性,WD与LA以及SLA之间存在显著负相关;3个功能性状的β组分之间紧密相关(|r|> = 0.55),其中βLA与βSLA(r=0.55)呈极显著正相关关系,βSLA和βLA分别与βWD呈极显著负相关;αLA与αSLA之间无显著的相关,αSLA和αLA分别与αWD呈显著负相关或极显著负相关;群落尺度,LA与SLA具有显著的正相关性(r=0.54),SLA和LA分别与WD呈极显著负相关 (图2).
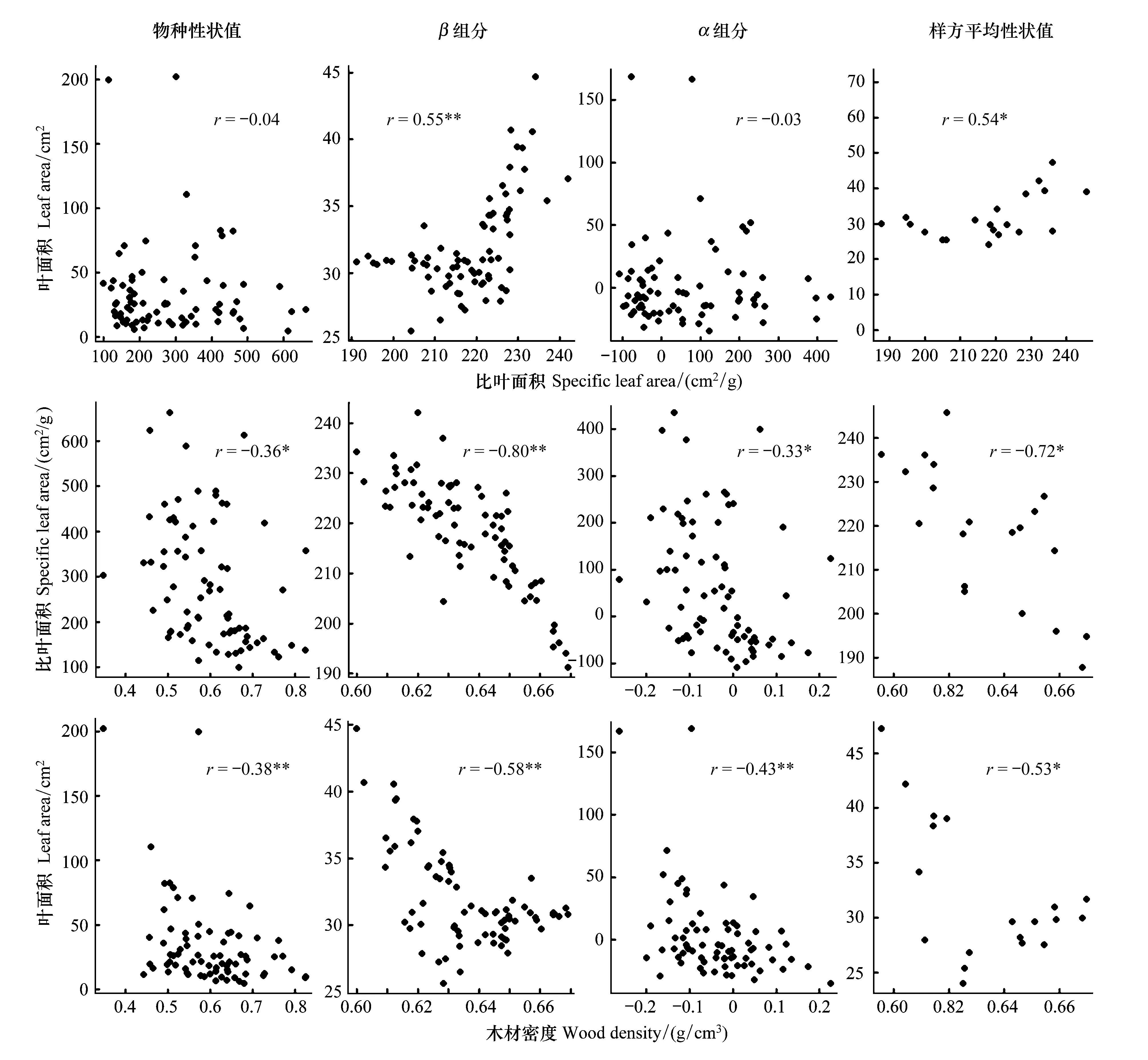
图2 性状相关性散点图 Fig.2 Correlation scatter diagram of plant function traits3种功能性状(叶面积leaf area, LA;比叶面积specific leaf area,SLA; 木材密度 wood density,WD)在物种平均性状值、β组分,α组分,样方平均性状值水平上的相关性散点图;图中数值代表两两性状间的pearson相关系数,*代表显著性。* P < 0.05; ** P < 0.01;无*代表的数字表示P>0.05
3 讨论
3.1 植物功能性状在群落内、群落间的变化
植物功能性状在群落内及群落间的变化规律,体现了环境筛选和生物竞争的共同作用[33]。一方面,环境筛选作用控制着物种性状值的分布范围,使群落内共存物种的性状趋同,另一方面,生物竞争会导致共存物种间产生生态位分化,使群落内共存物种性状趋异[34]。植物功能性状的α组分是某一物种与所处群落的平均性状值的差值,表征了群落构建中的生物竞争作用。β组分是物种在由群落平均性状值所构成的某一性状梯度上的位置,表征了群落构建中的环境筛选作用。本研究中,对α组分和β组分的分析结果发现,LA、SLA和WD这3个性状的α组分取值范围都大于β组分取值范围,暗示了α组分取值的离散程度均大于β组分。这表明在桂林岩溶石山青冈群落中物种相对于共生物种功能性状值的变化大于沿着群落平均性状值梯度的变化,即叶面积、比叶面积和木材密度在群落内的变化大于其在群落间的变化。这也暗示了在桂林岩溶石山青冈群落构建过程中,生物间的相互作用大于环境筛选的影响,这是因为群落内不同的物种在相似性限制(similarity limit effects)的作用下为了减少竞争进化出不同的功能性状组合来适应同一群落环境[35]。
植物功能性状不仅在物种间存在很大的差异,由于遗传多样性和表型可塑性的影响,同一物种不同个体的功能性状也会产生变异[36]。如,Messier等[37]研究巴拿马地峡沿降水梯度分布的3处原始低地热带森林功能性状跨多个嵌套生态尺度变异发现,功能性状的种内变异的总量大致等于种间变异量。杨建军等[38]在高山栲叶性状种内变异及其与环境因素关系的研究中表明高山栲叶性状在种内存在较大的变异,一个种群的性状平均值并不能代表整个物种的性状情况。本研究通过计算物种种内的3个功能性状和其群落平均性状构建梯度的比值,来反映各功能性状在种内的变化,即bi值。bi为正值,说明种内性状变化能够反映物种沿梯度的变化趋势,且bi<1,说明种内性状变化小于群落间因物种更替引起的性状变化。本研究中青冈群落的叶面积、比叶面积和木材密度的bi平均值分别为-0.08,0.03和-0.06,比叶面积的bi值在[0,1]区间内,而比叶面积和木材密度的bi值不在区间内,说明桂林岩溶石山青冈群落物种比叶面积在种内有变化,且种内比叶面积的变化小于群落水平比叶面积随环境梯度变化。这与丁曼等[5]对于黄土丘陵区植物功能性状的研究基本一致。
3.2 植物功能性状相关关系的尺度依赖
植物在漫长的进化过程中,植物功能性状之间彼此密切联系并通过协调进化来适应外界环境的变化(物理、生理等因素的综合作用),从而造成各功能性状之间在不同组织尺度往往会表现出一定的相关性[39]。Westoby等[40]的研究表明在全球以及区域尺度上,各植物功能性状之间表现出不同的相关关系,如基于全球和区域尺度的研究发现,叶面积与比叶面积均呈显著正相关[41-42],而同一群落内发现2者为不相关或负相关[43-44]。本研究发现,LA、SLA和WD这3种植物功能性状在群落和物种水平上的相关性存在明显差异。比如,LA、SLA这2个功能性状之间在物种尺度没有显著相关性(r=-0.04),这可能是由于α组分间不相关导致。但β组分和样方平均性状值均呈显著的正相关。唐青青[45]对亚热带常绿落叶阔叶混交林植物功能性状变异的研究发现:比叶面积与叶面积的样地平均性状值显著正相关。Ackerly[10]的研究也发现,2者在群落水平上正相关性较强,但物种水平上不相关或相关性较弱。而习新强等[4]对于黔中喀斯特山区植物功能性状的变异与关联研究中却表明两性状的β组分呈极强的负相关关系。综上所述,不同尺度的性状相关性差异可能是由于研究区群落类型、物种组成等的差异,导致不同尺度的植物功能性状间相关会表现出不同的地域依赖性。对比本研究中LA与SLA、SLA与WD、LA与WD的物种平均性状值、α组分、β组分和群落平均性状值之间的相关性,发现β组分的相关性最强,α组分的相关性则较弱,这表明LA与SLA、SLA与WD、LA与WD两两性状间的相关性在群落间的依赖性大于群落内共生物种的依赖性,植物功能性状对于环境筛选表现出较高的整体趋同适应性,即3个性状两两间β组分的相关性均较高。Westoby 等人在研究中证明了植物功能性状在自然选择或环境胁迫的作用下会表现出趋同适应(Convergent adaptation)[46]。在本研究中,青冈群落是岩溶石山特殊生境下的优势种群,比较能适应贫瘠,通常具有耐干旱生境的特殊能力,采取“保守型“策略。此外,青冈群落是岩溶石山演替晚期群落类型,研究发现样地内青冈物种多度占主导地位且常绿物种比例较大,说明在演替晚期的环境筛选下,植物在外貌形态、内部结构、生活史特点、生理特征及发育等诸多方面都表现出相似性,呈现出自然选择或环境胁迫植物功能性状趋同适应的现象。本研究发现WD与LA和SLA在各水平上均具有负相关性,这与Ackerly 和Cornwell对于加利福尼亚滨海木本植物群落的功能性状相关性的研究结果基本一致[10]。曹科等[47]对古田山木本植物功能性状的研究也表明叶面积与木质密度呈极显著的负相关性。李峰等[48]对于昆明西山半湿润常绿阔叶林木本植物功能性状的比较研究也发现比叶面积大、茎比密度小的植物具有较高的生长速率和生产力,而比叶面积小、茎比密度大的植物则具有较强的抵御能力。
4 结论
植物群落构建不仅受到环境因素的影响,也是生物相互作用的结果。研究植物功能性状的尺度变异和性状间的相关性能够更好的理解环境筛和生物竞争在群落构建中的作用,为揭示物种对生境的适应策略、物种共存和群落构建提供了新途径。本文采用性状梯度分析法,基于性状值降解得出:青冈群落的3个性状的α值范围均大于β值范围,表明物种相对于共生物种性状值的变化大于沿着群落平均性状梯度的变化。此外,物种比叶面积在种内呈现变化,且种内比叶面积的变化要小于群落水平比叶面积梯度变化。相关性分析表明3个性状的β组分的相关性均呈现最强,而α组分的相关性则较弱,即LA与SLA、SLA与WD、LA与WD两两性状间的相关性在群落间的依赖性大于群落内共生物种的依赖性,植物功能性状对于环境筛选表现出较高的整体趋同适应性。
