基于线粒体ATP6基因全序列分析中国绵羊痒螨(兔亚种)的遗传多样性
2019-08-29尹潇潇李燕方杨光友古小彬
尹潇潇,李燕方,古 江,廖 艳,谢 跃,杨光友,古小彬,*
(1.四川农业大学 动物医学院,四川 成都 611130; 2.眉山市畜牧站,四川 眉山 620000)
痒螨病是由痒螨科(Psoroptidae)、痒螨属(Psoroptes)的绵羊痒螨(P.ovis)寄生于绵羊、山羊、兔、水牛等家养动物及麋鹿等野生动物体表[1-2],引起的以结痂、脱毛、瘙痒为主要特征的慢性传染性皮肤病[3-5],可造成养殖业重大的经济损失,带来严重的动物福利问题,同时威胁野生动物的生存[6]。
中国养兔业发展迅速,家兔饲养量达7亿只以上,占全球饲养总量的40%以上[7]。痒螨病是危害我国养兔业的一种常见外寄生虫病,在我国23个省中流行,死亡率高达70%[8]。目前,对我国各地绵羊痒螨(兔亚种)种群的种群变异、种群结构域基因交流信息的了解较少,而这些信息对于研究绵羊痒螨(兔亚种)的动态历史及对养兔业中痒螨病的控制与消除是非常关键的。线粒体基因因其遗传稳定性好、进化速率快,可用于痒螨遗传分化、遗传结构和系统发生关系研究[9]。本研究基于线粒体ATP6基因全序列分析,旨在探讨我国绵羊痒螨(兔亚种)的遗传变异程度及遗传多样性。
1 材料与方法
1.1 绵羊痒螨(兔亚种)样品来源
从我国各地自然感染痒螨病的獭兔和肉兔耳部采集结痂,收集在洁净的平皿中,于37 ℃培养箱放置30 min,除去痂皮,收集虫体并鉴定[10],最终获得88个痒螨虫株(表1),-20 ℃保存备用。
1.2 引物设计
从绵羊痒螨(兔亚种)的线粒体基因组(GenBank No. KJ957822.1)获得ATP6基因全序列,利用Primer Premier 5设计引物:ATP6-F: 5′-AGAGTGTAAAAACTAACTGTTCCAA-3′;ATP6-R: 5′-TAATCACCCCCAAAGCAAAA-3′。引物由生工生物工程(上海)股份有限公司合成。
1.3 单个虫体DNA的提取
取出-20 ℃保存的88份痒螨样品,参考文献[11]的方法,采用OMEGA公司Mollusc/Arthropod基因组DNA抽提试剂盒进行单个虫体基因组DNA提取,-20 ℃保存备用。
1.4 PCR扩增及测序
PCR扩增体系(25 μL):Mixture 12.5 μL,上下游引物各1.0 μL,模板DNA 2.0 μL,双蒸水8.5 μL。PCR反应条件:94 ℃ 5 min;94 ℃ 45 s,55 ℃ 45s,72 ℃ 45 s,35个循环;72 ℃ 10 min。
表1 我国绵羊痒螨(兔亚种)不同种群样品采集情况表
Table 1 Sampling sites ofPsoroptesovisvar.cuniculipopulations in China

种群组 Population group省份 Province采样点 Sampling site样本数 Sample size编号 No.兔种类 Species西南地区 Southwest China四川 Sichuan德阳 Deyang3XN1~XN3獭兔四川 Sichuan眉山 Meishan1XN4肉兔四川 Sichuan资阳 Ziyang7 XN5~XN11肉兔四川 Sichuan雅安 Yaan4XN12~XN15肉兔云南Yunnan昆明 Kunming10XN16~XN25獭兔西北地区 Northwest China陕西Shaanxi宝鸡 Baoji19XB1~XB19獭兔华中地区 Central China湖北 Hubei荆州 Jingzhou8HZ1~HZ8獭兔河南 Henan开封 Kaifeng13HZ9~HZ21獭兔华北地区 North China河北 Hebei保定 Baoding9HB1~HB9獭兔华东地区 East China山东 Shandong济南 Jinan14H1~HD14肉兔
扩增结束后,将PCR产物进行1.0%琼脂糖凝胶电泳检测,将条带大小符合预期的胶块切下,经胶回收(天根普通琼脂糖凝胶DNA回收试剂盒,编号:P4418)、胶回收产物连pMD19-T载体、转化DH5α感受态细胞、蓝白斑筛选出阳性克隆,交生工生物工程(上海)股份有限公司进行正反向测序。
1.5 序列分析
1.5.1 遗传多样性分析
测序序列经DNAMAN 5.2软件拼接、比对,经DNASTAR 5.1软件剪切[12],通过MEGA5.0软件计算核酸分歧度(nucleotide divergence,Dxy)、平均核苷酸差异指数(average index of nucleotide difference,K)和遗传距离[13],运用DnaSP 5.1软件计算核苷酸多样性(nucleotide diversity,π)、单倍型多样性(haplotype diversity,Hd)和单倍型数(number of haplotypes,H)[14],运用Arlequin 3.1软件作分子方差分析(AMOVA)计算出种群间遗传差异指数(Fst),基因流(Nm)经下列公式计算:Nm=1/4(1/Fst-1)[15]。
1.5.2 遗传结构分析
利用Arlequin 3.5软件计算中性检验值(neutrality tests)结果(Tajima’sDtest,Fu’sFStest)[15],DnaSP 5.0软件分析错配分布曲线[14]。以欧洲屋尘螨(Dermatophagoidespteronyssinus,GenBank No. GQ469891.1)为外群,MEGA 5.1软件构建NJ系统发育树[13],为检验聚类树置信度bootstraps 1 000次,同时结合Network 5.0软件作单倍型网络图[16]。
2 结果与分析
2.1 绵羊痒螨(兔亚种)ATP6基因全序列的碱基组成及变异
88个绵羊痒螨(兔亚种)样品全部扩增成功,且测序获得ATP6基因全序列,88条ATP6序列长度均为672 bp,碱基平均组成由高到低分别是:T(45.53%)>A(22.92%)>G(17.82%)>C(13.73%),表现出明显的AT偏倚性(68.45%)。88条ATP6序列共有60个变异位点,其中简约信息位点28个,单变异位点32个。
2.2 种群遗传多样性
如表2所示,我国5大地区绵羊痒螨(兔亚种)种群的单倍型数(H)为41,单倍型多样性(Hd)范围为0.828 57(华中种群)~0.964 91(西北种群),种群整体单倍型多样性(Hd)为0.925 81,表明种群单倍型多样性丰富。5个种群中,核苷酸多样性(π)最低的是华东种群(0.005 82),西北种群最高(0.016 96),种群整体核苷酸多样性表现较高水平(0.014 94)。我国5大地区种群的平均核苷酸差异指数(K)范围为8.166 67(华北种群)~11.397 66(西北种群)(表2)。
2.3 种群分化和遗传结构
如表3所示,我国西北种群与西南种群遗传距离最大0.016 47,且差异极显著(P<0.001);华北种群与华中种群遗传距离最小0.012 28,且差异极显著(P<0.001);核酸分歧度范围为0.014 49(华东种群与华北种群)~0.017 63(西北种群与西南种群),整体表现与遗传距离的大小相一致。AMOVA分子变异分析显示,我国绵羊痒螨(兔亚种)种群变异来源分别为种群内部(97.92%)与种群间(2.08%),可见种群内分子遗传变异主要来自于种群内部。
表2 绵羊痒螨(兔亚种)种群遗传多样性参数
Table 2 Genetic diversity indexes ofP.ovisvar.cuniculipopulations in China
同列数据中“*”表示P<0.05,“**”表示P<0.01,“***”表示P<0.001。下表同。
In the same line, “*”indicatedP<0.05, “**” indicatedP<0.01, “***” indicatedP<0.001. The same as below.
表3 我国绵羊痒螨(兔亚种)各地理种群间的核酸分歧度(上三角)与遗传距离(下三角)
Table 3 Pairwise comparisons of the nucleotide divergence (above triangle) and genetic distance (below triangle) between geographical populations ofP.ovisvar.cuniculiin China
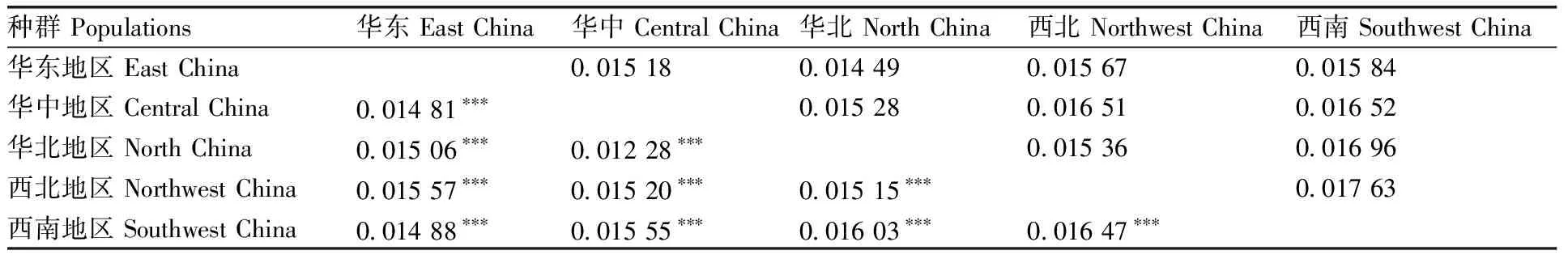
种群 Populations华东 East China华中 Central China华北 North China西北 Northwest China西南 Southwest China华东地区 East China0.015 180.014 490.015 670.015 84华中地区 Central China0.014 81∗∗∗0.015 280.016 510.016 52华北地区 North China0.015 06∗∗∗0.012 28∗∗∗0.015 360.016 96西北地区 Northwest China0.015 57∗∗∗0.015 20∗∗∗0.015 15∗∗∗0.017 63西南地区 Southwest China0.014 88∗∗∗0.015 55∗∗∗0.016 03∗∗∗0.016 47∗∗∗
如表4所示,我国5个地理种群间的Fst值为-0.044 52~0.106 12,以华北与西南种群间的遗传分化最高(Fst=0.106 12),西南、华北种群间基因交流程度最低(Nm=2.105 83)。我国绵羊痒螨(兔亚种)种群的总Fst值为0.020 81,总基因流11.763 5,表明我国种群内存在频繁的基因交流。
2.4 群体历史动态分析
中性检验结果见表2,所有种群的Tajima’sD值均为正数。华中、西北种群的Fu’sFs值为负值(-1.138 47和-2.714 84),且差异不显著(P>0.05),其余3个种群Fu’sFs值均为正值,说明5个种群在历史进化过程中均没有经历过种群扩张。种群整体的Tajima’sD值和Fu’sFs值均为正值(0.937 98和0.522 06),且差异都不显著(P>0.05)。核苷酸不配对差异分布曲线图具有多峰现象(图1),说明我国5个绵羊痒螨(兔亚种)种群在进化过程中稳定,无种群扩张事件。
2.5 单倍型网络图及进化树分析
从构建的NJ树可见(图2),外群欧洲屋尘螨(Dermatophagoidespteronyssinus,GenBank No. GQ469891.1)明显区分于绵羊痒螨(兔亚种)种群单独成一支,而绵羊痒螨(兔亚种)虫株被分为2个大支,且5个种群的单倍型在2个大支中都有分布。
表4 我国绵羊痒螨(兔亚种)各地理种群间基因流(上三角)与遗传分化度(下三角)
Table 4 Gene flow (above triangle) and genetic variation index values (below triangle) among 5 populations ofP.ovisvar.cuniculiin China

种群 Populations华东 East China华中 Central China华北 North China西北 Northwest China西南 Southwest China华东地区 East China3.850 242.342 59∞∞华中地区 Central China0.060 97∞∞3.622 30华北地区 North China0.096 43-0.044 5235.728 182.105 83西北地区 Northwest China-0.028 81-0.004 250.008 07∞西南地区 Southwest China-0.025 300.064 560.106 12∗-0.006 75

图1 基于线粒体ATP6基因的绵羊痒螨(兔亚种)种群核苷酸不配对差异分布曲线图Fig.1 The distribution curve of nucleotide mismatch in P.ovis var. cuniculi population based on mitochondrial ATP6 gene
单倍型拓扑结构表明,5个种群未出现基于地理位置的聚集,即绵羊痒螨(兔亚种)没有形成基于地理分布的遗传变化(图2)。由单倍型网络图可知(图3),大部分种群单倍型并未对应地理位置,所有单倍型以星状网络结构进行分布共享,大部分单倍型以5个单倍型(T1、T6、T7、T16、T24)为中心辐射分布,其中T6可能是最为原始的单倍型,并进化形成其它单倍型。整体单倍型分布聚类情况与NJ进化树类似,说明我国绵羊痒螨(兔亚种)种群间未形成基于兔品种、温度带或地域来源的种群结构。

CC,华中地区;NWC,西北地区;NC,华北地区;EC,华东地区;SWC,西南地区。CC, Central China; NWC, Northwest China; NC, North China; EC, East China; SWC, Southwest China.图2 基于线粒体ATP6基因单倍型构建的系统发育树Fig.2 Neighbor Joining (NJ) tree developed by haplotypes based on mitochondrial ATP6 gene

圆圈的大小与单倍型的分布频率成正比。The circle sizes were proportional to the distribution frequency of haplotype.图3 基于线粒体ATP6基因构建的单倍型网络图Fig.3 Haplotype network map based on mitochondrial ATP6 gene
3 讨论
寄生虫的生活史、宿主种类、生活习性、外环境、人类活动等因素均会影响其种群变异与种群结构[17-19]。绵羊痒螨作为广宿主寄生的全球性流行外寄生虫,其种群变异与种群结构研究非常有限。mtDNA以其严格的母系遗传和较快的进化速率[20],能够全面地反映种群间和种群内的遗传差异[21-22]。母系遗传可促使有利的突变迅速地形成、分离和表达,并迅速在该群体中得到富集,有利于探查自然选择在mtDNA留下的印记[23]。另外,ATP6、Cytb基因对自然选择的作用较为敏感[24],已见有学者将Cytb基因用于绵羊痒螨(兔亚种)种群的相关研究[25]。因此,本实验基于绵羊痒螨线粒体ATP6基因全序列探究我国绵羊痒螨(兔亚种)种群的遗传多样性,以期为痒螨病的合理防控提供基础资料。
截止2018年12月,GenBank中未见绵羊痒螨(兔亚种)的ATP6基因序列,故而本研究仅对我们扩增得到的88条ATP6序列进行分析。研究发现,绵羊痒螨(兔亚种)具有较高的单倍型多样性,且种群整体的遗传多样性水平较高。88个痒螨样本来自于我国7个省的10个采样点,而这10个采样点当地的平均气候、降雨量、光照等自然因素均有差异,由此可见,高单倍型多样性和高核苷酸多样性可能有利于各地的绵羊痒螨(兔亚种)发生进化,从而更好地适应当地环境而存活下来[26]。
分析种群遗传距离可知,种群间的遗传距离和地理距离未表现正相关关系。采用固定指数法度量种群间的遗传分化程度[27-28]时,我国绵羊痒螨(兔亚种)种群存在广泛的基因交流,且遗传分化较弱。本研究的各地种群间地理距离较大,且各种群间有山川、河流等天然屏障隔离,单靠痒螨虫体的移动来完成不同地理种群间的基因交流较难。绵羊痒螨个体较小,其离开宿主体后最长能存活18.8 d[29],痒螨离开宿主后,5 min内的最长运动距离不超过3 cm[30]。
本研究中,长距离痒螨种群间的频繁基因交流可能与以下因素有关:(1)我国各省间兔的频繁调运。以四川省为例,四川肉兔的消费占全国30%,每年需要从山东、陕西、云南等地调入兔只,尤以山东调入最多,这种频繁的调运导致四川与山东两地兔体寄生的痒螨群体间进行基因交流的机会增大,从而可能导致西南种群与华东种群的基因交流趋于无穷大。(2)我国养殖户的养兔模式和痒螨的生活史。我国养兔场多以兔自然交配的形式繁殖后代,兔的交配增加了不同痒螨群体的接触机会,从而增加群体间的基因交流机会;同时,痒螨属于终生寄生虫,其整个生活史均在宿主体完成,这种呈群居生活且同一群体内交配繁殖的特性,可增加群体间的基因交流,降低分化度,这点在线虫中亦得到证实[17]。(3)绵羊痒螨宿主范围广泛。已有研究证实,分离自兔的痒螨可感染牛,分离自绵羊的痒螨可以感染牛和家兔[31-32],这种宿主的广谱性可增加基因流动与交流。
我国痒螨整体种群的中性检测Tajima’sD和Fu’sFs检验结果均为不显著的正值,歧点分布为明显的双峰分布,表明中国地区的绵羊痒螨种群在长期的历史上一直处于较为稳定的状态,种群早期可能并未经历过群体扩张[33]。NJ进化树显示,我国绵羊痒螨各种群的单倍型并未基于地理来源聚类,而是混杂分布,未形成明显的系统地理格局,这点在单倍型网络图中亦得到进一步的证实。由此可见,我国绵羊痒螨(兔亚种)种群未形成基于兔品种、温度带或者地理来源的种群遗传结构,与赵习彬等[25]研究的结论一致,这可能与痒螨在我国养兔业中的高流行率及虫体高密度寄生有关[8]。痒螨在养兔业中广泛的流行率和虫体高密度的寄生方式导致种群遗传结构的缺乏,这点在微细毛圆线虫(Trichostrongylustenuis)的种群遗传研究中得到证实[34]。
