黔中喀斯特区不同光环境下光皮桦枝叶关系特征研究
2019-08-23袁丛军严令斌蔡国俊皮发剑
吴 华 袁丛军 严令斌 蔡国俊 皮发剑 吴 磊
(1. 开阳县林业局 贵州贵阳 550300;2.贵州省林业科学研究院 贵阳 550005;3. 贵州大学 贵阳 550025)
喀斯特区是我国典型的生态脆弱带之一,植被恢复对该地区森林生态系统的重建有着重要意义[1]。植物个体对环境的反应是通过其上具有重复性的基本组成单元—构件(Module)在数量、形状和空间排列上的变化实现的[2]。光是影响植物生长和分布的重要生态因子,对植物群体和个体的生长和发育具有重要影响。光照条件的变化能够直接影响植物的生理生态特性,从而改变植物的生存和生长状况,复杂多变的森林光环境对森林树种的生长发育和完成正常更新过程起着决定性的作用。植物在环境中的适应和演化过程中,植物构件中的各种功能性状是植物在其生长环境中做出的长期适应和响应[3]。枝、叶是植物进行光合作用的重要支撑器官和功能器官,其形状的表现对植物生长和发育极其重要。枝叶生长关系是植物在环境胁迫条件下提高空间资源利用能力的一种策略[4],对理解植物应对异质性环境的表型可塑性具有重要意义。目前,已有许多学者做了许多枝叶关系的研究工作。如杨瑞等人研究了喀斯特森林林分环境对光皮桦(Betulaluminifera)幼树的影响,结果表明不同的林分环境对光皮桦幼树枝和叶构件生物量影响较大,尤其是林中木叶构件受抑制强烈,生物量所占比例最小[5]。生物量分配是植物净碳获取的重要驱动因素[6],枝、叶关系对不同环境或干扰因素的响应存在差异。杨冬梅[7]等研究得出叶柄作为叶片生物量最大化的一个不利因素,影响了小枝内的生物量分配,并且叶片与其支撑结构之间的异速生长关系随叶片生活型的变化而变化。枝条大小和数量关系反映了植物适应环境胁迫的构型和生物量的分配策略,包括对气候环境和林分环境的适应。枝叶是林木树冠结构的主要骨架,枝条大小、长度、着枝角度决定了树冠的体积,为植物叶面充分合理利用光、温、水、气等提供空间平台,因此,开展不同林分环境光皮桦枝叶解析因子研究,通过分析不同冠层部位中的枝叶结构特征对描述树冠形状、预估林木生产力具有重要的现实意义。基于此,本文以不同林分环境下的光皮桦为对象,分析光皮桦枝-叶大小关系特征,试图解释枝-叶大小是否影响生物量分配以及叶面积支持效率等问题,从而有利于估算测定森林生物量、给森林经营调控提供理论支撑,本研究可为喀斯特石漠化地区光皮桦林的经营及天然次生林结构调控提供依据。
1 研究地概况
研究区位于贵州省贵阳市修文县龙场镇沙溪村修文河右岸,地理坐标为东经106°36′,北纬26°51′,海拔1100m~1500m。属温暖湿润的北亚热带季风气候,年平均温16.5℃,≥10℃活动积温 4097.40℃,年降雨量 1235mm,年日照时数1359.4h,土壤主要是以白云质灰岩为主发育的黄色石灰土和黑色石灰土,局部地段为黄壤,pH值4.0~7.4。现存植被主要为各类次生乔木林、乔灌从、灌木林、藤刺灌从等,植被盖度31.6%[8]。光皮桦为区域植被的重要组成树种,其现有林分为天然次生林和人促更新林。
2 研究方法及数据处理
2.1 解析木取样判别及数据获取
(1)解析木选择:参照杨瑞等人在研究区开展的不同林分环境下光皮桦的选择标准[9],解析木的因子分别为孤立木(DBH:9.48cm、H:9.91cm、HB2.91cm)、林缘木(DBH:9.15cm、H:11.42cm、HB:5.37cm)和林中木(DBH:9.01cm、H:11.67cm、HB:5.79cm),用围尺测树的胸高处(1.3m)胸径(精确到0.1cm)、标尺测树高及枝下高(精确到0.01m)、皮尺测量树冠东、南、西、北四个方向的冠幅(精确到0.01m)。
(2)解析木及枝叶数据采集:伐倒解析木前,记录解析木的立地条件、林分状况和主要树冠结构指标,并在树干上标记胸径的位置及南北方向。①采用主轴切割分层法测定树冠枝条解析因子。在“打枝”之前,详细记录各枝条的着枝高度,记为枝编号、枝高(Branch height 简称BH/m)、枝径(距离枝条基部约2cm处)、枝长、枝弦长、着枝角度、着枝深度(枝条距离树冠顶部的长度 DINC)等指标。② 标准枝生物量测定,分别测定各个标准枝的鲜重(FWb1),摘除叶片后再称得去叶枝重(FWb),叶鲜重为枝条鲜重减去叶枝鲜重(FW1=FWb1-FWb),按比例随机选取枝叶样品,带回实验室烘干至恒重后称重,计算标准枝生物量。
(3)枝、叶大小及数量指标确定:为合理量化样木枝条属性指标,采用一级枝横截面积(Primary branches of cross-sectional area,BCS/mm2)、一级枝条的枝体积(BV/mm3,采用枝横截面积BCS与枝长乘积的三分之一)、一级枝鲜重(BFW/g)表示枝大小。枝条数量采用一级枝上着生的二级枝条数量(BN/个)表示。采用一级枝条上面的叶片鲜重(LFW/g)表示叶片大小,一级枝上总叶面积大小(LFA,采用比鲜叶面积转换,光皮桦的比鲜叶面积取0.018 cm2/ g)表示叶数量。

表1 基本参数及其含义
2.2 数据处理
采用Excle 2013和SPSS 21.0进行数据整理与分析,进行One-Way ANOVA方差分析之前,将非正态分布的数据转换为自然对数形式,以满足正态分布要求和ANOVA 假设;采用Two-tailed的Pearson相关分析方法,数据结果以平均值或平均值±标准误表示。
所收集到的枝、叶属性包括枝鲜重、叶鲜重、小枝长度等。属性之间的关系用方程y=bxa来描述, 线性转换成log(y)=log(b)+alog(x), 其中x和y 分别代表所调查的不同枝条属性, b 代表属性关系的截距, a 为两者关系的斜率, 当a=1 时即为等速生长;当a≠1 时为异速生长。当数据两个变量都是正态连续变量,而且两者之间呈线性关系时,采用Two-tailed的Pearson相关分析方法。
3 结果分析
3.1 不同林分环境枝-叶结构分布特征
3.1.1 不同冠层部位的枝结构特征
不同光环境条件下的光皮桦平均枝长度随着冠层部位的往上而减小。在孤立木和林缘木中,II级枝随着冠层部位的往上而减少;而在林中木中,II枝数随着冠层部位的往上而增加,这是由于林中木受到的光照较差,因而需要通过多产生二级枝来增大与光的接触面积来转化生物量。平均枝夹角在林缘木和林中木中冠层中部为最大,分别是55.79°和53.05°。

表2 不同林分环境下各冠层部位的枝结构特征
注:针对解析木的树冠长均等分为三份,从树冠底部往树高顶端分别记为下部、中部、上部。
3.1.2 不同冠层部位的叶结构特征
在不同光环境条件下的光皮桦,其叶枝长、枝长度随着冠层部位的往上而变小。在孤立木和林缘木中,其一级枝个数随着冠层部位的往上而增加,其二级枝个数随着冠层部位的往上而减少;但在林中木中,其二级枝个数是随着冠层部位的往上而增加的,这是由于它需要更多光养分而做出的环境胁迫对策。在孤立木中,由于光养分充足,平均叶倾角是随着冠层部位的往上而增加的;在林缘木中,冠层中部光照较为良好,所以冠层中部的平均叶倾角最大(44.19°)。在林中木中,由于光养分的不足,所以冠层上部的叶倾角最大。
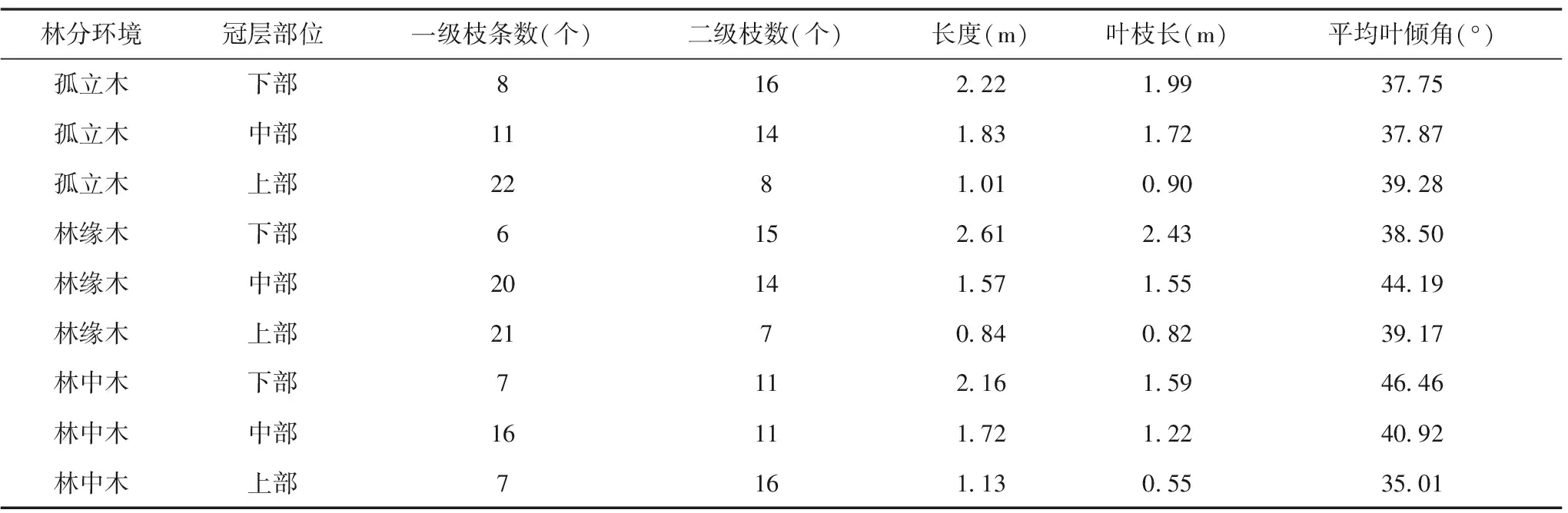
表3 不同林分环境下各冠层部位的叶结构特征
3.1.3 不同冠层部位的枝叶鲜重及生物量特征
树冠是林木叶片分布的主要部位,叶量大小、叶面积大小及其不同排列组合的分布形式直接决定林木枝条及全株林木的生产力[10]。在3种不同光条件下的光皮桦,孤立木的枝叶生物量在冠层中是冠层下部>中部>上部,林缘木的枝叶生物量在冠层中是冠层中部>下部>上部,林中木的枝叶生物量在冠层中是冠层下部>上部>中部;孤立木的枝基径、平均枝总鲜重、平均叶总鲜重、枝总鲜重、叶总鲜重在冠层部位中都是下部>中部>上部。在林缘木中,冠层中部枝总鲜重最多(1451.1g),所含枝叶生物量最多(2069.0g),其次是冠层下部(1500.5g)和冠层上部(812.1g);其枝基径、平均枝总鲜重、平均叶总鲜重、平均枝叶总鲜重都是冠层下部>冠层中部>冠层上部。在林中木中,冠层上部的叶生物量>冠层中部>冠层下部,这是由于林中木冠层上层部位的叶片能更好的得到阳光的照射的原因,所以平均叶总鲜重也是如此;但平均枝总鲜重、平均枝叶总鲜重是冠层下部>冠层上部>冠层中部。这是由于枝所含生物量比叶大得多的原因,而冠层下部因为要保持树体稳定,养分运输等原因往往是最重的。枝基径和枝叶生物量都是冠层下部>中部>上部。

表4 不同林分环境下各冠层部位的枝叶鲜重及生物量特征
注:不同冠层部位(下部、中部、上部)的叶片含水率分别为0.6255、0.6402、0.6566;不同冠层部位的枝含水率分别为0.5027、0.5235、0.5894。
3.2 不同林分环境枝-叶大小关系特征
3.2.1 不同光环境枝鲜重与叶鲜重的关系
林中木枝鲜重与叶鲜重(R2=0.587,b=0.041,斜率P=0.764);林缘木枝鲜重与叶鲜重(R2=0.731,b=0.293,斜率P=0.731);孤立木枝鲜重与叶鲜重(R2=0.701,b=0.326,斜率P=0.640)。

图1 不同光环境光皮桦枝鲜重与叶鲜重间的关系
3.2.2 不同光环境枝数量与叶鲜重的关系
林中木枝数量与叶鲜重(R2=0.450,b=0.331,斜率P=1.219);林缘木枝数量与叶鲜重(R2=0.477,b=0.648,斜率P=0.999);孤立木枝数量与叶鲜重(R2=0.440,b=0.469,斜率P=1.136)。

图2 不同光环境光皮桦枝数量与叶鲜重间的关系
3.3 枝叶面积相关方程拟合及评价
3.3.1 不同生长环境光皮桦枝叶面积与枝径关系
枝径对叶片鲜叶面积大小预估模型如图3所示。结果显示,孤立木的关系方程拟合效果依次为冠层上部(0.758)>冠层下部(0.757)>冠层中部(0.626)。林中木的关系方程拟合效果依次呈现冠层上部(0.708)>冠层中部(0.613)>冠层下部(0.425)。林缘木拟合方程决定系数大小依次为冠层上部(R2=0.834)>冠层下部(R2=0.799)>冠层中部(R2=0.805)。



图 3 光皮桦不同冠层部位的鲜叶面积大小与枝径间的关系
3.3.2不同生长环境光皮桦枝叶面积与枝长相关分析
采用枝长对叶片鲜叶面积大小预估拟合的方程如图4所示,从中可以看出,所有拟合的方程中不同林分环境中的差异较大,孤立木冠层上部的最优拟合模型为三次方程(R2=0.588)、冠层中部的最优拟合模型为幂函数拟合(R2=0.591)、冠层下部的最优拟合模型为三次方程(R2=0.628);林中木冠层上部的最优拟合模型为三次方程(R2=0.555)、冠层中部和下部的方程拟合优势度不好(最大决定系数没有超过0.250);林缘木冠层上部(决定系数R2=0.569)和冠层下部(决定系数R2=0.756)以三次方程拟合最优,而冠层中部的则以幂函数拟合相对较好。
4 结论与讨论
4.1 不同林分环境对冠层枝叶性状及生物量分配格局特征
林木树冠是树木进行光合作用和呼吸作用的重要场所,通过芽在树冠中的空间分布、数量动态、萌生格局等可塑性表达[11],不同程度的人为干扰或许是导致林木树冠冠形结构变化的重要原因[12],光皮桦枝、叶等结构性状特征综合表现塑造了其冠形结构特征,不同林分的光环境作用于其枝、叶等构件结构中,影响了光合产物在枝、叶等构件器官间的分配。研究结果显示,不同光环境下的光皮桦平均枝长度、枝基径、第一分支干径、分支干径、叶枝长等均随着冠层部位的往上而变小;不同林分环境光皮桦的树冠枝叶生物量分布格局不同,孤立木表现为冠层下部>中部>上部,林缘木为冠层中部>下部>上部,林中木为冠层下部>上部>中部。


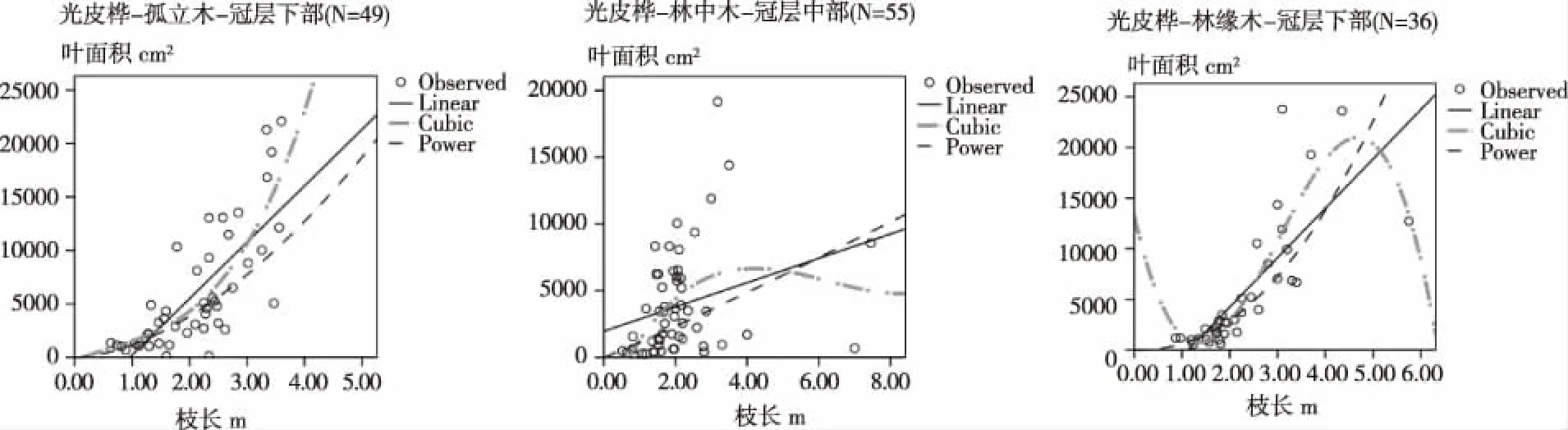
图 4 光皮桦不同冠层部位的鲜叶面积大小与枝长度间的关系
4.2 不同林分环境枝性状对枝叶面积模型拟合特征
枝与叶是林木冠层结构的主要骨架,枝条大小、长度、枝夹角决定了树冠的体积,为植物叶面充分合理利用光、温、水、气等提供空间平台。冠层结构是植物用以适应环境和提高整体光合效能所采取的一种生态对策,是植物在长期进化过程中形成的生理生态特征。光在冠层内部的分布是由进入冠层的太阳辐射经过植物体和地表面的多次投射、反射和吸收等一系列复杂的物理过程之后形成的[13]。不同林分环境下的光皮桦,枝基径等枝叶性状表现各异。孤立木的枝基径、枝总鲜重、叶总鲜重等枝叶性状在树冠中的大小表现为下部>中部>上部,林缘木为下部>中部>上部,林中木为上部>中部>下部;平均枝总鲜重、平均枝叶总鲜重是下部>上部>中部;枝基径和枝叶生物量都是下部>中部>上部。不同光环境中的枝叶各性状间相互关系的解释量不同。林中木、林缘木与孤立木的枝鲜重与叶鲜重函数关系分别为(R2=0.587,b=0.041,斜率P=0.764)、(R2=0.731,b=0.293,斜率P=0.731)和(R2=0.701,b=0.326,斜率P=0.640)。林中木、林缘木与孤立木的枝数量与叶鲜重函数关系分别为(R2=0.450,b=0.331,斜率P=1.219)、(R2=0.477,b=0.648,斜率P=0.999)和(R2=0.440,b=0.469,斜率P=1.136)。以枝长、枝基径为自变量,枝叶面积为因变量,建立了不同林分环境下各冠层的枝叶面积预估方程,不同光环境下光皮桦枝径预估枝叶面积最优方程依次为孤立木的冠层上部(R2=0.758)、林中木的冠层上部(R2=0.708)、林缘木冠层上部(R2=0.834);枝长预估枝叶面积最优方程依次为孤立木冠层下部(R2=0.628)、林中木冠层上部(R2=0.555)、林缘木冠层下部(决定系数R2=0.756)。研究结果对光皮桦林分抚育和结构调控提供理论依据。
