葡萄HAK基因家族的鉴定与表达分析
2019-07-17马丽娟何红红梁国平陈佰鸿
马丽娟,马 钰,何红红,梁国平,万 鹏,陈佰鸿,毛 娟
(甘肃农业大学 园艺学院,兰州 730070)
葡萄(VitisviniferaL.)作为世界四大水果之一,食用价值高,而且营养丰富。近年来,葡萄种植区域逐渐向南、北扩增种植[1]。西北地区由于具有适宜葡萄生长的生态环境,已经发展成为国内重要的葡萄与葡萄酒产区。然而,由于西北地区的土壤和天气状况,如:干旱、低温冻害频繁发生,土壤盐渍化程度高,使得西北地区贫瘠的土壤和脆弱的生态环境成为制约葡萄产业发展的瓶颈。
高亲和性钾离子转运体基因(high affinity K+transporter,HAK)是高亲和K+吸收转运体基因KUP/HAK/KT家族中的一员,HAK转运家族也称为KUP或KT转运家族[2],是植物钾离子转运载体4个家族(KUP/HAK/KT家族、Trk/HKT家族、KEA家族、CHX家族)中成员最多的一个家族,为H+/K+同向转运载体,主要介导植物中K+等元素的吸收转运和根部生长素的运输[3-5]。研究表明,该家族基因的表达受植物自身生长发育,调节因子和环境刺激等因素的调控,参与植物的生长发育和逆境响应机制[6]。其中,HAK基因的表达能使植物在低钾及盐胁迫条件下提高对K+的吸收能力,维持正常的K+浓度和生理功能,保持低Na+/K+比,从而提高植物的耐盐性[7]。在拟南芥中研究发现,盐胁迫下AtKUP2、AtKUP3、AtKUP6、AtKUP12的表达存在不同程度的上调[8]。HAK转运家族在旱胁迫方面也有重要的作用,在玉米中,存在多种假定的顺式作用元件,其中水分胁迫顺式元件ACGTATERD1在玉米的HAK基因家族的27个成员中都存在[9]。另外,还有元件ABRERATCAL、Ca2+-responsive element广泛存在于玉米HAK基因家族的23个成员中[10]。除此之外,还有其他非生物胁迫反应元件存在于大多数的成员中,ABRELATERD1、PRECONSCRHSP70A和SEBFCONSSTPR10A作用于脱水反应[11]。目前,对HAK基因家族的研究主要在拟南芥、水稻(Oryzasativa)和玉米(Zeamays)[10,12-14]中。
葡萄(VitisviniferaL.)是继拟南芥、水稻和杨树后完成全基因测序的第4种开花植物,其基因组大约是拟南芥基因组的3~4倍(约为500Mb)。目前,葡萄抗逆相关基因的研究已经比较广泛,但对于葡萄中的HAK基因家族成员抗逆性研究也未见报道,因此,本研究旨在利用生物信息学的方法进行葡萄HAK基因家族的分离与功能鉴定,进而为葡萄抗逆栽培调控与遗传改良等方面的研究提供理论依据。
1 材料与方法
1.1 试验材料
试验所用材料为葡萄‘红地球’(VitisviniferaL.‘Red Globe’)试管苗,保存于甘肃农业大学果树生理与生物技术实验室。将红地球葡萄试管苗茎段接种于GS固体培养基上,置于LED白光(WLED 424~724 nm,459 nm)的人工气候箱中培养。培养条件为28 ℃,220 μmol·m-2·s-1光照16 h;23 ℃暗培养8 h。
1.2 试管苗RNA提取
待葡萄试管苗生长至30 d后,选取长势一致的试管苗,分别用400 mmol·L-1NaCl、50 μmol·L-1ABA和质量分数为10% PEG处理 6 h,每个处理设置3组重复,以蒸馏水处理为对照。处理后取叶片于液氮中速冻,然后置于 -80 ℃冰箱中保存,并用CTAB法提取总RNA。
1.3 葡萄HAK基因家族成员的获得
下载玉米、水稻和拟南芥HAK基因的CDS序列,在葡萄基因组库的(http://www.genoscope.cns)Blast-Search中,检索筛选得到的葡萄HAK基因候选成员。运用DNAMAN软件,将候选的CDS序列进行比对分析,筛除相似度 >98%的基因,最后得到无重复区段的葡萄HAK基因家族成员的序列。
1.4 葡萄HAK蛋白理化性质的分析
运用ProtParam(http://expasy.org/tools/protparam.html)获得葡萄HAK基因家族成员的分子质量大小、理论等电点、编码蛋白氨基酸数目等基本理化性质;在作图软件中进行染色体定位,通过在线软件SOPMA(NPS@: SOPMA secondary structure prediction)进行二级结构的预测;通过PSORT(http://psort.ims.u-tokyo.ac.jp/ form.Html)进行亚细胞定位的预测。
1.5 葡萄HAK基因结构和顺式作用元件分析
通过GSDS (http://gsds.cbi.pku.edu.cn/)对葡萄HAK基因家族成员外显子的长度和结构进行预测[15];在NCBI中获得每个成员的启动子上游2 kb区域的序列,使用PLACE (http://www.dna.affrc.go.jp/PLACE/)在线软件[16]进行顺式作用元件分析;采用TMHMM在线软件[17](http://www.cbs.dtu.dk/services/TMHMM)进行跨膜结构域分析。
1.6 葡萄HAK基因多序列比对和系统发育分析
运用ClustalX 2.1[18]和 MEGE 5.0[19],使用邻近法绘制系统发育树,用DNAMAN对VvHAKs进行多序列比对。
1.7 葡萄HAK基因芯片表达谱分析
从NCBI提供的GEO数据库中下载葡萄RNASeq数据,登录号为GSE31594、GSE31662、GSE31675和GSE31677,数据分别是盐、干旱、低温、高温、ABA和水分胁迫下的表达数据。用葡萄LBD核酸序列为探针检索出序列相同的Affymetrix GeneChip 16 K基因ID,然后提取葡萄LBD基因在各种非生物胁迫下的RNASeq数据,通过Excel对数据进行log2转换,最后用HemI制作电子表达热图(heatmap)。
1.8 实时荧光定量(qRT-PCR)表达分析
应用Bio-Rad iCycler iQ实时定量PCR仪对VvHAKs进行逆境表达分析,使用SYBR GreenⅠ(TaKaRa)试剂盒,以葡萄管家基因UBI为内参基因[20]。扩增体系含1 μL cDNA,上下游引物各0.8 μL(表1),10.4 μL反应 SYBR MIX, 7 μL ddH2O,总体系20 μL。反应程序为:95 ℃下变性30 s,95 ℃变性5 s,60 ℃下退火30 s,40 个循环;95 ℃ 15 s,60 ℃ 30 s,95 ℃ 15 s。反应结束后分析荧光值变化曲线及熔解曲线。基因的相对表达量采用2-ΔΔCT计算[21]。
1.9 数据统计与分析
试验设置3次技术重复,用Excel 2013软件和SPSS version 22.0软件对试验数据进行分析。
2 结果与分析
2.1 葡萄HAK基因家族的理化性质和染色体定位
染色体定位分析表明(图1):VvHAKs的18个家族成员中,有17个成员分别分布于9条不同的染色体上,VvHAK09的位置未知。其中1号染色体上有5个成员,7号染色体上有4个成员,14号染色体上有2个成员,2、6、12、13、17和19号染色体上各有1个成员。除了7号染色体上的家族成员分布较为集中外,其余成员分布的较为分散,未表现出集中性。位置结构信息表明(表2),VvHAKs中CDS序列最短的是VvHAK07,为 1 845 bp,最长的是VvHAK04,为2 400 bp,大多数成员的CDS序列长度为2 000~2 500 bp;葡萄HAK基因家族编码氨基酸个数为614~800;葡萄HAK基因家族中,VvHAK02等电点最小,为7.35,VvHAK18等电点最大,为9.03,大多数成员的等电点主要集中在8~9之间,表明葡萄HAK基因家族的蛋白质呈弱碱性,外显子数目为7~12个。
2.2 葡萄HAK基因家族的结构分析
葡萄HAK基因家族的外显子个数为7~12,其中,VvHAK04、VvHAK08、VvHAK15、VvHAK13、VvHAK14、VvHAK16和VvHAK18均含有9个外显子;VvHAK02、VvHAK06、VvHAK10、VvHAK11和VvHAK12均含有10个外显子;VvHAK01、VvHAK05和VvHAK09均含有11个外显子;VvHAK03含有12个外显子;VvHAK07含有7个外显子;VvHAK17含有8个外显子。将葡萄HAK基因家族的18个成员进行聚类并结合外显子结构分析表明(图2),进化关系远近与外显子结构分布有关,聚类关系近,外显子数量相近,如:VvHAK05、VvHAK14、VvHAK15、VvHAK16和VvHAK18,可推测具有相似编码区结构的成员功能上有一定的相似性。

表1 葡萄HAK基因家族表达分析的实时荧光定量引物Table 1 The primers for qRT-PCR analysis of VvHAKs gene expression

1~18. VvHAK01-VvHAK18

表2 葡萄HAK基因的位置结构信息Table 2 Position structure informations of VvHAKs

图2 葡萄HAK基因家族的基因结构Fig.2 Gene structures of VvHAKs

图3 葡萄HAK多序列比对Fig.3 The multiple sequence alignment of VvHAKs
2.3 葡萄HAK基因家族的多序列比对与进化分析
VvHAK07、VvHAK13和VvHAK17这3个基因家族成员之间区段的保守性较低,其余的15个基因家族成员之间区段的保守性较高且在N端附近发现有典型的钾离子KT/KUP/HAK的保守区域GVVYGDLGTSPLY(图3)。系统进化树可将HAK基因家族(图4)分为7个亚族,其中VvHAKs成员分布为:第1亚族中有4个成员,包含VvHAK02、VvHAK06、VvHAK08和VvHAK09;第2亚族中5个成员,包含VvHAK01、VvHAK03、VvHAK04、VvHAK11和VvHAK13;第3亚族中有3个成员,分别为VvHAK01、VvHAK11和VvHAK13;第4亚族中只有VvHAK10;第5亚族中仅有VvHAK17;第6亚族中有2个成员,为VvHAK07和VvHAK12;第7亚族中有5个成员,包括VvHAK05、VvHAK14、VvHAK15、VvHAK16和VvHAK18。
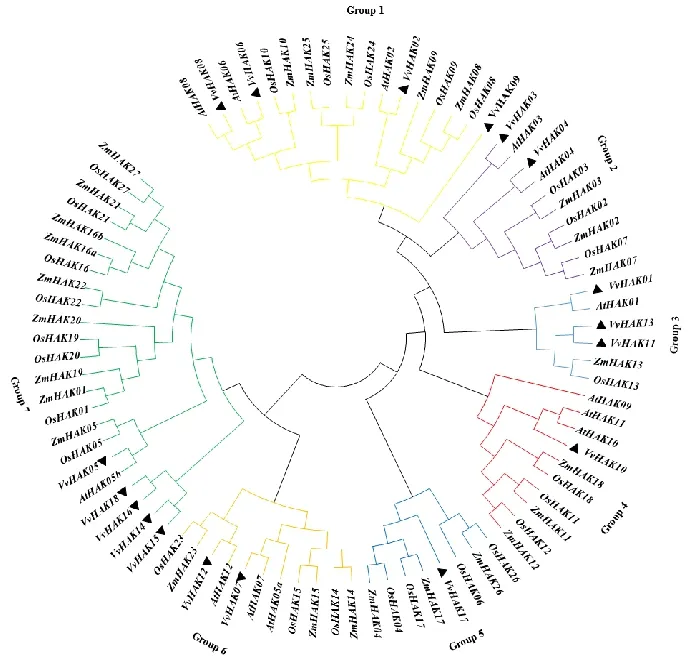
图4 拟南芥、玉米、水稻和葡萄HAK基因进化树Fig.4 Phylogenetic tree using the HAK amino acid sequence from grape,Arabidopsis,rice and maize
2.4 葡萄HAK基因家族编码蛋白Motif分析
Motif分析HAK基因家族的18个成员,得出有15个基序(图5)。Motif分析表明:VvHAKs中GroupⅠ的5个成员,VvHAK14、VvHAK15和VvHAK18有15个基序,VvHAK05不含Motif 15,VvHAK07不含Motif 11和Motif 12,整个GroupⅠ在 N端的保守性很高,结合聚类分析和外显子分析结果可推测,这5个成员在功能上具有相似性;GroupⅡ的9个成员,除VvHAK13外,其余8个成员在N端具有较高的保守性,具有较多相似的基序,推测这类亚组的成员执行功能相似;Group Ⅲ的VvHAK10和VvHAK12具有14个基序,没有Motif 15,VvHAK07不具有Motif 2、Motif 5、Motif 7和Motif 15,N端的保守性不高,与多序列比对结果一致。Group Ⅳ的2个成员,其中VvHAK07不具有Motif 7和Motif 15。由此可以得出:Motif 15为GroupⅠ中家族成员特有的基序,基序所表达的保守程度同多序列比对分析得出的结论一致。
2.5 葡萄HAK基因家族的二级结构分析和亚细胞定位
葡萄HAK基因家族编码蛋白质的二级结构主要以α-螺旋和不规则卷曲为主,成员跨膜区域数为9~14个(表3)。亚细胞定位结果表明(表4),葡萄HAK基因家族的18个成员在细胞质膜上均有表达,且表达量较高,这与其主要作为钾离子转运子参与K+吸收的功能是一致的;在细胞质和过氧化物酶体中几乎没有表达。但是VvHAK06在细胞质中的表达量高于在细胞质膜上的表达,推测此蛋白可能位于细胞质上;VvHAK08和VvHAK16这2个成员在细胞质和质膜上都有表达,推测这2个家族成员的蛋白在细胞质和质膜均有定位,剩余的15个VvHAKs成员则主要在质膜上表达。
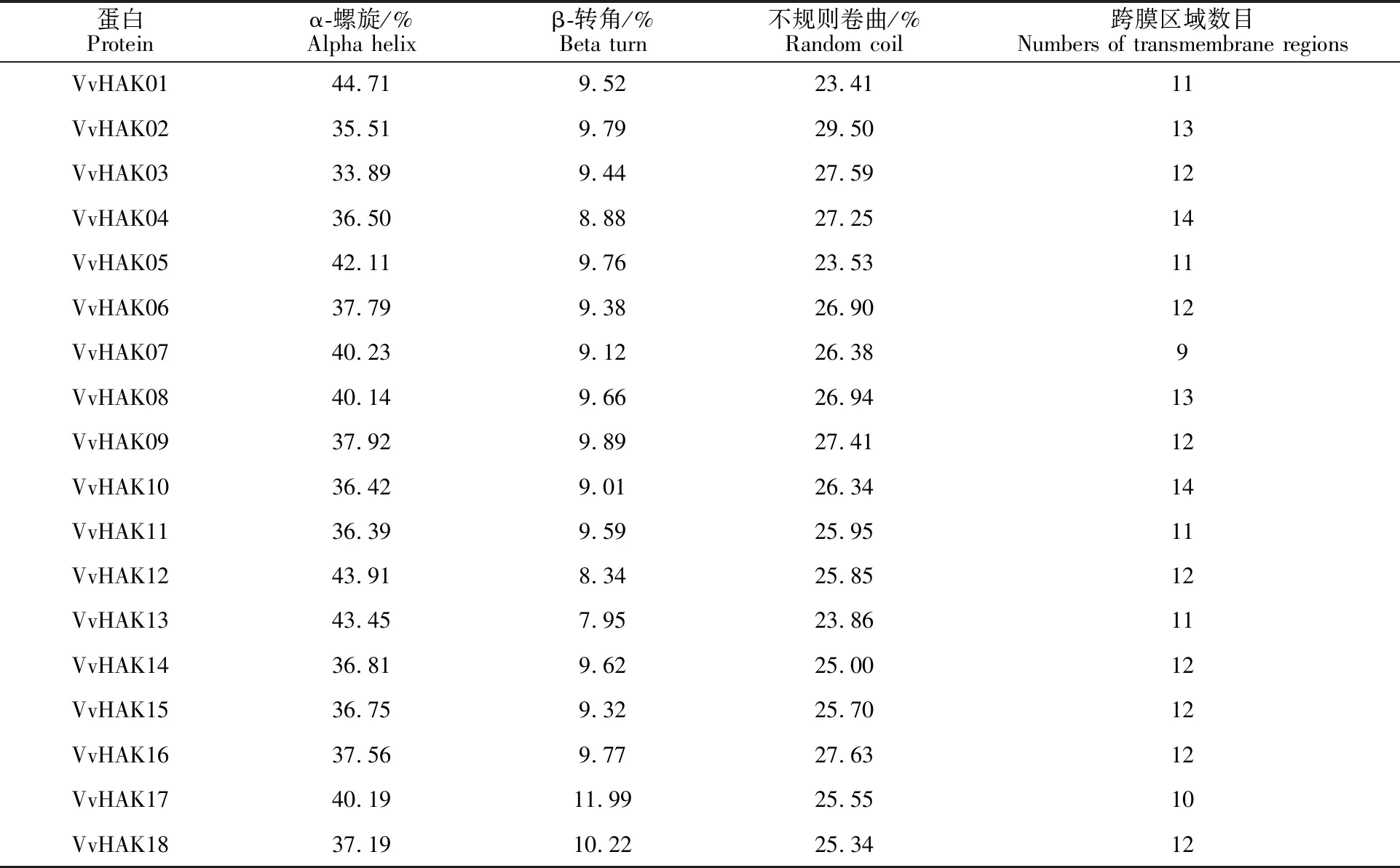
蛋白Proteinα-螺旋/%Alpha helixβ-转角/%Beta turn不规则卷曲/%Random coil跨膜区域数目Numbers of transmembrane regionsVvHAK0144.719.5223.4111VvHAK0235.519.7929.5013VvHAK0333.899.4427.5912VvHAK0436.508.8827.2514VvHAK0542.119.7623.5311VvHAK0637.799.3826.9012VvHAK0740.239.1226.389VvHAK0840.149.6626.9413VvHAK0937.929.8927.4112VvHAK1036.429.0126.3414VvHAK1136.399.5925.9511VvHAK1243.918.3425.8512VvHAK1343.457.9523.8611VvHAK1436.819.6225.0012VvHAK1536.759.3225.7012VvHAK1637.569.7727.6312VvHAK1740.1911.9925.5510VvHAK1837.1910.2225.3412

表4 葡萄HAK基因亚细胞定位预测Table 4 Subcellular location prediction of VvHAKs
注:- 表示无表达;Cyto代表细胞质;E.R代表内质网;Chol代表叶绿体;Mito代表线粒体;Vacu代表液泡膜;Plas代表细胞质膜;Nucl代表细胞核;Extr代表细胞基质;Pero代表过氧化物酶体;表内的数值代表准确率,分值越高,代表预测的准确性越高。
Note:- represents no expression; Cyto represents cytoplasm; E.R represents endoplasmic reticulum; Chol represents chloroplast; Mito represents mitochondria; Vacu represents tonoplast; Plas represents membrane; Nucl represents nucleus; Extr cells represent the matrix; Pero represents peroxisome; numerical representation form accuracy rate,the higher the score,the higher the accuracy of predicted.
2.6 顺式作用元件分析
对葡萄HAK基因家族的顺式作用元件进行分析表明(表5),VvHAKs中GroupⅠ的5个成员VvHAK05、VvHAK14、VvHAK15、VvHAK16和VvHAK18均存在ABA响应元件,其中VvHAK16还存在DRE和LTRE元件,说明葡萄HAK GroupⅠ的功能可能与ABA信号转导相关。VvHAKs中GroupⅡ的9个成员,分别为VvHAK01、VvHAK02、VvHAK03、VvHAK04、VvHAK06、VvHAK08、VvHAK09、VvHAK11和VvHAK13,它们均含有LTRE元件,与GroupⅡ在植物体内参与植物生理活动的规律一致,其中VvHAK04、VvHAK08、VvHAK09、VvHAK11和VvHAK13含有ABA和DRE响应元件,说明葡萄HAK基因家族GroupⅡ的功能除了参与植物生理作用外,可能参与信号转导。VvHAKs中GroupⅢ的VvHAK12不存在ABA响应元件、DRE和LTRE元件,只与转录因子的结合位点相关;GroupⅣ的VvHAK17中只有ABA响应元件,可能参与ABA信号转导。葡萄HAK基因上游2 kb区域所包含的顺式作用元件因所属亚族的不同,使元件表达具有差异性。同时发现,所有成员上游2 kb区域均存在MYB和WRKY转录因子的结合位点,说明VvHAKs的表达与MYB和WRKY转录因子作用密切相关,并且这些顺式作用元件可能单个发挥作用,也可能共同参与葡萄HAK基因家族的表达。
2.7 葡萄HAK基因芯片表达谱分析
对葡萄HAK基因芯片表达谱进行表达分析(VvHAK11除外),结果显示,葡萄HAK基因家族在受到盐、干旱、低温、高温、ABA及水分胁迫等非生物胁迫时,基因表达水平变化显著(图6)。VvHAK06在响应水分胁迫时表达量明显上调,而VvHAK17明显下调;VvHAK01、VvHAK15和VvHAK18在响应高温胁迫时表达量明显上调,而VvHAK06则呈现先下调再上调的趋势;VvHAK09和VvHAK13在响应高温胁迫时表达量呈现先上调再下调的趋势;VvHAK04、VvHAK06、VvHAK07、VvHAK08和VvHAK12在响应盐、干旱、高温胁迫下的表达量呈上调趋势。VvHAK17在响应盐、干旱胁迫时的表达量呈现先下调再上调的趋势。VvHAK10在响应低温胁迫时表达量呈现明显上调趋势,VvHAK15和VvHAK18在响应干旱和低温胁迫处理后呈现下调表达。葡萄HAK基因家族对ABA胁迫的响应不明显。
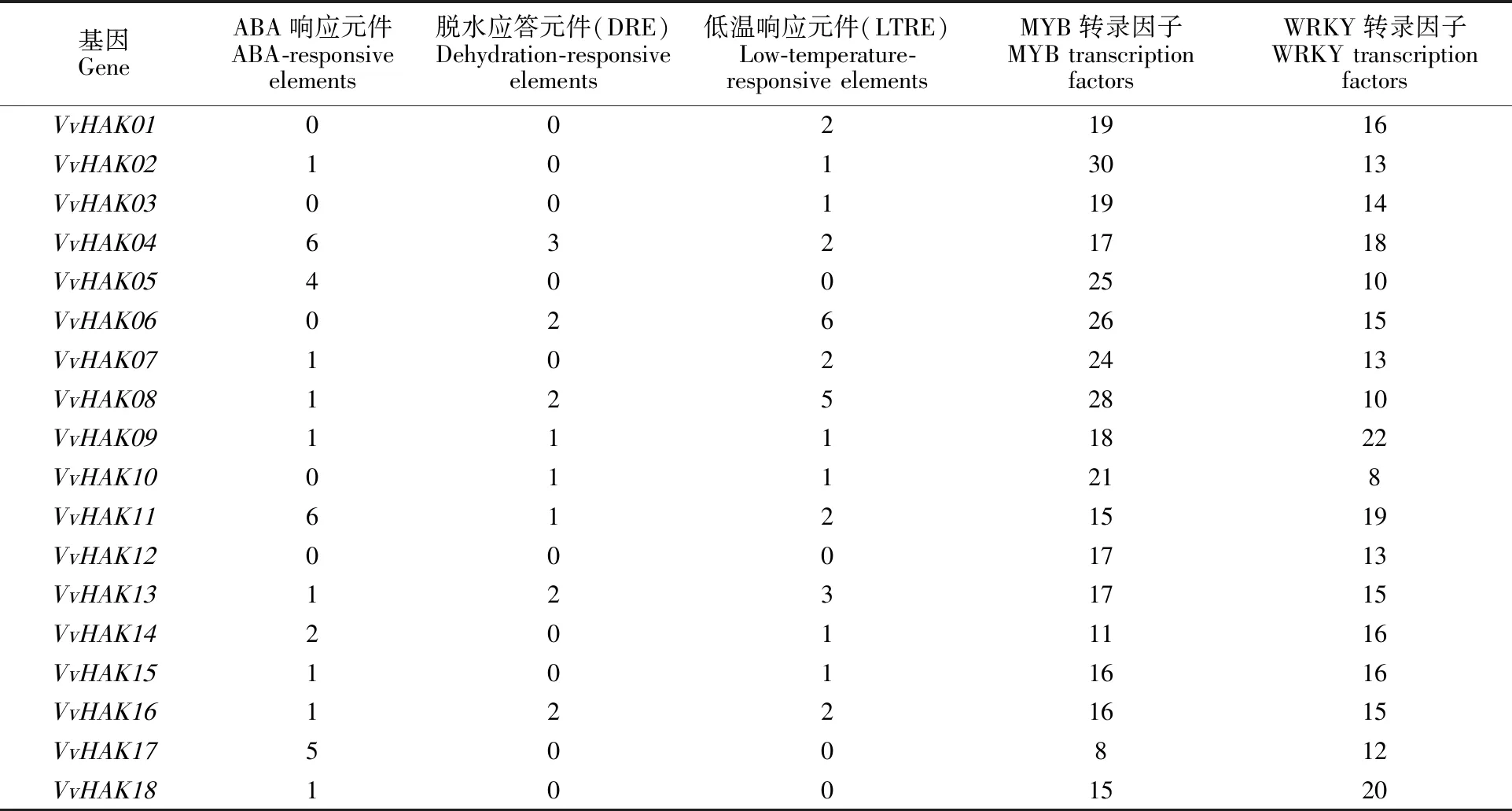
表5 葡萄HAK基因顺式作用元件的分布Table 5 Distribution patterns of VvHAKs cis-acting elements
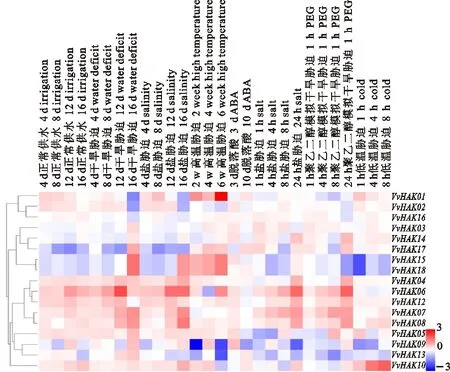
图中用红色、白色、蓝色代表基因表达水平。蓝色表示基因表达弱,红色表示基因表达强 Red,white and blue are used to represent gene expression levels.Blue indicates weak gene expression,red indicates strong gene expression
图6 葡萄HAK基因表达谱
Fig.6 Expression profile ofHAKgenes in grape
2.8 葡萄HAK基因家族成员在非生物胁迫下的表达分析
葡萄HAK基因家族在ABA、PEG和NaCl 3种胁迫下的表达量具较显著差异(图7)。VvHAK01、VvHAK05在50 μmol·L-1ABA处理6 h后,叶片表达量明显下调,其余均为上调;其中,VvHAK04、VvHAK06、VvHAK08、VvHAK09、VvHAK10、VvHAK13和VvHAK15同时在50 μmol·L-1ABA 和10% PEG处理 6 h后上调;VvHAK06、VvHAK07、VvHAK09、VvHAK10、VvHAK11和VvHAK13在50 μmol·L-1ABA 和400 mmol·L-1NaCl处理6 h后上调。VvHAK01、VvHAK02、VvHAK03、VvHAK04、VvHAK05、VvHAK08、VvHAK14、VvHAK15、VvHAK16、VvHAK17和VvHAK18在400 mmol·L-1NaCl处理6 h后叶片表达量明显下调,说明6 h内,盐胁迫在一定程度上抑制基因的表达。VvHAK03、VvHAK05、VvHAK07、VvHAK11、VvHAK14在10 % PEG处理6 h后叶片表达量下调,可能与其不存在顺式作用元件DRE有关,同时VvHAK03、VvHAK05和VvHAK14在400 mmol·L-1NaCl处理6 h后,叶片表达量明显下调,说明400 mmol·L-1NaCl抑制VvHAK03、VvHAK05和VvHAK14基因的表达。
3 讨 论
植物在发育过程中进化出一套保护机制,以此来应对干旱、高盐、温度和氧化等逆境胁迫。在这些不利因素下,植物可以通过启动相关基因的转录,形成具有不同功能的蛋白来保护细胞内的各种新陈代谢反应,进而维持植物体结构与功能的完整性[22]。
HAK转运体家族首次发现于细菌中[23],现已在植物中发现4个主要的钾离子转运基因家族,其中KUP/HAK/KT转运子家族是最大的基因家族[3]。本研究共得到18个葡萄HAK家族成员,Motif 分析将葡萄HAK基因家族分为4个亚族,VvHAKs成员主要以GroupⅠ和GroupⅡ为主,GroupⅢ和GroupⅣ中的成员相对较少。系统发育树中葡萄HAK的家族成员与拟南芥HAK基因家族成员有较高的同源性,且发现同一物种间的成员在进化树中分布较为集中,玉米HAK和水稻HAK这两个家族成员之间同源性较高。
已有研究表明:结构上存在数目不等的跨膜域,并且N端具有相对保守区域,是KUP/HAK/KT基因家族的共同特征;多序列比对发现VvHAKs的大多数成员在N端有相对保守区域,而且二级蛋白结构分析发现整个葡萄HAK家族均有至少9个跨膜区域,这与玉米基因家族二级结构分析类似[10],VvHAKs外显子数目和结构在不同亚族间差异很大,但同一亚族中编码区结构类似,这一规律与大豆HAK家族中的类似[24]。染色体定位表明,除了第7条染色体上的4个成员分布比较集中以外,其余8条染色体上的成员的分布相对分散,整体未表现出集中性。亚细胞定位结果表明,VvHAKs主要是在质膜上表达,这与其作为钾离子转运体的功能是一致的[24],且这个结果与林烟草中HAK基因的亚细胞定位研究结果相同[25]。在玉米和水稻中的多数成员中存在ABRE作用元件,这些作用元件可能会在信号转导中起着重要的作用,而ABA在生物体内最主要的功能是作为激素参与植物对外界胁迫的适应。在植物对非生物胁迫应答中,胁迫诱导基因的表达存在ABA依赖途径,在这些基因的启动子区域有一些保守的ABA反应元件,ABRE作用元件对一些受ABA诱导的抗逆基因的表达起调控作用,对基因响应ABA诱导至关重要[26]。
芯片表达谱分析表明,30 ℃高温条件下,VvHAK01、VvHAK15和VvHAK18的表达量最高,表明其对葡萄浆果成熟具有正调控作用;VvHAK06在响应水分、高盐、干旱及高温胁迫时,在葡萄茎尖和叶片组织中的表达呈现明显的上调表达趋势。定量分析表明,在不同胁迫下葡萄HAK基因家族成员在叶片中的表达程度不同。VvHAK13在50 μmol·L-1ABA、10% PEG处理6 h后,叶片表达量显著上调;VvHAK05在50 μmol·L-1ABA、400 mmol·L-1NaCl、10 % PEG处理6 h后,叶片表达量显著下调;与芯片表达谱分析一致。VvHAK07和VvHAK09在50 μmol·L-1ABA、400 mmol·L-1NaCl处理6 h后显著上调;VvHAK09的表达与芯片表达谱分析一致。VvHAK01、VvHAK05在50 μmol·L-1ABA、400 mmol·L-1NaCl处理6 h后显著下调;与芯片表达谱分析一致。VvHAK03、VvHAK05和VvHAK14在 400 mmol·L-1NaCl、10 % PEG处理6 h后显著下调;VvHAK05和VvHAK14与芯片表达谱分析一致。拟南芥盐胁迫后AtHAK06和AtHAK11的表达显著增加,而AtKUP02的转录则显著下降,在低盐的胁迫下AtHAK05和ThHAK05都受到Na+的抑制[27],HvHAK1和NrHAK1在100 mmol·L-1NaCl处理后表达上调,盐胁迫对SeHAK1转录水平的影响因盐角草的发育时期以及盐胁迫处理时间和浓度的不同而异[28]。葡萄HAK基因家族中的11个成员在高盐胁迫后,叶片的表达量下调,有可能是HAK基因家族在葡萄叶片中表达较少或者是在生长期表达量低,也可能是400 mmol·L-1NaCl浓度超过葡萄HAK基因家族耐盐程度。植物生长过程中基因表达水平的变化实质上是受到植物激素的调节。在实时荧光定量分析中葡萄HAK基因家族成员的表达均受ABA调节,表明ABA对HAK/KUP/KT基因的表达可能具有调控作用。
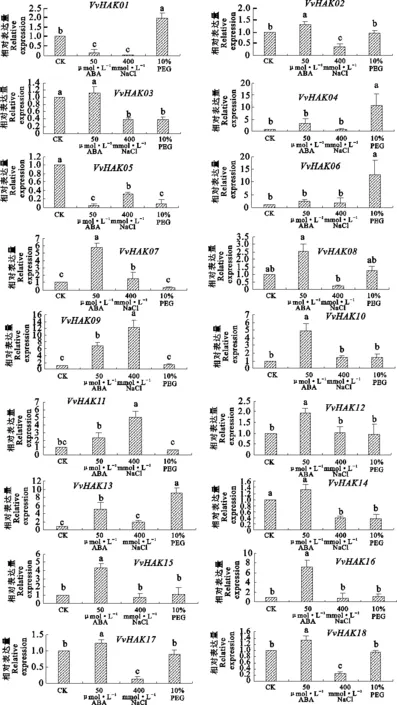
图7 葡萄HAK基因家族在不同处理下的相对表达水平Fig.7 Relative expression of VvHAKs under different treatments
