高等食用真菌集硒特性研究进展
2019-07-01向戌莲张驰
向戌莲 张驰
摘要:硒(Se)是人体重要的必需微量元素,具有营养、解毒、致毒三重生物学功能。硒的地理分布极不均衡,食物是最重要的直接硒源。微生物在硒的地球化学循环中发挥着重要作用,高等食用真菌作为一种很好的富硒载体,在富硒产品的开发上具有广阔的发展前景。从硒、富硒食用菌、富硒作用、硒形态、硒的吸收代谢机理方面进行了概述,分析了富硒高等食用菌目前的困境和展望,以期对此方面的研究工作和生产提供参考。
关键词:食用真菌;硒(Se);硒形态;代谢机理
中图分类号:S662.3 文献标识码:A
文章编号:0439-8114(2019)06-0005-04
DOI:10.14088/j.cnki.issn0439-8114.2019.06.001 开放科学(资源服务)标识码(OSID):
Abstract: Selenium is an important essential trace element in the human body. It has three biological functions of nutrition, detoxification and toxicity. The geographical distribution of selenium is extremely uneven and food is the most important source of direct selenium. Microorganisms play an important role in the geochemical cycle of selenium. As a good selenium-enriched carrier, higher edible fungi have broad prospects for the development of selenium-enriched products. The selenium detection, the selenium-enrichment effect of selenium-enriched edible fungi, the selenium form, and the mechanism of selenium absorption and metabolism were summarized, and the current difficulties and prospects of selenium-enriched edible fungus were analysed in order to achieve research and production in this area for reference.
Key words: edible fungus; selenium(Se); selenium form; metabolic mechanism
硒作为人体重要的必需微量元素,被国内外医药界和营养学界称为“生命的火种”,具有营养、解毒、致毒三重生物学功能[1]。硒分布不均匀的现实状况导致补硒成为人们的一项艰巨任务[2]。食用菌在将毒性强的无机硒转化为有机硒,大量生产硒产品方面占有很大優势,成为硒有机化的研究对象。自1817年瑞士化学家Jons Jakob Berzelius发现硒(Se)元素以来,硒经历了作为有毒物到人体必需的微量元素的角色转变。本研究将对富硒高等食用菌的主要种类、集硒方法的特点进行概述,以期为相关研究提供参考。
1 硒与食品
1.1 硒的生理功能
硒不仅是谷胱甘肽过氧化物酶的组成部分,参与免疫反应和眼中光转换电的过程[3],还能减弱镉、砷、汞等重金属和黄曲霉等毒素的作用[4]。硒作为良好的抗氧化剂、解毒剂成分的同时,在预防心血管疾病,降低各种癌症的风险方面起着重要作用[5]。
1.2 人体硒的需求量
目前,WHO对健康成人的每天硒推荐摄入量是55 μg,中国营养学会膳食硒营养素参考摄入量为60 μg/d。成年人每日摄入Se少于40 μg,即定义为硒摄入缺乏[6]。缺乏Se容易造成多种疾病,如儿童缺Se容易造成大骨节病,成人则容易发生克山病。通过测定人血、尿中的硒含量来调整食物硒的摄入量,正常人尿中硒含量为0.01~0.15 mg/L,血清正常硒含量为15~25 μg/L。
1.3 硒富集
人体摄入硒的主要来源是食物,天然食物特别是蔬菜中硒含量较低,而在中国居民膳食宝塔中,蔬菜的摄入量很高。因此,一般仅靠天然食物不能满足人体对硒的正常需要。硒盐毒理分析表明,有机硒毒性远低于无机硒,且生物利用率更高。所以,无机硒的有机化显得尤为重要[2]。
对中国大部分地区而言要富硒,即增加土壤中有效硒的含量,而对于极少部分高硒地区而言则要降低硒的生物可利用性。由于微生物参与了土壤中硒转化的各个方面,因此利用微生物来调节土壤硒的分布和形态很重要[3],而且相对于天然植物合成转化法历时长、成本高,目前最具有实际应用的转化方法是微生物合成转化法[5]。通过在食用菌生长的基料中加入无机硒盐,适宜条件下正常培养即可得到含有机硒的食用菌并开发相关富硒食品。食用菌产品不仅能有效富集硒,同时自身含有多种氨基酸、维生素等成分,深受消费者的青睐。
2 富硒食用菌
2.1 食用菌富硒种类
许多食用真菌不仅含有丰富的活性物质,味道鲜美,药、食用价值都很高,而且具有较强的耐硒、富硒能力[7,8]。可利用食用真菌作为硒的载体,培养富硒食用真菌,提高食用菌的价值,得到较为理想的有机硒补剂产品[9,10]。
2.1.1 平菇 覃晓娟等[11]通过比较13种平菇品种的耐硒能力,发现不同平菇菌株对亚硒酸钠的耐受力差异明显。祝传望[12]通过在平菇栽培料中添加硒酸精氨酸研究发现,平菇不同部位的富硒能力不同;硒浓度对菌丝和子实体的促进作用也不同,且富硒后子实体挥发物质醇类的含量增加。
2.1.2 香菇 成群[13]比较了香菇、平菇、姬菇3种食用菌的集硒能力,结果表明香菇的集硒能力最强。吴丹[14]也比较了富硒的香菇和平菇抗氧化活性的差异,说明不同种的食用菌集硒能力、效果不同。Nunes等[15]进行富硒香菇栽培试验,结果表明,Na2SeO3增加了氧化酶的活性,不影响水解酶活性,且呼吸活性与Na2SeO3添加量呈负相关。硒更多的是对氧化酶的作用,并能通过自身的作用减少香菇呼吸消耗,进而促进其生长和营养积累[16]。
2.1.3 灵芝 程红艳等[17]研究发现在液体培养中,亚硒酸钠可促进菌丝体的生长,但对菌体合成多糖没有明显的促进作用;栽培培养中,亚硒酸钠对灵芝的生长和多糖的合成均有一定的促进作用;富硒发酵生产中,硒对灵芝菌丝产量无明显的提高作用,但能提高蛋白质的含量。硒在不同的生产方式中添加,作用效果不同[18]。
赵镭[19]研究发现,富硒灵芝蛋白质含量与其含硒量呈正比,相对于普通灵芝,富硒灵芝菌丝体中各种氨基酸含量更高,而且铁、镁等含量更高,锌、铜、钙等重金属含量则较低[20]。因此,硒的加入不仅提高了有益成分,还降低了重金属的有害成分。
2.1.4 黑木耳 周丽洁等[21]对黑木耳培养基及最适硒浓度进行筛选,得到麦麸30 g、黄豆粉15 g、葡萄糖20 g、琼脂20 g、水1 000 mL为最适培养基,最适硒浓度为6.0~7.5 μg/mL,硒含量可达30.16 μg/mL。培养基的不同,硒的富集效果不同。
武芸[22]在培养料中加入低浓度亚硒酸钠溶液,使黑木耳富集硒,检测发现蛋白硒、多糖硒占总硒的质量分数分别为52.03%、22.64%,说明黑木耳中硒主要分布在有机物中,蛋白硒是黑木耳中硒的主要赋存形态。
2.1.5 金针菇 金针菇的多种营养成分含量在蔬菜中名列前茅,堪称一个微型的营养“宝库”。金针菇的子实体、菌丝体和培养液具有抗癌、益智、防治高血压和抗衰老的作用。
李华为等[23]对金针菇进行富硒栽培,结果表明培养基中硒浓度小于30 mg/kg时,对金针菇的生长有促进作用,富硒系数为2.8~9.9;硒浓度在40~200 mg/kg范围时,对金针菇的生长产生抑制作用;硒浓度高于200 mg/kg时,不适于富硒金针菇的培养。硒的作用效果和其浓度有关,且表现为双重性,还能抑制镉的富集[24]。
2.1.6 羊肚菌 张强等[25]从羊肚菌菌丝中分离出羊肚菌蛋白(MEP),在微波辅助下加硒。FTIR结果显示,Se-MEP中含有Se-O-C键,实现了MEP的硒化。但MEP与硒的结合机制、Se-MEP的构效关系尚未明确,有待于进一步深入研究。孟超等[26]、冮洁等[27]对羊肚菌多糖提取工艺、菌丝体富硒条件优化及其硒多糖抗氧化活性进行了研究。丁健峰[28]对羊肚菌菌种和培养基进行了筛选,采用响应曲面法设计试验,利用Design-Expert软件优化了富硒羊肚菌深层发酵工艺参数,且对硒多糖PN进行了红外结构分析,结果表明PN是α-糖苷键连接的吡喃多糖,硒元素可能是以H-Se形式存在于多糖的支链上,且分子链分支结构彼此缠绕,呈现多股紧密的螺旋结构。
2.2 硒的主要赋存形态
硒的常见化学形态有硒酸盐(SeVI)、亚硒酸盐(SeIV)、硒代蛋氨酸(SeMet)、硒代胱氨酸(SeCys)、硒多糖等,其中有机态的硒多糖、硒蛋白的研究最多、最深入[29-31]。
2.2.1 硒多糖 尚德静等[32]从灵芝加硒培养的菌丝中分离纯化得到SeGLP-1和SeGLP-2两种灵芝硒多糖,经红外光谱、核磁共振光谱和激光拉曼光谱分析表明,SeGLP-1和SeGLP-2均是由α-糖苷键连接的吡喃多糖,硒取代了灵芝多糖中-OCH3上的 -CH3,與O以双键的形式结合,从而形成硒氧双键O=Se=O结构,首次提出了硒进入灵芝多糖的分子机理。不同食物中硒多糖不尽相同[33],组成硒多糖的单体、硒多糖结构的差异决定了硒多糖的功能差异[34]。试验证明,金针菇中至少含有3种硒多糖,且由于硒的加入改变了原碱溶性多糖的构型[35,36]。
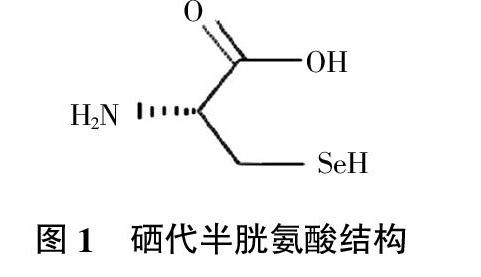
2.2.2 硒代半胱氨酸 硒代半胱氨酸(图1)不仅能合成硒代胱氨酸、硒代甲硫氨酸,通常还位于活性部位,与硒蛋白的众多生物活性有关[37]。但它的合成机理有些特殊,硒代半胱氨酸参与蛋白质合成受其mRNA的框内UGA密码子和下游链的硒代半胱氨酸插入元件(SECIS)的特殊茎-环结构所控制[38]。通常情况下,UGA是蛋白质合成的终止码,但mRNA的3′-非翻译区(3′-UTR)有SECIS时,UGA就成了编码Sec插入蛋白质链的三联密码子[39]。
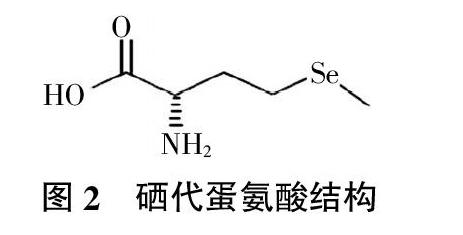
2.2.3 硒代蛋氨酸 当人体膳食供应硒不足时,机体会启动转硫途径将代谢库中的硒代蛋氨酸降解成硒代半胱氨酸,以供合成硒蛋白,而人体因缺少相关酶不能合成硒代蛋氨酸,所以,外源供应成了硒代蛋氨酸的惟一途径[40]。硒代蛋氨酸可以替代蛋氨酸渗入到蛋白质分子中,但这种作用是非特异性的。当生物体内蛋氨酸不足时,硒代蛋氨酸可以被诱导而替代蛋氨酸参与蛋白质的合成,取代后的蛋白质活性不会受到影响[41]。图2为硒代蛋氨酸结构。
2.3 吸收代谢机理
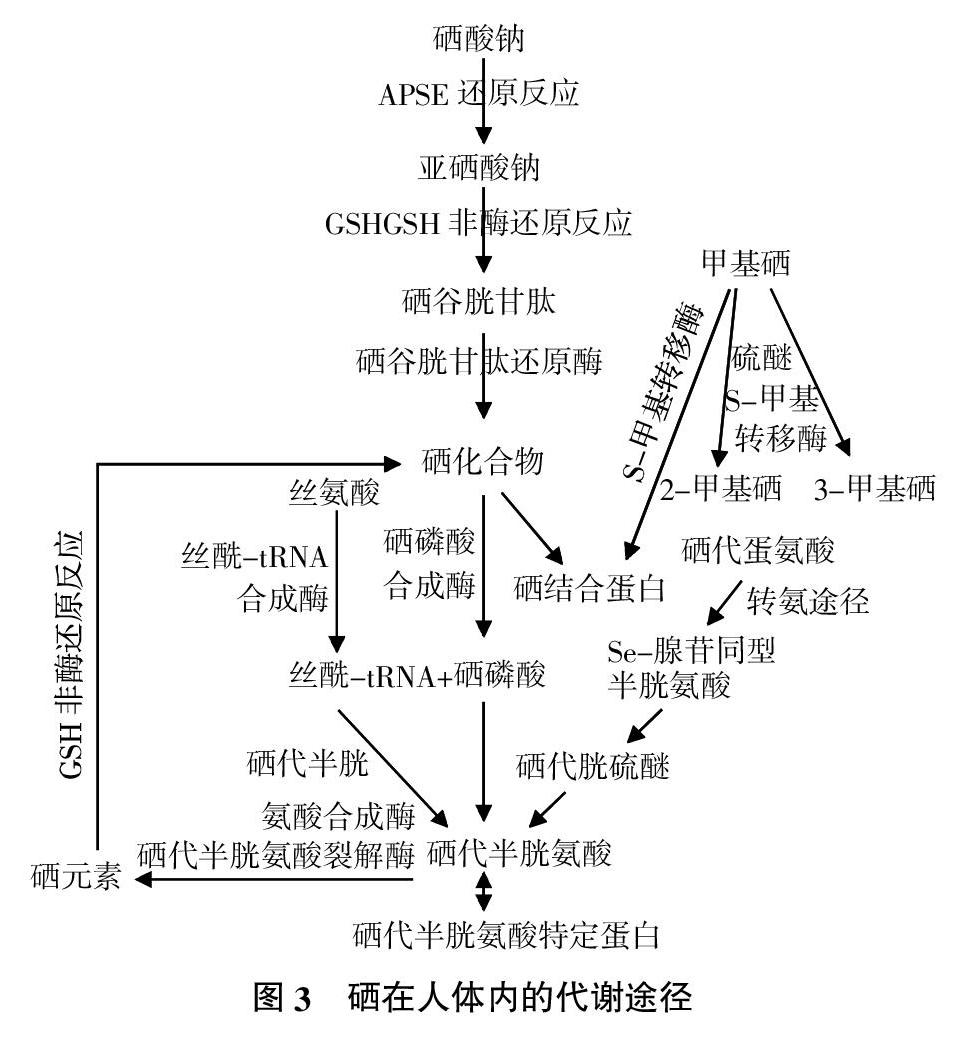
食用菌细胞壁上存在硫基、氨基等离子基团,生物吸附硒元素并通过菌丝细胞内的物质代谢转化,将无机硒吸收并与大分子活性物質结合,最终以硒蛋白和硒多糖等有机硒的形式存在[42]。Schrauzer[43]研究香菇细胞内硒的可能代谢机理为香菇菌丝通过主动运输或协助扩散方式将SeO32-或SeO42-转运至细胞内,无机硒盐通过氧化还原转化为H-Se-H,再进一步转化为硒半胱氨酸和硒甲硫氨酸。硒半胱氨酸可由特异密码子(UGA)介导代替半胱氨酸插入到蛋白中,构成酶活性中心。在甲硫氨酸的生化合成途径中,SeCys经过硒代胱硫醚和硒代高半胱氨酸(SeHomoCys)形成SeMet,硒甲硫氨酸可代替甲硫氨酸渗入蛋白质中构成硒蛋白。
微生物对硒的代谢主要包括硒的转运、还原、氧化、同化、甲基化等[44],硒的转运与硫酸盐或磷酸盐转运系统有关,还会受温度、pH、碳源、氮源等因素影响。而对于硒氧化的微生物了解甚少,缺乏硒氧化机制的系统研究。真菌中硒的同化机制已阐明,但目前尚未发现专一性的亚硒酸盐还原酶或其他亚硒酸盐还原关键基因,而在硒的甲基化研究中发现了一些关键的基因和酶。硒在人体内的代谢途径见图3。
3 问题与展望
3.1 机理研究
虽然高等食用菌中的硒含量、种类都可以测定,但硒的吸收、代谢机理尚不十分清楚。高浓度硒酸盐和亚硒酸盐可促使微生物进行异化还原并形成单质纳米硒颗粒,但是具有良好生物活性的纳米硒的生物合成机制也尚未研究清楚;另外,肠道微生物如何影响机体对不同形态硒的吸收与转化尚不清楚。目前,湖北省生物资源保护与利用重点实验室对植物硒吸收代谢分子层面研究已经取得不小成果,这将对食用菌方面的研究起到重要参考作用。
3.2 富硒食品安全性评价与国家标准
硒的最低摄入量与安全剂量的宽度相对较窄,对富硒食品的安全性检测和标准限量就显得尤为重要。由于不同地区的人群对硒的需求不同,不同年龄段、不同身体状况的人们补硒的标准也不太一样,而硒的安全剂量很窄[45],所以食品富硒国家标准很难界定。根据地方情况不同,地方相关部门有制定相关的地方标准。但同时缺少更多的安全性调查,由于饮食习惯差别等原因,对实际补硒状况需要进行调查,以供研究得到补硒的推荐食谱,合理的膳食搭配。补硒很重要,但不可盲目补硒,不然适得其反。总体来说,除了超高硒区,其他地区的人都需要补充一定量的硒。
3.3 室内栽培技术
在食用菌方面,已实现一系列的栽培技术突破,但对于珍贵食用菌的栽培技术难关还没有完全解决,比如羊肚菌的室内栽培技术。羊肚菌有“素中之荤”的美称,含有多种人体必需氨基酸,但由于生长条件苛刻,国内目前未实现人工培育,国外虽然有报道实现了室内培养,但并未实现商业化、大规模培养。总体而言,富硒食用菌的室内栽培技术研究尚不完备。
参考文献:
[1] 梁克红,卢林纲,朱 宏,等.食物中硒的研究进展[J].中国粮油学报,2018,33(3):134-138.
[2] 王庆华,黄 伟,李前勇,等.中国富硒食品的生产现状及趋势[J].广东微量元素科学,2008,15(3):7-10.
[3] 邹 宇,于俊林,徐 晶,等.硒及微生物富硒研究进展[J].食品研究与开发,2006,27(9):171-173.
[4] 吴珍和,祝寿嵩,陈德明.食用菌类中的硒元素[J].中国食用菌,1990(2):3-4.
[5] 秦恩华,唐巧玉.微生物富硒研究进展[J].安徽农业科学,2006(6):1212-1214.
[6] 田 芳,李 勇,张周洋,等.GB 5009.93-2017《食品安全国家标准食品中硒的测定》标准解读[J].食品安全导刊,2017(36):57.
[7] 凌宏通,宋 斌.富硒食用菌的研究进展[J].微生物学杂志,2008, 28(4):78-84.
[8] 黄春燕,张柏松,万鲁长,等.食用菌富硒培养研究进展[J].山东农业科学,2012,44(7):81-87.
[9] 刘书畅,马布平,周忠发,等.富硒食用菌的研究进展(综述)[J].食药用菌,2018,26(2):74-78.
[10] 李玲飞.四种食用菌的富硒特性研究[D].杭州:浙江大学,2012.
[11] 覃晓娟,熊建文,何 忠,等.广西主栽中高温平菇耐硒能力比较及生物富硒条件初步优化[J].南方农业学报,2012,43(9):1349-1352.
[12] 祝传望.平菇生物富硒及富硒平菇蛋白的差异表达[D].天津:天津科技大学,2015.
[13] 成 群.富硒培养基栽培富硒食用菌研究[J].陕西农业科学,2012(2):127-128.
[14] 吴 丹.富硒香菇多糖和富硒平菇多糖体外抗氧化活性研究[J].安徽农业科学,2010,38(11):5841-5843,5856.
[15] NUNES R G F L,DA LUZ J M R,FANTUZZI E,et al.Regulation of respiratory and ligninolytic enzyme activity of Lentinula edodes by selenium[J].Advances in microbiology,2013,3(8):31-36.
[16] 乔艳明,陈文强,徐 晖,等.富硒香菇的研究现状及应用展望[J].陕西理工学院学报(自然科学版),2015,31(6):46-50,56.
[17] 程红艳,孙绪春,孟俊龙,等.灵芝富硒栽培研究[J].安徽农业科学,2012,40(5):2626-2627.
[18] 胡海涛,袁林喜,尹雪斌,等.耐硒驯化灵芝菌种的液态发酵中硒的富集特征[J].食品与发酵业,2012,38(8):7-11.
[19] 赵 镭.灵芝生物富硒及富硒灵芝硒蛋白的分离纯化和抗氧化性研究[D].北京:中国农业大学,2004.
[20] 全卫丰,何静霞,汪 洁,等.原生质体融合选育富硒高产灵芝的研究[J].食用菌,2012(3):19-21,31.
[21] 周丽洁,亢学平,胡志强,等.延特5号黑木耳富硒培养基筛选初报[J].江苏农业科学,2012,40(11):259-260.
[22] 武 芸.富硒黑木耳中硒的分布规律及赋存形态的初探[J].食用菌,2008(5):5-6.
[23] 李華为,铁 梅,张 崴,等.金针菇子实体富硒栽培特性及HPLC-ICP-MS法对硒的分布研究[J].菌物报,2012,31(1):86-91.
[24] 李优琴,吴素玲,王雪峰,等.吸收剂和硒制剂对金针菇镉富集的控制[J].江苏农业学报,2011,27(6):1363-1366.
[25] 张 强,王松华,孙玉军,等.羊肚菌蛋白的硒化修饰及其体外抗氧化活性[J].精细化工,2017,34(11):1252-1259.
[26] 孟 超,史洪舰,赵 静,等.富硒羊肚菌多糖提取工艺的研究[J].中国西部科技,2013,12(2):83-84.
[27] 冮 洁,麦海美,解 彬,等.羊肚菌菌丝体富硒条件优化及其硒多糖抗氧化活性研究[J].食品与发酵工业,2016,42(9):120-125.
[28] 丁健峰.羊肚菌富硒深层发酵工艺及产物功能性研究[D].长春:吉林大学,2014.
[29] 铁 梅.食用菌中硒的形态分析[D].上海:华东师范大学,2006.
[30] 丁 莉,武 芸,尚红喜.植物体内含硒生物大分子基础与应用研究进展[J].湖北民族学院学报(自然科学版),2004,22(2):1-4.
[31] 黄 峙,郑文杰,郭宝江.含硒生物大分子化合物研究进展[J].海南大学学报(自然科学版),2001,19(2):169-175.
[32] 尚德静,王关林,王孝敏,等.两种灵芝硒多糖分离纯化及性质鉴定[J].大连理工大学学报,2001,41(2):165-168.
[33] 李锦金,盛家荣,邹修文,等.硒多糖的研究进展[J].广西师范学院学报(自然科学版),2014,31(1):41-45.
[34] 关 力,李永峰.微生物的天然多糖及其组成结构[J].中国甜菜糖业,2007(2):41-44.
[35] 张文州,许 嵘.食药用真菌多糖的研究进展[J].食品工业科技,2014,35(15):395-399.
[36] 饶圣宏.硒多糖的研究进展[J].黑龙江科技信息,2016(33):174.
[37] 丁诗华.硒蛋白合成的特殊机制[J].生物工程进展,1999(2):38-41.
[38] 周宝宏.终止密码UGA编码硒代半胱氨酸[J].生命的化学,1988(4):26.
[39] 向天勇,杨三东,吴永尧.硒代半胱氨酸的生物合成及参入[J].湖北民族学院学报(自然科学版),2005,23(2):114-117.
[40] 孙正祥,王瑞霞.食用菌中生物活性蛋白的研究进展[J].食用菌学报,2009,16(2):85-90.
[41] 黄 峙,向军俭,郭宝江.硒蛋白的分子生物学研究进展[J].生物化学与生物物理进展,2001(5):642-645.
[42] 徐巧林,吴文良,赵桂慎,等.微生物硒代谢机制研究进展[J].微生物学通报,2017,44(1):207-216.
[43] SCHRAUZER G N.Selenomethionine:A review of its nutritional significance,metabolism and toxicity[J].The American society for nutritional sciences,2000,130(7):1653-1656.
[44] 朱燕云,吴文良,赵桂慎,等.硒在动植物及微生物体中的转化规律研究进展[J].农业资源与环境学报,2018,35(3):189-198.
[45] 卿 艳,张立实.硒毒性研究进展[J].预防医学情报杂志,2012,28(3):216-218.
