肉豆蔻科4种植物幼苗生长与光合生理特性研究
2019-06-05张凤良李小琴者国雄毛常丽
张凤良 李小琴 杨 湉 者国雄,2 毛常丽 赵 祺 吴 裕
( 1. 云南省热带作物科学研究所,云南 西双版纳 666100;2. 西南林业大学林学院,云南 昆明 650233)
植物与环境的关系体现在植物生长发育的各个环节,包括其生活史、物候、形态及生理生化等多个方面,不同植物种类会随生态环境形成不同的适应策略。目前,国内外对植物性状以及它们与环境因子之间的关系进行了大量的研究[1-4]。在所研究的各类性状中,叶片性状、光合生理等与植物的生长及生物量的合成关系最为密切[5-7]。叶片性状可划分为功能型性状和结构型性状2类,功能性状如叶片结构及与植物碳的同化等与生长代谢密切相关的性状,它随时间和空间的变化复杂多变,这些性状之间的协同变化关系使得植物的叶片在一定的环境条件下能以最小的资源投入获得最大的碳同化能力[8];结构型性状主要包括叶片大小、比叶面积(SLA)、叶片干物质含量及叶厚等,其中比叶面积是最重要的结构性状之一,现已成为植物比较生态学研究的热点性状之一[9-10]。光合作用是植物生长和对环境变化响应的主要决定因子之一[11],它不仅能反映植物在不同光环境下的存活和生长,还能说明植物对长期环境变化的适应能力。植物叶片结构特征和光合生理共同体现了植物的生长策略和对资源的利用能力,是植物与环境长期互作的结果。因此,研究同一地域或不同地域不同植物光合和叶片特征之间的差别与联系,对于理解和预测植物对资源利用及对生态功能结构影响具有重要的意义[12]。
肉豆蔻科(Myristicaceae)是热带雨林主要代表树种,对研究热带雨林区系构成、地理分布和生态习性有重要价值,其中一些种类具有很高的经济价值,多种植物的种子含以肉豆蔻酸为主的油脂,是化工用油重要材料。肉豆蔻科全世界约有21属,近500种[13],主要分布在南美洲和中美洲、非洲大陆、马达加斯加及东南亚到西太平洋等4个地区[14]。我国有风吹楠属(Horsfieldia)、肉豆蔻属(Myristica)和红光树属(Knema)3个属10余种[15],分布于云南、广西、广东、海南、台湾等地,常生长于海拔1 000 m以下的热带雨林中,且以云南南部为主要分布中心,约占全国种类总数的80%以上。由于人类的破坏和盗伐,中国分布区内的肉豆蔻科植物野生种质资源数量日渐稀少。目前,有关肉豆蔻科研究较多的集中在分类学、形态学、油脂化学、资源调查和遗传多样性等方面,尤其是从油脂成分及应用上做了大量研究[16-23],对于这些濒危种、渐危种的生态适应性研究极少[24],而关于植株生长与形态、光合生理特性之间的关系研究未见研究。本研究对云南分布区内的肉豆蔻科资源进行调查、收集,共收集到风吹楠属3个种和红光树属1个种共4种幼苗的种子,并播种保存,本研究以这4种幼苗的实生苗为材料,对其1年生幼苗的生长、叶片性状及光合特性等进行了比较分析,以期为4种幼苗苗木适应性评价及资源保护和利用提供依据。
1 材料与方法
1.1 研究材料
于2016年4—6月,对西双版纳州景洪市(100°36′~101°06′E,21°41′~22°26′N)、勐腊县(101°00′~101°15′E,21°51′~22°00′N)、勐海县(100°02′~100°08′E,21°41′~21°55′N)、普洱市澜沧县(99°47′~99°53′E,23°13′~23°16′N)、德宏州盈江县(97°36′~97°51′E,24°29′~24°36′N)及临沧市沧源县(99°01′~99°05′E,23°14′~23°18′N)等6个县内肉豆蔻科植物资源进行调查,按同一地段内株间距离大于50 m以上的原则进行调查采样,采收到成熟种子的资源包括风吹楠(Horsfieldia glabra)、大叶风吹楠(Horsfieldia kingii)和琴叶风吹楠(Horsfieldia pandurifolia)及红光树(Knema furfuracea),采收种子带回,按株行距60 cm×80 cm直接播种于云南省热带作物科学研究所苗圃地内(混合播种),常规育苗管理,苗木生长良好。云南省热带作物科学研究所(100°40′~100°50′E,21°59′~22°01′N),海拔约 550 m,年平均气温18.6~21.9 ℃,最冷月份平均气温15.6 ℃,最热月份平均气温25.2 ℃,年均降雨量约1 200 mm,土壤为酸性红土,pH值4.5~5.5,适合风吹楠正常生长。
1.2 测定方法
1.2.1 生长和叶片性状测定
2017年8月中旬,分别对4种幼苗的苗木生长进行每株测定,用WKE3205型塔尺(苏州亿升五金工具有限公司,中国)(精确到0.01 m)测定株高,用IP67型游标卡尺(深圳市沈良精密测量有限公司,中国)(精确到0.01 mm)测定离地10 cm处地径。每个树种选择10株长势均匀的植株,每株采集主干中部健康叶片15片,测定指标及标准见表1,叶片性状分别采用直尺(精确到0.1 cm)及YMJ-D型叶面积测量仪(浙江托普仪器有限公司,中国)(精确到0.01 cm2)测量,叶片鲜质量及干质量用1/10 000天平称量,用烘箱烘干至恒质量。
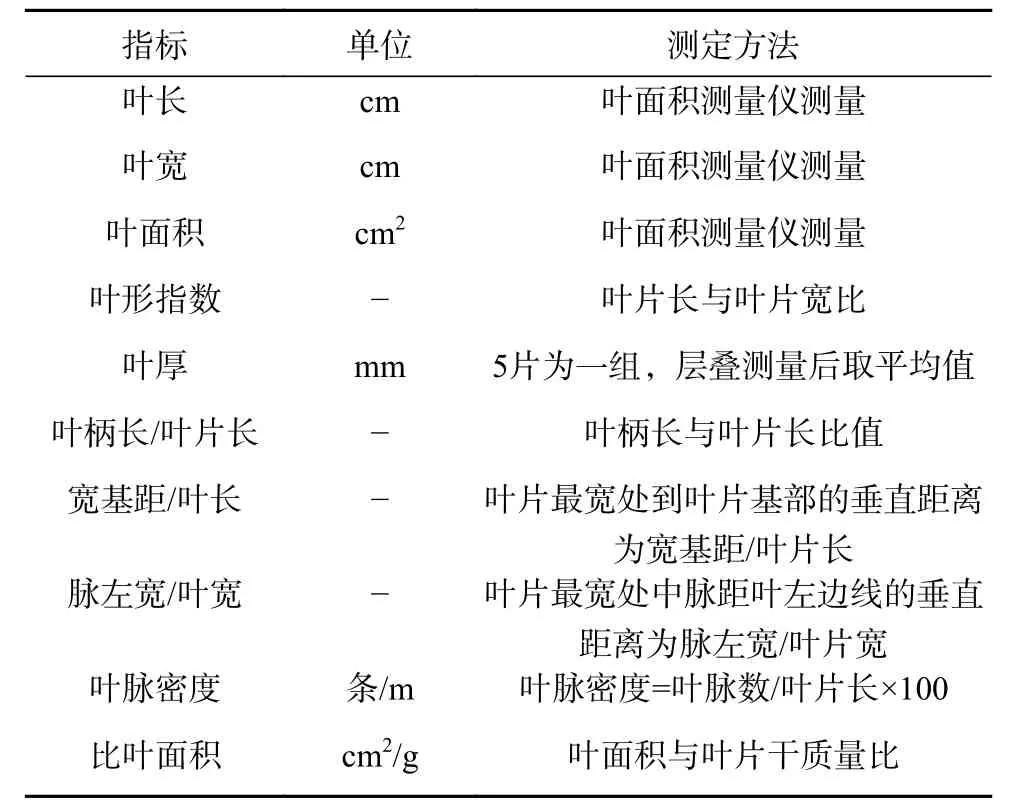
表 1 叶片性状指标与测定方法Table 1 Indexes and measurement method of leaf traits
1.2.2 叶片光合生理测定
1)叶片气体交换参数测定 于2017年8月下旬,选择晴朗无风的天气,利用LCpro-SD便携式光合测定仪(ADC Bioscientific Ltd.,英国)进行植株叶片气体交换参数测定,时间为上午9:30—11:00,测定过程中全程使用自然光,将光合有效辐射控制在 1 000 μmol/(m2·s),叶温设定 30 ℃,CO2浓度为400 μmol/m2,相对湿度为75 %。选择采集过叶片的植株作为测定光合生理的样株,每个种标记3个单株,选择向阳面主干中部Ⅰ级侧枝中部叶片,每株选择5片健康功能叶进行测定,数据稳定后读数。气体交换参数测定指标主要包括净光和速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)及蒸腾速率(Tr)等。叶片瞬时水分利用效率(WUEi),用净光合速率与蒸腾速率之比来表示。
2)光响应曲线测定 于2017年8月下旬,利用LCpro-SD便携式光合测定仪配套的LED红蓝光源叶室,在晴朗无风时,对4种幼苗的光响应曲线进行测定,测定时间为上午9:30—11:00。设定CO2浓度以当地环境CO2浓度为基准,气体流速控制在(500±0.1)μmol/s,样品室叶片温度控制在(30±3)℃,光合有效辐射(PAR)梯度设置为 1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50 μmol/(m2·s),每个光强下稳定2min后记录数据,3次重复。
利用叶子飘报道的双曲线修正模型进行4种植物光合—光响应曲线拟合[25],得出相应的光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)、表观量子效率(AQY)及决定系数(R2)。
1.3 数据分析
采用Excel 2007软件对数据进行录入、汇总并制图,用SPSS 23.0软件进行方差分析和Duncan多重比较。
2 结果与分析
2.1 肉豆蔻科4种幼苗生长性状比较分析
由表2可知,4种幼苗间苗高和地径差异达极显著水平(P<0.01),4种幼苗苗高生长排序从大到小依次为风吹楠>大叶风吹楠>琴叶风吹楠>红光树,地径生长与苗高生长一致,红光树生长量最小。从变异程度来看,红光树苗高变异最大,地径变异且最小,而大叶风吹楠、风吹楠和琴叶风吹楠较接近。
2.2 肉豆蔻科4种幼苗叶片性状比较分析
由表3可知,4种幼苗的叶片性状差异较大,其中叶长、叶宽、叶面积、叶厚、比叶面积及叶脉密度等指标种间差异均达到极显著水平(P<0.01),风吹楠、大叶风吹楠和琴叶风吹楠在宽基距/叶长、脉左宽/叶宽及叶柄/叶长间差异不显著,4种幼苗叶片主脉两侧均具有良好的对称性。4种幼苗叶面积大小排序为大叶风吹楠>琴叶风吹楠>红光树>风吹楠;4种幼苗比叶面积差异极显著(P<0.01),排序依次为琴叶风吹楠>风吹楠>大叶风吹楠>红光树,比叶面积在不同种间的变化趋势与叶片厚度相反,即叶片越厚,比叶面积越小;叶脉分布上红光树呈紧密状,大叶风吹楠最稀朗。以上分析说明,4种幼苗叶片大小呈现出了较大的差异,而对于叶片伸长生长方面,风吹楠、大叶风吹楠和琴叶风吹楠较相似,而红光树与之存在较大差异,叶片更狭长一些。
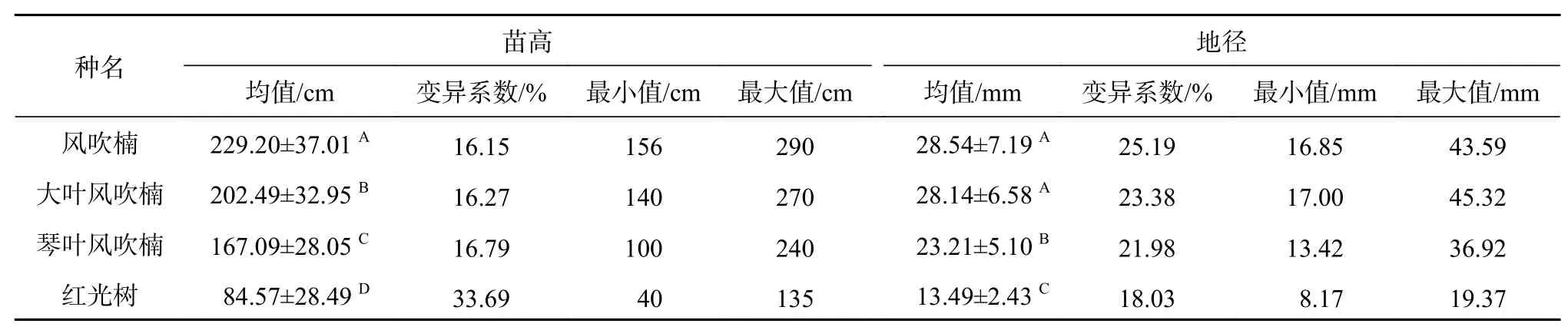
表 2 肉豆蔻科4种幼苗生长量统计Table 2 Statistics of growth in 4 species of Myristicaceae
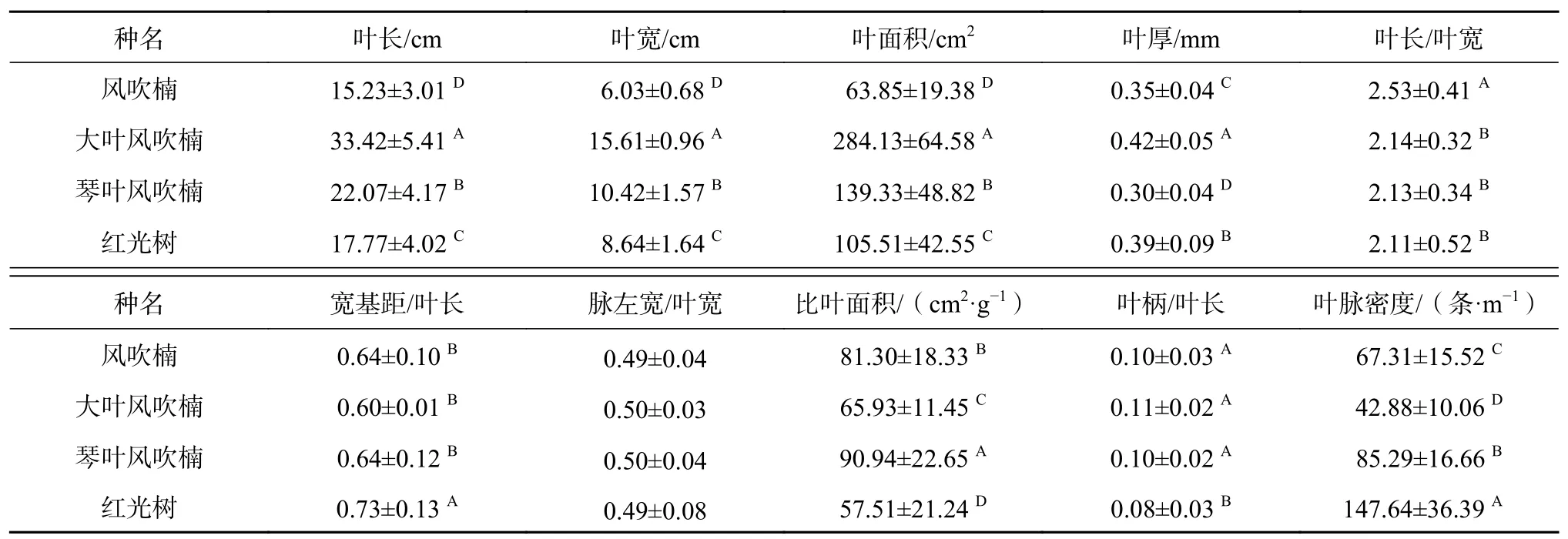
表 3 肉豆蔻科4种幼苗叶片性状比较Table 3 Comparison of leaf traits in 4 species of Myristicaceae
2.3 肉豆蔻科4种幼苗光合生理特性比较分析
2.3.1 肉豆蔻科4种幼苗叶片的气体交换参数
由表4可知,4种幼苗的Pn、Tr、Gs和Ci均达到极显著水平(P<0.01),WUEi差异达显著水平(P<0.05)。大叶风吹楠的Pn极显著高于其他3 个种(P<0.01),达到 8.35 μmol/(m2·s);琴叶风吹楠和风吹楠相当,其值分别为7.71 μmol/(m2·s)和 7.35 μmol/(m2·s);红光树最小,为6.88 μmol/(m2·s)。风吹楠、大叶风吹楠和琴叶风吹楠的Tr相当,显著高于红光树(P<0.05),进而导致红光树WUEi显著高于其他3个种(P<0.05)。4种幼苗Gs变化情况是琴叶风吹楠最大,风吹楠和大叶风吹楠相当,红光树最小,与Pn的变化基本一致;Ci变化与Pn趋势基本相反,红光树极显著大于其他3种(P<0.01)。
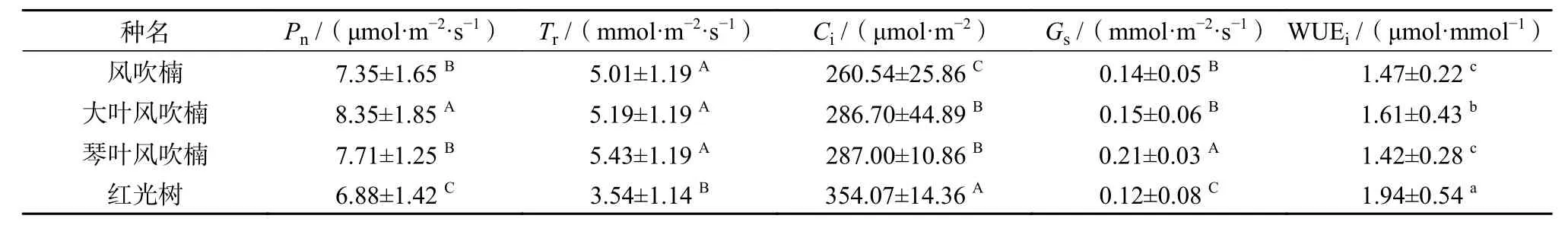
表 4 肉豆蔻科4种幼苗叶片气体交换参数统计表Table 4 Statistics of leaf gas-exchange parameter in 4 species of Myristicaceae
2.3.2 4种幼苗光响应曲线比较分析
由图1可知,4种植物幼苗叶片光合—光响应曲线拟合良好,总体趋势相似,整个过程大致可分为4个阶段。第1阶段(PAR为50~300 μmol/(m2·s))为Pn随PAR增加而呈直线上升阶段,即Pn迅速增加,这一阶段风吹楠和大叶风吹楠上升速度更快,其次为琴叶风吹楠,红光树最慢;第 2 阶段(PAR 为 300~900 μmol/(m2·s))为Pn随PAR增加略呈曲线缓慢上升阶段,仅有红光树 PAR 从 300 μmol/(m2·s)开始就逐渐保持平稳状态;第3阶段(PAR为900~1 500 μmol/(m2·s))几乎保持平稳状态,这时Pn几乎不再随PAR增加而变化过大,Pn基本达到饱和水平;第 4 阶段(PAR 超过 1 500 μmol/(m2·s))Pn随PAR增加而呈下降趋势,这时PAR过大可能对苗木产生了光合抑制。不同光强下,风吹楠、大叶风吹楠和琴叶风吹楠的Pn相当,均远高于红光树,4种幼苗中红光树的Pn一直处于较低水平。
前人对许多树种光响应曲线研究得出,最佳光合—光响应曲线拟合模型为直角双曲线修正模型[26-28],故本研究采用叶子飘[25]研
究的直角双曲线修正模型对4种幼苗的光响应曲线进行了拟合,并计算其特征参数(图1,表5)。在一定生长环境下,Pnmax反映了单叶光合能力的强弱,大叶风吹楠、风吹楠和琴叶风吹楠的Pnmax相当,均极显著大于红光树(P<0.01),总体红光树光合能力较弱,苗木生长较缓慢,植株矮小。各树种 LCP 为 10.03 ~ 16.92 μmol/(m2·s),LSP 为885.08~1 067.29 μmol/(m2·s),其中红光树和风吹楠具有较低的LCP和较高的LSP,说明这2个树种对光环境适应范围更宽,而琴叶风吹楠和大叶风吹楠对光的要求更高,在实际中观测到强光会对这2个种叶片造成灼伤,对光环境的改变更敏感。
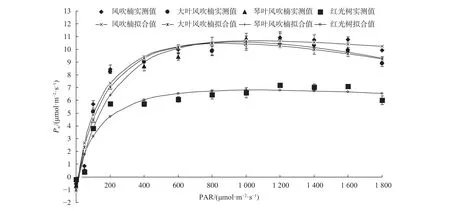
图 1 肉豆蔻科4种幼苗叶片光响应曲线Fig. 1 Light response curves of leaves in 4 species of Myristicaceae
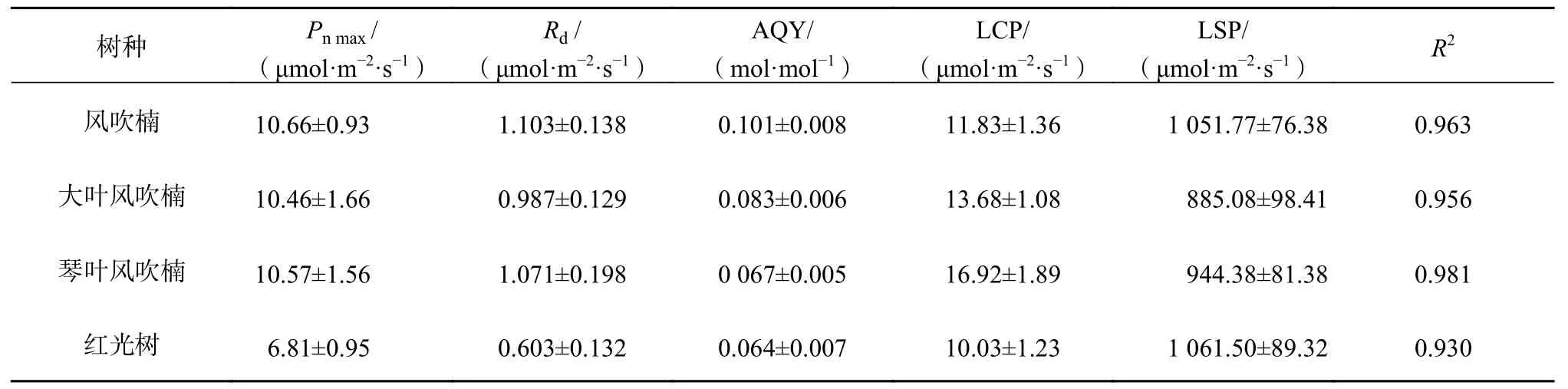
表 5 肉豆蔻科4种幼苗光响应曲线特征参数Table 5 Characterstica parameters of light response curves in 4 species of Myristicaceae
3 结论与讨论
生态环境因素和植物适应性是导致物种稀有和濒危的重要原因。由本研究及前人对云南分布区内肉豆蔻科野生资源调查资料可知,风吹楠生长于平坝疏林或山坡,居群落的中层;大叶风吹楠生长于雨林充足的沟谷密林及坡地密林中,零星分布,喜湿热环境,居乔木上层树种;琴叶风吹楠生长于热带雨林内水热条件较好沟谷密林中,喜湿热环境,属于群落中的主要建群种,居上层者多,下层零星分布;红光树多生长于沟谷密林中,居群落的中下层树种,零星分布[19,22,29]。在云南分布区,风吹楠多生长于山坡和山脊,土地干旱和贫瘠,光照充足;红光树多生长于密林,树体不大,长年光照不足;而琴叶风吹楠和大叶风吹楠居上层树种,树体高大,属于群落中的主要建群种,主要生长于沟谷或洼地,土壤相对湿润和肥沃。一定环境条件下,风吹楠、大叶风吹楠和琴叶风吹楠这3个种幼苗生长都较快,而红光树生长较慢,这与植物种性有关,即野生分布状态下,成年大树大叶风吹楠、琴叶风吹楠和风吹楠生长势粗壮而高大,红光树较矮小,前三者幼苗生长排序虽与成年大树有一定的差异,这可能是与林木的生长节律和生长环境有一定的关系,但总体上野生分布地属于高大的树种,其幼苗生长也较快,说明该区域适合4种植物的生长,其苗木的生长特性主要由植物种性决定。在实际观测中,发现局部遮阴严重的地方,风吹楠和大叶风吹楠苗木生长严重受影响,琴叶风吹楠次之,红光树受到遮阴胁迫,说明遮阴环境对这几种植物的生长可能影响比较大,具体影响程度如何有待进一步研究。
关于植物叶片性状与苗木生长及光合特性的关系前人做了大量的研究,得出苗木的生长与叶面积呈显著正相关,比叶面积大的植物,往往光合速率低,并指出叶的大小是一项可以遗传的性状[30-31]。从同一物种内部无性系或家系间做了关于叶片性状与生长的关系得出,生长快的苗木往往具有较大的叶面积、叶脉稀朗,但本研究中4种幼苗叶面积大小排序为大叶风吹楠>琴叶风吹楠>红光树>风吹楠,对应的大叶风吹楠和琴叶风吹楠单叶净光合速率也较高,其中大叶风吹楠比叶面积相对较小,苗木生长较快,琴叶风吹楠比叶面积较大,苗木生长相对大叶风吹楠小一些,这与前人研究的结论较一致;但风吹楠的单叶叶面积最小,比叶面积较大,苗木生长最快,这跟前人在很多树种上研究的差异较大,在实际测定中发现,风吹楠整株叶片总数显著大于其他3个种,结合气体交换参数综合分析可以推测风吹楠苗木整株Pn应该是4种幼苗中最高的;红光树叶片数量较少,叶片Pn最低,植株生长最慢。这就说明,种与种遗传背景差异很大时,仅从单一性状进行相关性分析,往往指标间会存在诸多的不一致,导致许多研究结果与实际存在较大差异,加之生物量的合成是长期积累的过程,与季节和年份间气候变化、生长环境及其林龄等关系密切。
由光响应曲线模型拟合的结果来看,Pnmax风吹楠、琴叶风吹楠和大叶风吹楠相当,红光树最低。LSP风吹楠也较大,参数Pnmax和LSP综合反映植物在强光下的光合效率[32],可见风吹楠对强光有较强的适应能力,这可能是野生状态下长期生长于山脊的结果。红光树具有较高的LSP和较低的LCP,表明该种具有一定的耐阴性又适应阳光充足的环境,对光适应范围大,而红光树瞬时水分利用效率也较高,综合说明虽然红光树植株相对较小,但对环境的适应能力则较强。通过方程回归拟合计算出4种幼苗的AQY结果表明,风吹楠最大,红光树最小,而对应的LCP又相对较小,LSP相对较大,导致这3个指标评价出来的结果不一致,故重点通过4种幼苗间LCP和LSP的差异性来评价苗木的适应性,对AQY作为反映植物吸光性重要参数的讨论,在后续研究工作中将作进一步论证。
综上所述,红光树对光适应范围较大,植株生长相对慢一些,风吹楠适应范围次之,生长最快,而琴叶风吹楠和大叶风吹楠对光的适应范围相对窄一些,生长较快。在大田调查中发现,琴叶风吹楠苗期叶片容易在夏季高温时受到太阳灼伤,但这些树种具体适合怎样的生长环境,还需要结合不同的生态环境做深入研究。
