渔业对海洋生态系统的影响与生态系统水平的管理模式
2019-05-28黄大明邵海培李如意LarryB.Crowder
黄大明 邵海培 李如意 Larry B. Crowder
摘要:渔业是影响海洋生态系统的最大因素之一。渔业是人类活动的一部分,渔业管理正从单一种群水平管理模式向海洋生态系统水平管理模式过渡。系统分析是理解各组分之间的联系、建立生态系统动态模式和制定管理方法的关键手段。生态系统水平的管理模式不但要考虑顺捕、休闲渔业、个体渔业和环境变化的影响,还要考虑直接和间接影响。这些影响可能是巨大的,很可能是难以预料的。渔业、环境多样性和气候变化的协同效应日益威胁着海洋生态系统,同时提升了综合治理的复杂性。本文讨论渔业的全球性影响,并提出一个用于管理海洋生态系统的生态学框架。
关键词:个体渔业;顺捕;商业捕捞;直接和间接影响;生态系统水平的管理模式;食物网;生境影响;网络;休闲渔业
中图分类号:F326.4 文献标识码:A 文章编号:1671-2064(2019)24-0000-00
0 引言
虽然传统的海洋渔业管理致力于解决目标物种广泛减少的问题,但过度捕捞已经显著改变了整个食物网[20][56]。渔业对海洋生态系统中的食物网有各种直接和间接的影响,这些影响具有复杂和潜在的链锁效应。渔业的很大部分集中在对顶级捕食者的捕捞,而这种捕捞的强度是不可持续的。另外,饲料渔业捕捞位于食物网中部的优势种类,如白眼狭鳕(Theragra chalcogramma)或秘鲁鳀鱼(Engraulis ringens)等,捕捞减少了鱼类、海洋哺乳动物和海鸟等捕食者的猎物[83]。捕捞不仅去除食物网中特定营养级上的生物量,而且还有间接影响,如移除非目标物种、改变生境、调整生物行为以及向食腐动物提供福利。
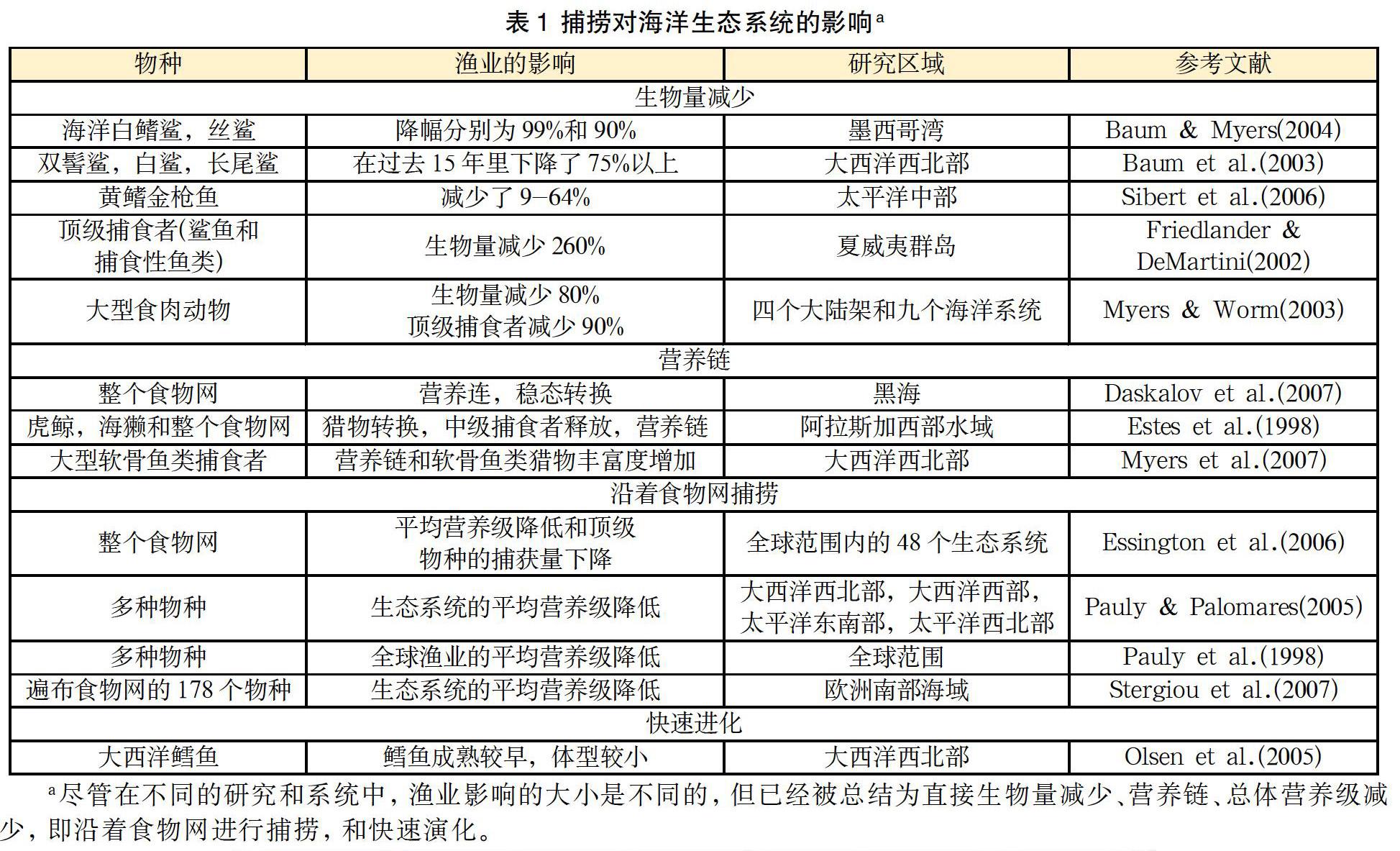
研究指出,渔业是改变海洋生态系统的最古老、最主要的因素之一[56]。渔业已经与其它人类活动一起导致了生物多样性的惊人损失,并会产生能在整个生态系统中传播的无法预料的影响[119]。现代渔业已捕捞90%以上的远洋捕食性动物[83](表1)。但是也有人认为这一观点:统计技术有缺陷、数据分析不完整[48][115]。保守估计认为接近50-70%,这一数据仍然为远洋捕食者大规模减少提供了证据[48]。Sibert等人分析了太平洋金枪鱼渔业全部可获得的数据,并得出结论:随着生物量减少9-64%,捕捞强度对渔业有巨大影响,具体程度取决于现存种群数量[100]。
尽管对于远洋中捕食性动物数量减少的幅度存在争议,但几乎没有研究人员对底栖捕食性动物数量减少90%甚至更多这一论断提出异议[20][97]。Friedlander和DeMartini发现,未被捕捞的夏威夷西北群岛顶级捕食性动物的平均生物量比夏威夷主岛高出260%以上,夏威夷主岛的顶级食肉动物和其它鱼类被大量捕捞[43]。还有,大西洋西北部和墨西哥湾沿岸的鲨鱼和远洋鲨鱼的种群数量急剧下降,某些种类的下降幅度高达99%[7][8][82]。在评估渔业(特别是沿岸和底栖渔业)的影响时,不能忽视顶级捕食者生物量的直接减少量这一指标。
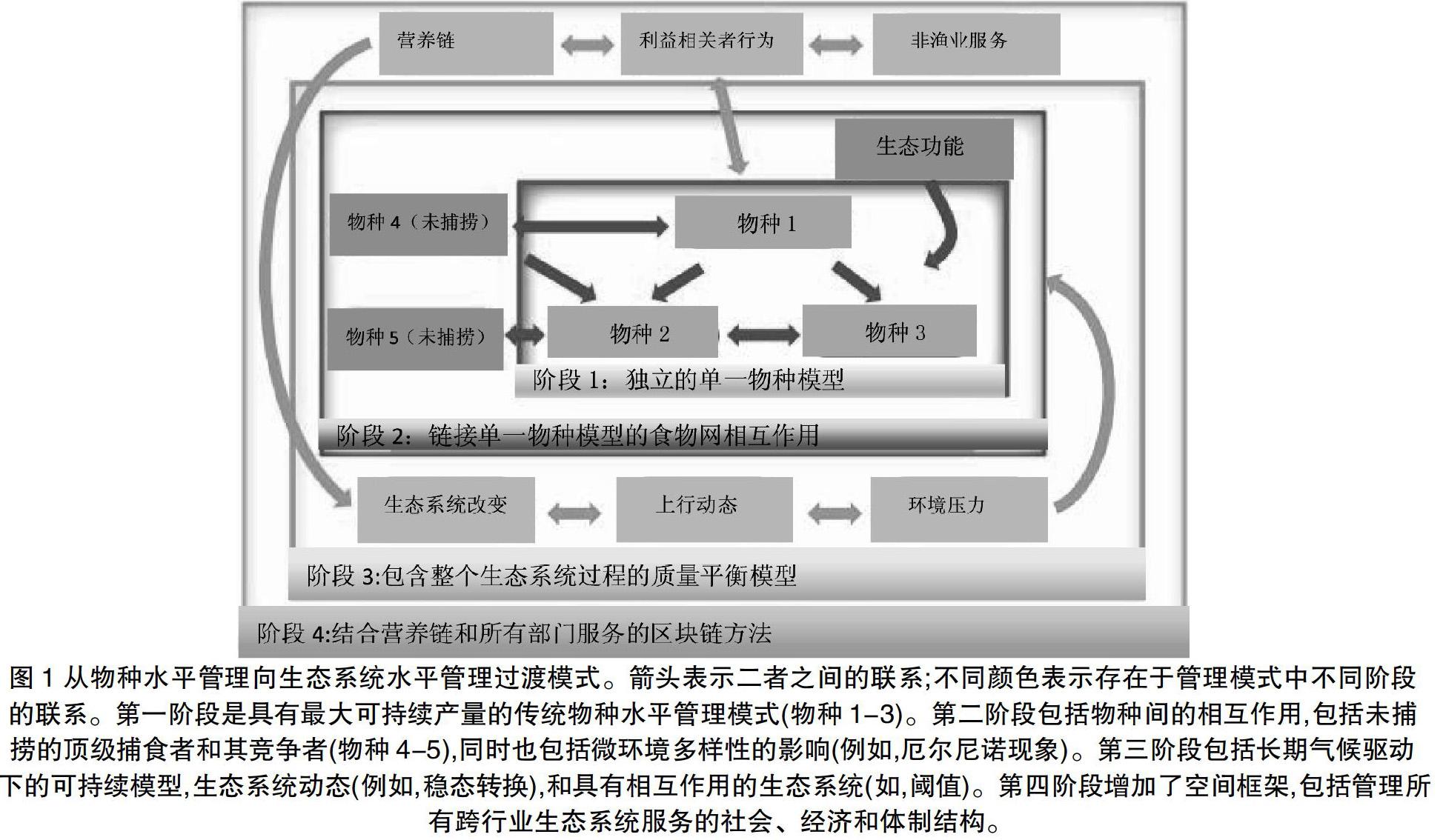
在定义并管理渔业的直接影响方面存在困难,更不用说间接和潜在的生态系统影响。在海洋生态系统水平管理海洋渔业应该将食物网作为生态系统中各组分相互作用的关键纽带[71]。虽然生态学家认识到了食物网理论、关键种和强相互作用体在生态系统中作用的重要性,但传统的渔业管理仍然只关注单一目标物种,并采用单一物种水平的管理方法[6]。尽管最近强调海洋生态系统水平的管理方法的必要性,但在如何推进该办法的实施问题上几乎没有一致看法[76][93]。如:ECOPATH、ECOSIM等模型 (图1)。
箭头表示二者之间的联系;不同顏色表示存在于管理模式中不同阶段的联系。第一阶段是具有最大可持续产量的传统物种水平管理模式(物种1-3)。第二阶段包括物种间的相互作用,包括未捕捞的顶级捕食者和其竞争者(物种4-5),同时也包括微环境多样性的影响(例如,厄尔尼诺现象)。第三阶段包括长期气候驱动下的可持续模型,生态系统动态(例如,稳态转换),和具有相互作用的生态系统(如,阈值)。第四阶段增加了空间框架,包括管理所有跨行业生态系统服务的社会、经济和体制结构。
生态系统水平的渔业管理需要考虑到食物网中的相互作用、气候动态因素和专业管理措施。强相互作用体通过捕食和竞争等生态过程影响食物网。当受到渔业影响时,强相互作用可以将影响扩大成对整个食物网的影响。渔业的影响可以通过食物网向上和向下传递,导致物种多样性、总生物量和生态系统服务功能的减少[79]。最近一个假说认为,具有强相互作用和高度专门化的食物网(例如,低杂食性)最容易因为渔业的影响而崩溃,而更复杂的食物网具有功能冗余和韧性[6]。海洋食物网从三个低多样性的营养级(如南极浮游生物→磷虾→鲸鱼)到复杂的热带系统(加勒比地区有249个营养类群)[6]各不相同。健康的生态系统通过增加食物网间的联系(物种间的相互作用)和提高功能多样性(每个营养级的物种数量)来缓冲系统崩溃造成的影响,但它们也提供生态系统服务(即,人类获得的好处)[74][80]。最近研究认识到一些营养级间潜在机制以及这些生态效应产生的潜在的捕食或竞争影响,但重要的是将对渔业所产生影响的理解扩展到商业捕捞之外[18][22][32][84][98]。本文研究商业捕捞的影响包括渔业对生境和生态系统的影响。海洋生态系统水平管理方法应该包括非目标渔获②(顺捕③)、休闲渔业④、个体渔业⑤的影响。海洋生态系统管理方法随数据控制能力的丰富而不断提升。
1 渔业对海洋食物网的直接和间接影响
商业捕捞通过直接移除食物网的关键组分,导致整个海洋食物网发生巨大变化。随着顶级捕食者的减少,捕捞压力继续向低营养级的方向转移,这个过程被称为沿着海洋食物网捕捞[89][90]。以低营养级物种为目标的渔业的发展将会进一步降低被捕捞鱼类的平均营养级[37]。对饲料鱼类(包括浮游生物)捕捞的增加也可能导致利用者冲突,例如油鲱(Brevoortia tyrannus)的商业利益和休闲利益之间的冲突[96],或渔业和受保护物种之间的利益冲突,例如底栖鱼在渔业捕捞的同时被北海狮食用。在美国东海沿岸,鲱鱼因其鱼粉和鱼油的商业价值而被捕捞,它们也是条纹鲈鱼(Morone saxatilis)等捕食鱼类的重要猎物。
沿着和通过海洋食物网捕鱼产生平均营养级改变的结果[104]。比较渔具对渔获平均营养级的影响,发现带有大钩的长线能够选择性地去除最高营养级,带来最高的潜在生态系统效应[104]。Friedlander和DeMartini认为,如果以同等比例捕获较低的营养级,有效地切割营养级三角的一侧而不是将其“斩首”,将有可能减少捕捞顶级捕食动物的负面影响[43]。但这种情况很少发生。更为常见的模式是对大型捕食动物的过度捕杀。例如,按生物量计算,夏威夷群岛西北部的禁渔区域内顶级捕食性动物占50-60%,而这一比例在夏威夷本岛是2-3%。食物网的改变可能导致营养级链稳态转换,或者在最坏的情况下,食物网崩溃[71][84][119]。通过各种数据分析,包括物种多度的时间序列数据、食物结构数据、实验性捕食者隔离的结果,Myers等人描述了一种可能的营养级链,他们认为这是大西洋西北部食肉性鲨鱼大规模减少(接近90%)的原因[82]。与此同时,牛鼻魟(Rhinoptera bonasus)的数量急剧增加,其捕食强度使在海湾的扇贝(Argopecten irradians)渔场迅速崩溃。
在多样的系统中,替代种可以增加物种多度,并占据原有物种移除后所空余的生态位[87]。陆地系统的一个例子显示,由于生境破碎,中级捕食性动物(野猫、负鼠和浣熊)的数量增加,而郊狼的数量减少[28]。在顶级捕食者减少后,中级捕食者的数量增加,称为中级捕食者释放(mesopredator release),并且可以通过食物网产生偶联效应。食物网可以从自上而下的控制⑥转变为自下而上的控制⑦,例如在加勒比珊瑚礁系統中,随着食植动物的减少,这种情况很常见[85]。当曾经复杂的食物网被简化时,生态系统可能会崩溃,即使有一定复原的可能,也很难成功。这方面最好的例子是,由于疾病爆发,在食植性海胆(Diadema antillarum)崩溃后,藻类控制了加勒比珊瑚礁;这一事件的发生可能是因为其它食植动物已经近乎灭绝,其中包括一系列海礁鱼类[9]。另一个例子是北大西洋鳕鱼灭绝后鱼类群落组成成分的变化:包括角鲨在内的小型食肉动物数量的增加,这可能是延缓鳕鱼种群恢复的原因之一[14]。
过度捕捞顶级捕食者可直接导致其猎物丰盛度增加,从而可能改变食物网的其余部分。例如,在大规模渔业盛行之前的黑海中,捕食者数量受系统生产力变化的控制。在20世纪60年代和70年代顶级食肉动物(食鱼鱼,包括mackerel,Scomber scomber)被大规模捕捞之后,营养级关联导致浮游生物丰富度增加,随后浮游动物减少,浮游植物增加。改变后的食物网为凝胶状浮游生物的爆发提供了合适的条件,而这些浮游生物现在主导着该生态系统,这是一种在受到干扰的生态系统中常见的模式[37]。
正如许多海洋哺乳动物种群,从食物网中段捕获饲料鱼类可以降低顶级食肉动物的捕食量[35]。一个相关的例子是“蜂腰动力学”,处于中间的浮游生物的丰盛度控制初级生产者和顶级捕食者数量,而不是自上而下或自下而上的控制。其它例子包括高产的秘鲁上升流系统。沙丁鱼(Sardinops sagax)和凤尾鱼(E. ringens)的丰盛度以上升流的强度为基础进行循环,在较冷的拉尼娜年中最强,在较温暖的厄尔尼诺年中最弱,进而影响顶级捕食者的分布:产生与渔获量相关联的趋势[4]。由于这些综合效应,许多顶级捕食者如海狮(Otaria flavescens)在生产效率较低的年份面临营养补充的减少和存活率的降低[102]。
渔业还可以改变生态过程,如生境选择和猎物婚配行为。例如,棕短鳍海鲫(Brachyistius frenatus)会改变它们的行为以应对捕食压力;在没有捕食者副鲈(Paralabrax clathratus)的情况下,棕短鳍海鲫对生境的选择和行为模式不同于捕食者存在时的选择与模式[3]。正如在海洋物种中观察到的那样,配偶选择和婚配行为的变化也可能受到捕食压力的影响,比如双色雀鲷(Stegastes partitus)[40]。与直接移除生物量的影响相比,这些影响可能较小,但却有仍然有可能妨碍生态系统的恢复,并进一步加速生态系统的衰退。
个体大小选择性的捕捞模式对海洋物种具有种群水平上的影响,高强度的选择可以改变捕捞种群的遗传组成。演化发生在暴露于相对短期的选择压力的种群中,渔业捕捞就是选择压力的一种[65]。Olsen等人指出,多年来大小选择性的捕捞模式似乎使得北方鳕鱼(Gadus morhua)较早成熟,体型较小[86]。在特定种群崩溃之前,观察这些趋势可以为管理者提供警示信号[86]。在实验室中经过多代大小选择性收获后的银鱼(Menidia menidia)显示出向生长缓慢的个体的可遗传变化[24]。种群的这种变化可能是不可逆的,如果我们要保护物种并且最大化产量,就需要在渔业管理方面融入进化性观念(例如,保护未成熟个体和最大的个体,进行渔区限制)[24][54][65]。
捕捞特定物种对其生活的生态系统的影响取决于该物种的生活史,移除的程度,密度依赖性补偿的可能性,其在群落中的作用以及生态系统中的功能冗余[84]。这些影响因自然表型的可塑性,动物行为变量,猎物转换,个体食物结构的变化,物种多样性以及猎物的数量反应和聚集反应而变得复杂[3]。食物网的改变是否具有长期的生态系统影响还取决于所捕捞物种的恢复能力[16][56]。 如果没有其它物种在被捕捞物种缺少的情况下占据其生态位,它们的猎物基础仍然完好无损,除非将其恢复,否则没有任何依赖性影响,那么暂时移除该物种可能不会产生重大的长期后果。该生态系统可能能够回到先前的状态,适应新的条件,或在不同的稳定状态之间交替[37]。
不幸的是,很少有受到严重开发的鱼类能够成功恢复的例子。例如,北大西洋箭鱼的恢复是成功的,因为其生物量并没有耗竭(种群数量达到最大可持续产量的70%),恢复时间相对较短(10年),捕鱼船队在管理方面进行了合作,这一合作管理是基于相似性和共同的激励措施而简化的成就[16]。否则,仅有有限的证据能说明长寿、成熟缓慢的顶级捕食者能够恢复,物种的恢复同样取决于许多其它因素,包括生物和政策因素[16][55]。
2 顺捕对生态系统潜在的影响
在捕捞过程中,目标物种和非目标物种都有可能会在无意中被杀死、伤害或以其它方式丧失生存能力,却不会被当作渔业产品而保留。这一现象叫做顺捕。被渔民保留的非目标物种应视为捕捞。对于一些海龟(例如红海龟Caretta caretta),捕捞造成的死亡非常大一部分都来源于顺捕。从种群动态的角度来看,与顺捕相关的死亡率与目标物种的死亡率不同[25][62]。对于顺捕类群和目标物种来说,确定其对海洋生态系统可能产生的高层次影响的挑战是相似的,但有关顺捕類群移除量的可用信息却更少。物种的生活史特征可以比营养级更好地预测捕捞影响,当顺捕类群与目标捕捞物种相比生长周期较长,成熟较晚时,在达到目标种群的死亡率限制之前,顺捕类群的移除量可能已经超过死亡率限制,使得顺捕类群在即使管理良好的渔业中也可能无意中被过度捕捞[51][58][67]。
海鸟,海龟,海洋哺乳动物和软骨鱼类是长寿类群中最常见的顺捕类群[66][67]。软骨鱼类(鲨鱼、鳐鱼和银鲛)在顺捕中很常见,并且在许多渔场中被越来越多地保留下来。自1984年以来,报告中所呈现出的全球渔获量稳步增长,但该类群的实际总渔获量可能是报告中所呈现的渔获量的两倍,而且有证据表明,软骨鱼类的物种丰盛度在下降[8][83]。对澳大利亚的一种在虾渔场中被偶然捕获的软骨鳃类鱼的研究估计,软骨鳃类鱼占该渔场顺捕的65%[105]。
许多种类海龟的数量与历史上的数量相比明显减少,一些种类的海龟的数量还在继续下降。据估计,每年有20万只红海龟(C. caretta)和5万只棱皮龟(Dermochelys coracea)作为大洋中长线渔业被顺捕[67]。关于个体渔业中被顺捕的濒危海龟数量的信息相对有限,但这一数量却不可否认的仍在增加[69][91]。
最近估计,每年被渔业捕捞的海洋哺乳动物超过65万只[95]。小头鼠海豚(Phocoena sinus)正处于濒危状态[57]。墨西哥巴哈河豚估计的最低死亡率是每年39只,超过了最近估计的小头鼠海豚种群数量的17% [30]。赫克托海豚(Cephalorhynchus hectori)也受到顺捕的严重威胁,顺捕是导致地中海僧海豹(Monachus Monachus)和北大西洋露脊鲸(Eubalena glacialis)种群数量持续减少的一个因素[95]。
热带太平洋东部围网捕捞金枪鱼的顺捕显著减少了东北近海斑点海豚(Stenella attenuata attenuata)和东方旋臂海豚(Stenella longirostris orientalis)的种群数量。虽然它们在这一渔业中的年死亡率下降了两个数量级,但它们的原有种群数量仍未恢复。尚未恢复的可能的假设包括(a)海豚的实际顺捕数量高于报告;(b)金枪鱼渔业的影响不限于造成顺捕类群的直接死亡;(c)制度的改变降低了海豚的生境;(d)恢复的期望不切实际。最后一个假设反映了没有考虑生态系统影响的特殊模式的不足,包括金枪鱼种群大规模迁移的未知影响[44]。
受顺捕威胁最大的海鸟包括信天翁、海燕和海鸥[106]。信天翁主要是在长线渔场中被捕获的,而海鸥和海燕最常在刺网中被捕获。每年有超过44,000只信天翁在南冰洋被日本人用长线捕获[13]。对于种群生存能力的分析表明,秘鲁的小型漂网渔业能够偶然捕获洪堡企鹅(humheniscus humboldti),但这一趋势是不可持续的[70]。
顺捕鱼类包括重要的经济鱼类的不可出售的个体(幼鱼、禁止出售的大小/长度类别)和不需要的鱼类,从而影响所有鱼类种群。最新估计全球丢弃的顺捕渔获量为730萬吨,而目标类群的捕获量为7800万吨[62]。虽然没有直接与早先的全球估计数值相比较,修改后的估计值比之前评估的2700万吨少[1]。对于被报道的长须鲸顺捕渔获量的下降的解释包括(a)越来越多的曾被当作顺捕渔获物所丢弃的物种如今作为捕获物而保留(b)有效地在渔具上使用避免顺捕现象的装置或有效应用其它捕鱼方法(c)随着海洋生态保护问题的增加使得相关部门减少了关于顺捕渔获量的报告。
这种对鱼类非目标渔获量的全球估计数据却也掩盖了区域和渔业特有的巨大差异。捕捞虾和底足鳍鱼的拖网渔船的丢弃量占估计的丢弃总量的50%以上,但只占记录捕获总量的22%[62]。其中,热带捕虾拖网渔船的弃虾率最高(达27%)。估计认为废弃生物量最多的地区是太平洋东北部和大西洋西北部,占全球估计总数的40%。据估计,北海废弃鱼类每年的废弃物和内脏总量分别为:内脏7万吨,底层鱼12万吨,比目鱼20万吨,底栖无脊椎动物18万吨[106]。美国渔业的丢弃率处于较高水平[49]。在美国,丢弃物中有86%以上是甲壳类和底栖类。虾类拖网捕捞和底栖类渔业产生的丢弃物占总废弃物重量的75%。
丢弃的非目标渔获物和动物内脏是对海洋环境的重要营养投入。生物废弃物通过向各种食腐动物提供营养补充,并引发物理化学变化,从而改变群落的组成和分布模式,并可以产生整个生态系统的变化。在某些情况下,丢弃物被认为是有益的:例如墨西哥湾虾渔业和大西洋中部各州的蓝蟹渔业。但微生物在未被利用的丢弃物上滋生,其对物质的分解会加剧这些地区水域的低氧环境。
能源补贴对食腐动物的生态影响受到越来越多的关注。生物废弃物的主要消费者包括底栖无脊椎动物、硬骨鱼、鲨鱼、海鸟和一些海豚(delphinids)。废弃物和内脏是具有觅食行为的海鸟的食物来源,包括塘鹅、臭鸥、海鸥和三趾鸥[17]。许多大型的海洋脊椎动物(海豚、海龟和鲨鱼)都喜欢捕食生物废弃物和在捕捞过程中逃跑的鱼。如果受影响的物种是强相互作用体,那么这种营养投入的影响可能是深远的。总的来说,生物废弃物可以通过抑制自上而下或自下而上的控制在食物网中的传播影响,间接地稳定生态系统[19]。但大多数处于濒危的顺捕类群都是顶级捕食者,因此,生态系统效应的例子,例如中级捕食者释放(例如,增加的牛鼻魟的丰盛度和减少的海湾扇贝收获)或生境退化(例如,食藻类动物的顺捕与珊瑚消费者的捕食性释放相结合)可能更频繁[9][82]。
3 商业捕捞对食物网的影响
商业捕捞对海洋生态系统的最大影响是使用与海底接触的移动渔具,它们能够同时捕捉目标和非目标物种,破坏底栖生物的生境。海底拖网包括水獭拖网和吊梁拖网,其主要目标是底栖鱼虾,还有从表层沉积物中清除扇贝、牡蛎、螃蟹和其它动物的设备[118]。固定在设备底部装置,如诱捕器、水罐、底部刺网和底部长线,通过挤压接触底部的生物和在浪潮中拖动来破坏底栖生物的生境[22]。深海渔具,包括中层拖网、深海长线网和围网,主要通过从食物网中清除目标和非目标物种来影响生态系统[22]。
生境的破坏、隐蔽场所丢失以及随后存活个体的减少是深海捕捞对海洋生态系统的基本影响。深海渔具减少了为目标和非目标物种提供庇护的硬底和软底生境的异质性和结构复杂性[110]。由于此类物种通常生长缓慢,寿命较长,形成群落结构的物种数量较少,渔民进入新水域后需要较长的恢复时间[60][110]。作为受商品渔业影响的生态系统的例子,软沉积物生境经常被忽视,但这些生境具有丰富的生物多样性,并覆盖了超过70%的海底,因此有必要考虑渔业对这些地区的生态系统影响[107]。墨西哥湾每一平方米的海底每年被拖网捕捞七次,这种扰动导致群落中的物种逐渐演变为快速移殖者[118]。
食物网的改变具有关联效应,影响其它食物网的组成部分,并可能导致海洋生态系统的稳态转换。食植性和捕食物种的消失导致了从珊瑚礁到开阔海域的生态系统的稳态转换[41]。在过去的100年里,北大西洋西部的缅因湾经历了两次由渔业引起的稳态转换[103]。在20世纪80年代末和90年代初,鳕鱼和其它主要底栖鱼类物种的灭绝与大型无脊椎动物和浮游植物的增加相关联,但大型浮游动物的数量却在减少,形成了四步营养级联和高度改变的生态系统[42]。虾蟹渔业取代了底栖鱼渔业并已经获得了较高的市场价值,但是以生态破坏为代价的低营养级渔业的发展是不可取的[42]。与Frank等人提出的营养级关联理论相反,人们认为,北极海冰融化导致的缅因湾盐度变化导致了浮游植物和浮游动物的自下而上的系统性变化[42][46]。由于气候变化与人为因素对海洋生态系统的作用是一致的,因此很难将因果关系准确分配给稳态转换的驱动因素。
在稳态转换和气候变化也可能改变海洋生态系统的情况下,很难分辨出商业捕捞的影响。自20世纪70年代以来,阿拉斯加西部海湾的Steller海狮数量减少了80%。对这种下降原因的假设包括营养压力、疾病、与商业目标物种的竞争以及虎鲸的猎物转换[34][83]。虽然这些过程可能部分或全部影响了种群的动态变化,但单个的假设并不能对观察到的种群变化趋势提供令人满意的解释[108]。历史上的种群波动的考古证据和生态系统的建模研究都表明,Steller海狮数量减少的最终成因可能是气候引起的全流域生态系统的变化 [83][108]。这个例子强调了在管理目标物种和非目标物种时,考虑多种相互作用的假设和潜在的复合效应的重要性。
由于大陆架渔业已被充分甚至过度开发,为了维持捕获量和利润,渔业已转向较深的水域。由于深海和海山生态系统生产力低下,这种转变可能无法持续。这些生态系统是值得注意的,因为其优势物种和目标物种都是非常长寿和生长缓慢的[36][64]。特别是海山生态系统,它们往往是孤立的,并有许多本地特有的物种,这些物种在很短的时间内遭受了严重的种群密度的下降[64]。根据国际自然保护联盟(IUCN)的标准,西北大西洋的五个深海渔场在17年的时间里总共下降了87-98%,这就足以将它们列为极度濒危物种[36]。
衡量渔业对海洋生态系统的影响是一项艰巨的任务,因为同时有许多非生物、生态和人为因素的协同作用。诸如生态路径/生态模拟系统等海洋食物网模型的发展使渔业生态学家能够评估对海洋生态系统的人为影响,这些影响太大、太复杂,无法进行实验研究[17]。物质平衡模型可以模拟各种渔业管理方法的潜在生态系统影响以及模拟复杂的食物网和空间相互作用效应[116][117]。气候驱动因素的可变性可以沿着食物网进行传播,影响北太平洋更高的营养级,尽管解释已预测的生物对气候驱动因子的反应可能很困难[78]。
最大可持续产量的单物种水平收获政策的广泛使用可能导致顶级捕食者的消失和生态系统功能⑧的广泛丧失[117]。其它模型表明,在远洋大型物种海洋保护区的设计中,海洋过程的可变性比分散性更重要[73],在太平洋长线渔业中,改变捕捞深度和使用圆钩代替长线可能比努力减少对旗鱼和鲨鱼种群的保护更有效[61]。
4 休闲渔业对食物网影响
休闲渔业是一个多样化且利润丰厚的行业,估计占全球渔获量的12%[26]。在世界范围内,约11.5%的人口参与休闲渔业,每年捕捞超过1000万吨的鱼类[26]。这种渔业的特点是为了休闲而捕鱼,而不是为了生存或利润,休闲渔业包括传统的钩线钓鱼,以及诱捕,排钩(渔叉)捕鱼,应用网、矛捕鱼和潜水捕鱼。休闲渔业的参与率因国家而异,留作消费的渔获物的百分比也大不相同[26]。虽然一个休闲垂钓者的行为可能看起来微不足道,但是数百万休闲垂钓者的累积作用可能对淡水和海洋环境产生深远的影响[23]。虽然休闲垂钓者仅占美国总捕鱼人员的一小部分(在2002年为4%),但休闲垂钓者收获了过高比例来自濒危物种的海产[23]。休闲渔业的收获在一些地区超过了商业捕捞收获,特别是在大城市和热门旅游目的地附近[77]。
在种群水平上,休闲渔业的潜在生态影响可与商业捕捞的潜在生态影响相比较,包括局部耗竭、增长和过度捕捞,生物量减少,制约机制的破坏,大小和年龄结构分布。垂钓者有时会把精力集中在更容易到达的近岸生境,所以有时候会捕获未成熟个体[26][77]。但许多垂钓者的目标“战利品”通常是体型更大、年龄更大的鱼,即从种群中剔除了繁殖能力最强的个体[11]。
休闲渔业的管理者采用许多同样适用于商业捕捞的管理策略,包括大小限制、渔具限制、占有限制和区域封锁。放生并避免伤害不能被合法饲养的捕获物,被认为是一种捕获-放归(C&R)捕鱼模式。因此,C&R捕鱼模式的广义定义包括任何涉及自愿或受到监管而放归所捕鱼类的渔业活动。虽然放归比例因物种而异,但目前估计释放了60%因休闲目的捕获的鱼类,高于1981年的34%[5][27]。
休闲渔业和商业捕捞从根本上是不同的,面临着不同的管理挑战。与大多数商品渔业不同的是,休闲渔业仍然是开放的,尽管美国大多數州颁发了咸水区域捕鱼许可证,但通常没有限制总捕捞量的机制,因此很难监管或减少捕捞量[77]。在许多地区,休闲渔业缺乏关于人力物力、捕获量、生物丢弃量和支出的一致的数据收集[77]。在美国,国家海洋渔业局目前使用电话和拦截调查来收集数据。但是垂钓者人数众多、分布广泛,这给取样带来了挑战,使得管理者很难实时监控休闲渔业趋势。
休闲垂钓者的动机与商业捕鱼者完全不同。管理者们常常错误地认为休闲渔业是能够自我调节的,如果钓鱼体验的质量下降,垂钓者就会退出。但是,如果垂钓者愿意容忍较低的捕鱼量,休闲垂钓者更有可能将渔业开发到崩溃的地步。因为休闲渔业的目标是一个小范围的营养级,所以与选择性较低的渔业相比,休闲渔业可能有更高的食物网效应[43][104]。
5 个体、小规模渔业对食物网的影响
联合国粮食及农业组织(粮农组织)的术语表将个体渔场定义为包括渔民家庭(而不是商业公司)在内的传统渔业,它们消耗相对较少的资金和能源,使用相对较小的渔船(如果有的话),在近海进行短途捕鱼航行,捕获的海产主要供给当地消费。在实际中,不同国家的定义是不同的,例如,有从贫穷发展中国家的收集船或单人独木舟到发达国家的20米以上的拖网渔船、围网渔船或远洋渔船。个体渔业可以是维持个体生计的渔业,也可以是供当地消费或出口的商品渔业。它们有时被称为小型渔业[39]。
即使是在考虑到渔民分类的差异之后,个体渔业在很大程度上超过了规模渔业。据估计,全世界5100万渔民中有99%是个体渔民[10]。
对渔民和捕鱼作业进行分类是困难的,个体和小规模(一个更为确切的描述词)的交替使用可能会增加评估这些渔业的影响的困难。首先,多概念的使用在各种局部条件下,会使数据收集和分析变得复杂和混乱。第二,由于个体渔业与其它经济活动(包括商业捕捞、航运、旅游和采矿)同时发生,很难将这些渔业在近岸海洋环境中的具体影响隔离开来,所有这些活动都会影响水质、幼体成长的生境和污染负荷,并会改变邻近的海洋生态系统的组成和功能[45]。在某些情况下,商业活动和个体活动的影响不是独立的。如果个体渔船队和工业渔船队都以单一鱼种为目标,如墨西哥湾的红石斑鱼渔场,则可以扩大商业渔船队的效果[15]。热带太平洋的长嘴鱼渔场与以这些洄游鱼类为目标的商业、休闲和个体渔场有类似的重叠[112]。
某些渔业(刺网捕捞、诱捕、人工捕捞)几乎完全由个体和小规模渔民进行,并在浅水海洋系统中占主导地位。这些相对非选择性的渔具能够捕获多物种,包括非目标物种的幼体[2]。副渔获物的保留是这些热带作业经济可行性的重要组成部分,因此这些渔场的生物丢弃率通常很低(例如,2006年报告的生物丢弃率为6.5%)[72]。
禁渔区或海洋保护区对个体渔业的影响提供了一些见解。关于个体渔业对群落聚集和食物网关系的影响的大部分信息是从热带系统收集的,包括印度洋和加勒比海[50][59]。Miller等人提供了一个加勒比纳瓦沙岛(Navassa Island)周围珊瑚礁生态系统的案例研究[81]。纳瓦沙岛相对独立于其它人为影响,这为研究海地本土渔民不断升级的个体渔业的影响提供了机会。礁鱼群落结构快速发生变化,主要表现为鱼类数量的减少。对厄立特里亚(Eritrea)渔获量的分析和研究也得到了类似现象的证据[109]。Tsehaye等人的结论是:“与珊瑚礁相关的底栖鱼类极易被捕捞,甚至在捕捞的早期就会减少[109]。”Hawkins和Roberts在比较加勒比珊瑚礁在一系列捕捞压力下的情况时指出:在主要食植动物(如海胆)消失后,在密集捕捞区域,食植性鱼类控制藻类过度生长的能力降低[50]。在加拉帕戈斯群岛(Galapagos Islands)也观察到了类似的关联效应、生态系统水平的影响,群岛中高度捕捞的区域有更多的海胆和结壳藻类[101]。Inhaca Island,Mozambique潮间带滩地的严重捕捞区表现出较低的捕捞率、较低的鱼类比例、较小鱼类比例的增加和物种多样性的减少[33]。
大量小规模渔民的影响可能是迅速的,其规模与商业捕捞的影响相似。在墨西哥南部下加利福尼亚州的两个小渔场中捕获的红海龟(C. caretta)的副渔获量相当于北太平洋整个工业长线钓船队的捕鱼量[91]。如果不停止或减少在刺网捕捞中意外死亡的海豚数量,那么位于大西洋西南部的Franciscana dolphin (Pontoporia blainvillei)的数量预计将在30年内大幅减少[63]。
尽管个体渔业通常不进行破坏或改变生境的作业,但它们会破坏形成群落的关键种。在肯尼亚南部的多渔具渔业中,使用海滩围网、长矛和刺网捕鱼的渔民对珊瑚造成了最直接的物理破坏[72]。
小规模渔业的异质性和复杂性为评估其影响并将其直接与大型商业捕捞作业进行对比提出了挑战[111]。因此,可以证明减少个体渔业的影响是具有挑战性的,并且可能是区域性的和/或依赖于特定渔具的[111]。小规模渔业明显改变了群落结构和群落内部的营养关系,但总体方向和过程与渔业的其它部分并无不同。个体渔业在以下方面可能有些独特:(a)近岸区的优势;(b)渔获物的多样性和低生物丢弃率;(c)对其它海洋动物的补贴较低。更好地管理采后损失可以减少需求和随后的清除率[39]。如果个体渔业与海龟或海洋哺乳动物等本质上易受影响的类群重叠,大量小规模渔民可以与商业捕捞的影响相媲美,并可能造成海洋生物类群的迅速减少。
6 人类活动对海洋食物网的累积影响
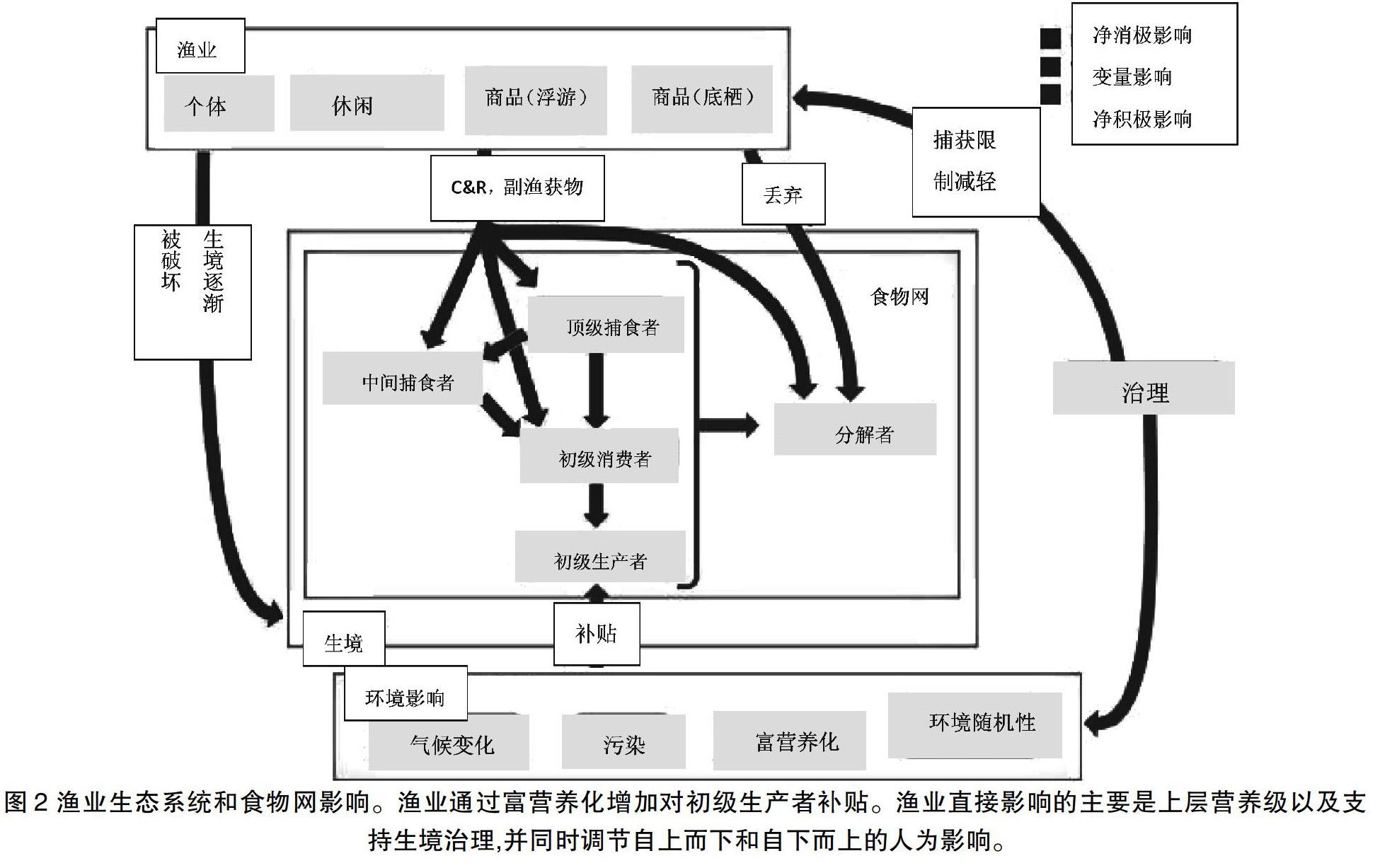
大多数海洋系统都能适应自然水平的干扰,但海洋生态系统受到自然和人类活动因素的影响不断增加、累积和协同作用会威胁海洋的恢复能力(图2)[47]。由于距人类活动近并处于下游,以及影响时间长,沿海食物网面临多种因素的影响[68]。虽然过去的研究侧重于商业捕捞对生态系统的影响,但副渔获物和个体与休闲渔业的额外影响可能进一步加剧食物网的退化,特别是在这些影响相互作用的地方。2006年,美国的休閑渔业主要集中在对顶级捕食者的捕捞,占所有捕获量的2.2%;被捕捞后又被放归的鱼的数量甚至更多(表2)。个体渔业也比规模渔业发挥了更大的作用。在印度洋,2006年个体渔场的捕鱼量比商业渔场的捕鱼量多20万吨(表3)。与大型商业渔场相比,个体渔场的捕鱼量是前者的4倍多,参与者是后者的2.5倍多。虽然大量的参与者和船只使得监测更加困难,但它们对生态系统的影响不容忽视。
![]()
![]()

![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()



![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
漁业通过富营养化增加对初级生产者补贴。渔业直接影响的主要是上层营养级以及支持生境治理,并同时调节自上而下和自下而上的人为影响。
由于牡蛎(Crassostrea virginica)产量的增加、富营养化水平高、水体大范围的缺氧以及水温的升高的协同作用,切萨皮克湾(Chesapeake Bay)等生态系统面临大规模崩溃[56]。这些条件导致有毒的鞭毛藻大量繁殖和鱼类死亡。沿海地区留下了大量腐烂的鱼,食物网也受到了破坏。不幸的是,众多尝试恢复牡蛎数量或整治整体水质的行为都几乎无效,但条纹鲈鱼的成功恢复仍然为未来的努力提供了希望[12]。由于系统的多种输入(陆地、河流、海洋)和参与者(流域到河口),沿海系统将继续面临来自收获、营养负荷和全球变化的协同效应[68]。协同效应需要通过食物网和包括全球变化在内的其它变化驱动因素来处理整个生态系统的管理策略。全球海洋生态系统的衰退刺激了研究领域的发展,包括历史生态学和恢复理论[56][75]。海洋生态系统管理需要充分考虑到历史影响、时间空间尺度、环境、自然和人类活动的影响[99]。
7 向海洋生态系统水平管理发展的渔业
只有从对单一方面的渔业活动管理转向以海洋生态系统水平的管理才能处理好多样化的渔业问题 [29,80,92,113]。然而,实施海洋生态系统水平管理的实际框架还在形成[76]。海洋生态系统具有很高的多样性[29][76][93][114][120],社会、文化、经济和政治属性也是重要的因素。基于地点的管理和海洋空间规划(MSP)可以提供一些基础的数据资料。以地点为基础的海洋生态系统管理需要从最一般的层次开始,即以基于生态系统的管理概念为基础,建立一套综合的管理办法,这种方法优先考虑维持健康的、具有生物多样性的、多产的和弹性的生态系统。在规划中必须明确多个时空尺度,以确保管理单元覆盖相关的生态过程和最终利益相关者的使用;理想的海洋生态系统水平管理办法的关键是设计治理系统,使利益相关者(在本例中是渔民)的激励措施与管理目标保持一致[52]。充分整合基本生态系统模板并明确纳入社会经济和治理覆盖的管理服务方案,可以为充分保护海洋生态系统和合理利用包括渔业在内的海洋资源奠定基础。
参考文献
[1] Alverson DL,Freeberg MH,Pope JG,Murawski SA.1994.A global assessment of fisheries bycatch and discards[T].Rep.339,FAO Fish.Tech.Pap.,Rome.
[2] Ambrose EE,Solarin BB,Isebor CE,Williams AB.2005.Assessment of fish by-catch species from coastal artisanal shrimp beam trawl fisheries in Nigeria[J].Fish. Res. 71:125-32.
[3] Anderson TW.2001.Predator responses,prey refuges,and density-dependent mortality of a marine fish[J].Ecology 82:245-57.
[4] Bakun A,Broad K.2003.Environmental ‘loopholes and fish population dynamics: comparative pattern recognition with focus on El Nino effects in the Pacific[J]. Fish.Oceanogr.12:458-73.
[5] Bartholomew A,Bohnsack JA.2005.A review of catch-and-release angling mortality with implications for no-take reserves[J].Rev.Fish Biol.Fish.15:129-54.
[6] Bascompte J,Melian CJ,Sala E.2005.Interaction strength combinations and the overfishing of a marine food web[J].Proc.Natl.Acad.Sci.USA 102:5443-47.
[7] Baum JK, Myers RA.2004.Shifting baselines and the decline of pelagic sharks in the Gulf of Mexico[J].Ecol.Lett.7:135-45.
[8] Baum JK, Myers RA,Kehler DG, Worm B, Harley SJ,Doherty PA. 2003. Collapse and conservation of shark populations in the Northwest Atlantic[J].Science 299:389-92.
[9] Bellwood DR,Hughes TP,Folke C,Nystrom M.2004.Confronting the coral reef crisis[J].Nature 429:827-33.
[10] Berkes F,Mahon R,McConney P,Pollnac R,Pomeroy R.2001.Managing Small-Scale Fisheries: Alternative Directions and Methods[T].Ottawa:Int.Dev.Res.Cent.250 pp.
[11]-[121]文獻略
注释
① 商业捕捞:通常以大船、高渔获量和高出口量为特征。
② 非目标渔获物:非目标渔获而被捕捞,并且用于消费或销售。
③ 顺捕:非目标渔获而被捕捞、直接受伤或死亡的生物;可能会被捕捞上岸或者被丢弃。
④ 休闲渔业:无营业执照的垂钓者从事捕捞-放生或者捕捞-个人消费的活动;非生活必需品。
⑤ 个体渔业:主要是自给自足的渔业,从陆地出发进行短途航行,以捕捞为主,供当地消费;也称为小规模渔业。
⑥ 自上而下的控制:食物网的结构和种群数量的动态受到顶级捕食者的控制。
⑦ 自下向上控制:食物网的结构和种群数量的动态受到营养物质和初级生产力的调节。
⑧ 生态系统功能:生态过程,如迁移和捕食,包括多营养级的食物网,功能的双重性,高关联度。从人类的角度来看,生态系统提供的服务(海产品、旅游、交通)是可持续的。
收稿日期:2019-11-22
作者简介:黄大明(1963—),男,北京人,博士,特别研究员,研究方向:生态学。
The Impacts of Fisheries on Marine Ecosystems and the Transition to
Ecosystem-Based Management
HUANG Da-ming1SHAO Hai-pei1LI Ru-yi1Larry B. Crowder2
(1.Laboratory of ecology, School of life sciences, Tsinghua University, Beijing 100084, PRC; 2.Center for Marine Conservation, Duke University Marine Laboratory, Nicholas School of the Environment, Beaufort, North Carolina 28516, US)
Abstract: Fishing remains one of the largest factors modifying marine ecosystems. Because Fisheries constitute only one of many anthropogenic effects, management is shifting from single-species approaches toward ecosystem-based management. Interaction webs are a critical nexus to understand linkages, to model ecosystem change, and to apply management. Ecosystem-based management requires consideration of both direct and indirect effects of commercial Fisheries. But it must also include impacts of bycatch, recreational Fisheries, artisanal Fisheries, and environmental change that can be large but unanticipated. Synergistic effects of Fishing, environmental variation, and climate change increasingly threaten marine ecosystems and complicate management. Here we review the global effects of Fisheries and propose an integrated framework for managing biophysical processes and human ecology. To incorporate the multitude of effects, this emerging approach focuses on the dynamics of interaction webs in a spatially explicit or place-based framework.
KeyWords: artisanal Fishing;bycatch;commercial Fishing;direct and indirect effects; ecosystem-based management;food webs;habitat effects;interaction webs; recreational Fishin
