水-肥耦合对新银合欢幼苗抗氧化酶活性及肥力利用能力的影响
2019-05-14徐建霞王建柱
徐建霞,王建柱
(1.三峡大学 三峡地区生态保护与治理湖北省国际联合研究中心,湖北 宜昌 443002;2贵州省农业科学院旱粮研究所,贵州 贵阳 550006)
【研究意义】干旱是影响植物生长发育的逆境因子,对作物造成的损失仅次于病虫害危害[1]。氮和磷是合成蛋白质、核苷酸、磷脂和植物体内某些生长素的主要元素,与植物的生理代谢活动密切相关,在植物细胞的分裂分化、发育生殖及各器官的分化发育过程中发挥重要作用,是影响植物生长的重要营养因子[2]。当氮和磷元素成为植物生长发育的限制元素时,会影响植物的生长与光合[3]、叶片形态变化[4]、根系统形态及结构变化[5]、地上地下生物量分配及植物功能群落组成[6]、根的形成和寿命[7]。新银合欢(Leucaenaleucocephala)适生性强,主根极发达,为豆科(Leguminosae)银合欢属(Leucaena)的多年生常绿乔木,是三峡地区及我国西南山区理想的水土保持和木本绿肥作物,探讨不同水肥水平对新银合欢幼苗抗氧化酶活性及肥力利用能力的影响,对新银合欢幼苗栽培及管理具有重要意义。【前人研究进展】在水分不足的干旱或半干旱地区施用肥料是否有利于植物或农作物的生长发育一直是一个有争议的问题[8]。Li[9]等研究认为,在土壤水分有限的条件下,施用某些肥料有利于缓解或消除植物营养亏缺对生长发育产生的不利影响。刘锁云[10]的研究也认为,在相当宽裕水势范围内适当增施肥料能提高作物对干旱的忍受能力,且随着施肥浓度的加大,土壤肥力增加,作物能增加对养分的吸收,从而提高旱地作物产量。Xue[11]等研究发现,施用肥料能促进作物根系生长发育,使之在干旱条件下能加速生长并从土壤深层探寻和吸收水分,从而提高作物对水分的利用效率,提高产量。但也有学者认为,在土壤极度干旱的情况下,施肥对作物无效,在轻度干旱的条件下,施肥则有利于提高作物生长和产量[12]。吕丽华[13]等研究水肥互作对冬小麦及夏玉米的生长影响,结果发现限水条件下土壤水是限制氮肥肥效发挥的主要因素,通过改善土壤水分条件可更有效提高氮肥肥效;闫春娟[14]等对大豆的研究发现,水肥互作对大豆茎秆、荚皮及籽粒产量呈显著性的影响。而目前关于新银合欢的研究报道都主要是对单一环境因素(如水分、土壤质地、土壤养分)的响应[15-17],然而关于水分和施肥对新银合欢幼苗的联合影响尚不清楚。【本研究切入点】目前,针对不同水肥条件下抗氧化酶活性和肥力利用能力影响的研究主要集中在农作物和经济作物上,尚无水肥互作对新银合欢幼苗抗氧化酶活性及肥力利用效率的报道。【拟解决的关键问题】本文以三峡库区边坡消落带土壤为基土,采用室内控制法研究新银合欢幼苗在不同水肥条件下的抗氧化酶活性及肥力利用效率,探讨新银合欢苗木的抗旱机理;通过新银合欢幼苗对施肥量的响应,确定其在受到干旱胁迫时的施肥最佳值,为边坡生态修复树种新银合欢幼苗前期引种及栽培管理提供科学依据。
1 材料与方法
1.1 试验材料
实验材料为豆科(Leguminosae)银合欢属(Leucaena)新银合欢幼苗,由三峡地区生态保护与治理湖北省国际联合研究中心提供。供试土壤为三峡库区边坡消落带土壤,其基本理化性质:全氮1.041 g/kg,有机质14.82 g/kg,全磷0.896 g/kg,pH 7.3(水土比为1∶2.5)。供试土壤经充分混合,随机分装于花盆中。所有花盆均放在同一个温棚隔间。所施氮肥选用硫酸铵(含N 21 %),磷肥选用过磷酸钙(含P2O518 %)。
1.2 试验方法
1.2.1 试验设计 将新银合欢幼苗移栽至内径25 cm、深30 cm的塑料花盆中,每盆1株。待塑料花盆中的幼苗长高至约16 cm(6月4日)时,开始进行施用水肥试验。设正常浇水组(土壤含水量约维持在田间持水量的75 %)和干旱胁迫组(试验前1 d正常浇水,自试验开始至结束(7月24日)不再浇水(共进行干旱胁迫50 d);试验过程中进行施加氮肥干预,设4个施肥梯度处理:不施氮CK(N0)、低氮(N1)、中氮(N2)和高氮(N3),施氮比例为0∶1∶2∶3,其中,N1为0.86 g/株。每处理重复6株,随机区组排列。各氮肥干预处理所施磷肥量相同,均为0.3 g/株。各肥料施用量见表1。
1.2.2 测定指标及方法 叶片取样及抗旱性相关指标测定:叶片取样和抗旱性相关指标测定参照李合生[18]方法。其中,叶片电导率采用电解质渗量法测定;叶绿素含量采用丙酮浸提法测定;可溶性糖含量采用蒽酮法测定;游离氨基酸含量采用茚三酮溶液显色法测定;SOD活性采用氮蓝四唑(NBT)法测定;POD活性采用愈创木酚法测定;CAT活性采用紫外吸收法测定;丙二醛(MDA)含量采用硫代巴比妥酸比色法测定。
新银合欢幼苗生长势和生物量的测定:幼苗株高等指标采用米尺进行测量,并观察记录植株叶片生长状况和分枝等形态生长特征。幼苗生物量于7月24日试验结束时测定:将各盆植株与土壤分开后洗净根、茎和叶上的土壤,晾干,分类存放。根和茎剪切成小块后与叶各自分开置于烘箱中先于105 ℃杀青处理15 min,后置于65 ℃烘箱烘干24 h至恒重,取出冷却至室温后在分析天平(精确度为0.0001)上称其干重,三者重量之和即为新银合欢幼苗的总生物量。
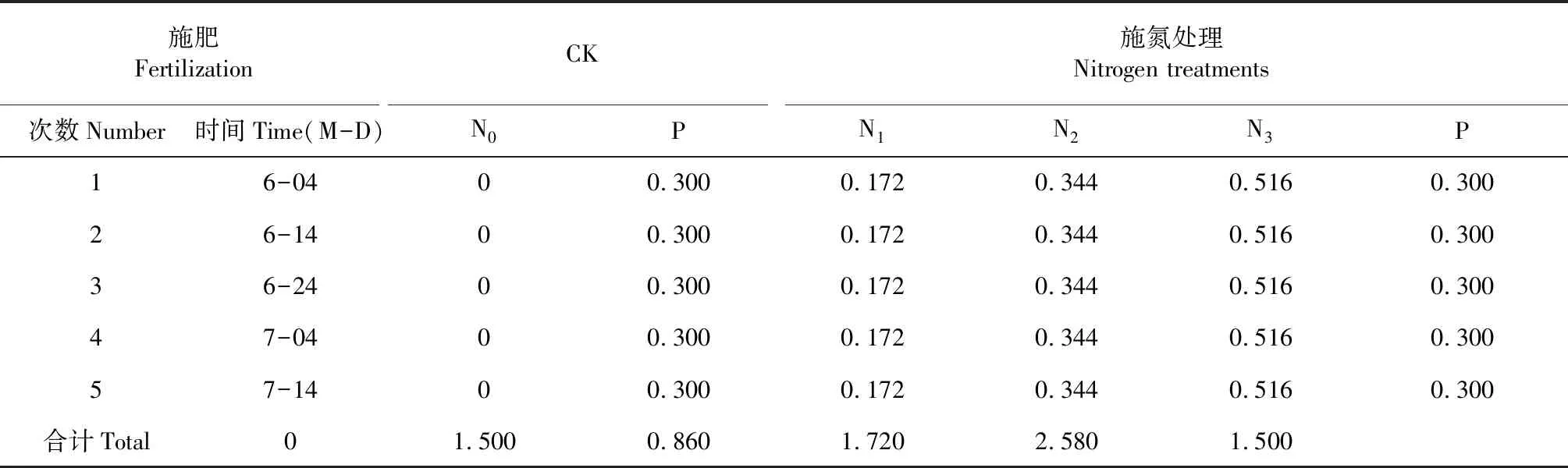
表1 新银合欢幼苗不同施肥方案(g/株)
新银合欢幼苗根、茎、叶各部分氮磷含量测定:植物氮、磷含量采用浓H2SO4-H2O2法消煮后,参照杨贵明[19]的方法进行测定;土壤氮、磷含量采用浓H2SO4-HCOl4法消煮后均采用连续流动分析仪进行测定。
相关参数的计算:氮(磷)运转效率(%)=叶片中氮(磷)积累量/整个植株(根、茎、叶)积累量×100;氮素肥料当年利用率(%)= [施氮处理整个植株(根、茎、叶)中氮素累积量-无氮处理整个植株(根、茎、叶)中氮素累积量]/氮素肥料施用量×100 %。
1.3 统计分析
试验数据采用Excel 2007进行整理,以SPSS 19.0进行统计分析,以LSD进行差异显著性检验和多重比较,以Origin 8.1和Excel 2007制图。
2 结果与分析
2.1 不同处理植株幼苗生物量含量的变化
正常浇水时N1、N2和N3处理的单株总生物量和各营养器官生物量均显著高于CK(N0)(P<0.05,下同),N1、N2和N3处理间也存在显著差异(表2),根占比重以CK(N0)最高,N2处理最低;茎和叶占比重以N2处理最高。干旱胁迫时,N2和N3处理的单株总生物量及各营养器官生物量均显著高于CK(N0)和N1处理,而N1处理与CK(N0)无显著差异(P>0.05,下同);根占比重以CK(N0)最高,随施氮量的增加,根占比重呈下降趋势;茎占比重以N2处理最高,叶占比重则以N1处理最高。随施氮量的增加,干旱胁迫和正常浇水条件下新银合欢幼苗生物量积累量的差值在减少,即干旱胁迫组N1处理幼苗的生物量减少量(1.09 g)大于N2处理(0.53 g)。

表2 不同土壤水分和施肥条件对新银合欢幼苗生物量及其分配的影响
注:同一水分条件同列数据后不同小写字母表示差异显著(P<0.05)。
Note: The different lowercase letters on the same water condition represent significant difference (P<0.05).
2.2 不同处理对新银合欢幼苗叶片抗氧化酶活性的影响
2.2.1 对超氧化物歧化酶(SOD) 活性的影响 叶片是植物光合作用的主要器官,也是对环境变化最为敏感的器官之一,叶片中的酶活性是植物对环境的适应或环境变化的最直接响应[20]。超氧化物歧化酶(SOD)是植物体内重要的抗氧化酶,可以清除植物体内自由基并对抗与阻断因氧自由基对细胞造成的损害,是植物抗逆生理的重要指示。相同施氮条件下,干旱胁迫组幼苗叶内SOD总活性明显高于正常浇水组;其中干旱胁迫时N2处理SOD总活性最高,正常浇水CK(N0)处理SOD总活性最低(图1)。正常浇水时, SOD总活性随施氮量的增加而增加,CK(N0)处理SOD总活性显著低于N1、N2和N3处理,表明水分充足条件下增施氮可提高新银合欢幼苗叶SOD的总活性;干旱胁迫时,CK(N0)处理SOD总活性最低,N2处理最高,而N3处理时SOD总活性有所下降,CK(N0)组SOD总活性与N2、N3之间差异性显著而与N1不显著,这表明适宜的氮浓度能显著提高新银合欢幼苗叶SOD的总活性,缓解干旱胁迫导致的活性氧或其它过氧化物自由基对幼苗叶细胞膜系统的伤害,延缓叶片衰老。
2.2.2 对过氧化物酶(POD) 活性的影响 植物体中过氧化物酶(POD) 活性与呼吸作用、光合作用及生长素的氧化等均有关,也是植物组织老化的一种生理指标[20]。同一氮肥处理下,干旱胁迫组幼苗POD总活性显著高于正常浇水组,表明水分对POD活性有显著影响。干旱胁迫下,POD总活性随施氮水平升高呈先上升后降低的趋势,CK(N0)处理显著低于N2和N3处理;正常浇水时,POD总活性随施氮水平的升高而升高,N3处理与其它处理组间均存在显著性差异(图2)。干旱条件下N2(1.72 g/株)处理叶片POD总活性的升高量(与CK处理的差值73.1 U/gFW·min)高于正常浇水条件下N2处理的升高量(43.8 U/gFW·min),说明在新银合欢幼苗受干旱胁迫时,可以通过适量施氮(1.72 g/株)来减轻自由基对叶细胞的伤害,从而减轻干旱胁迫对幼苗叶片的影响(图2)。

同一处理图柱上不同小写字母表示差异显著(P<0.05);下同Different lowercase letters on the same column represented significant difference(P<0.05). The same as below图1 不同水肥条件下幼苗叶片SOD活性含量变化Fig.1 Changes of SOD activity of seedlings leaves under different water and fertilizer concentrations

图2 不同水肥条件下幼苗叶片POD活性含量变化Fig.2 Changes of SOD activity of seedlings leaves under different water and fertilizer concentrations
2.2.3 对过氧化氢酶(CAT) 活性的影响 过氧化氢酶(CAT)是一种酶类清除剂,可促使H2O2分解为分子氧和水,清除体内的过氧化氢,从而使细胞免于遭受H2O2的毒害,是生物防御体系的关键酶之一。相同氮肥处理下,干旱胁迫组幼苗叶CAT总活性明显高于正常浇水组,说明水分缺失诱导了叶片内CAT的活性增强,在一定程度上减缓了干旱对幼苗叶片的伤害(图3)。正常浇水时,随氮素浓度的升高, CAT总活性含量呈上升趋势,N3与N2处理显著高于CK(N0)处理,而N3与N2处理间无明显差异;干旱胁迫下也呈现出相似的结果。水分充足和干旱胁迫下施氮均能提高新银合欢幼苗叶片CAT的总活性,有利于植物清除H2O2,使膜脂过氧化物维持在较低水平,实现自我保护。

图3 不同水肥条件下幼苗叶片CAT活性含量变化Fig.3 Changes of CAT activity of seedlings leaves under different water and fertilizer concentrations

表3 不同水分施肥条件下幼苗对氮磷的吸收情况
注:同一水分条件同列数据后不同小写字母表示差异显著(P<0.05);NC:含氮量;PC:含磷量。
Note: The different lowercase letters on the same water condition represented significant difference (P<0.05). NC:Nitrogen content; PC:Phosphorus content.
2.3 不同处理下幼苗对氮磷的吸收及转运
2.3.1 氮累积量 施氮相同时,正常浇水的氮积累量整体高于干旱胁迫组。正常浇水时,N3处理的氮累积量最高,且显著高于CK(N0)处理,但与N2处理差异不显著;干旱胁迫时,幼苗中氮累积量随施氮水平升高呈先增加后降低的趋势,在N2处理时最高(表3)。正常浇水条件N3处理(1.101 g/kg)、干旱胁迫条件N1(1.097 g/kg)和N3处理(1.203 g/kg)的土壤中氮累积量高于原始土壤氮累积量(1.041 g/kg),其他处理则均低于原始值。此外,从幼苗植株根、茎、叶氮素累积状况分析得出正常浇水和干旱处理组幼苗的氮累积量均表现为叶>茎>根的规律,叶的氮素占总氮素的50 %左右。
2.3.2 磷累积量 水氮互作下新银合欢幼苗中磷素累积量的变化为施氮相同时正常浇水组幼苗中磷积累量高于干旱胁迫组,与氮的总积累量变化趋势相同(表3)。正常浇水时幼苗体内磷积累量为N3处理最高,干旱胁迫时为N2处理最高;不同处理组的土壤磷积累量均小于原始土壤磷含量(0.896 g/kg)。不同器官的磷累积量在正常浇水和干旱处理下均表现出根>茎>叶的规律,且根累积量显著较高。

表4 新银合欢幼苗氮磷运转效率
2.3.3 幼苗氮、磷运转效率 不同水分和氮素条件下,氮、磷运转效率存在一定差异(表4)。正常浇水条件下N3处理氮运转效率最高为57.65 %,对照组CK(N0)最低为 45.63 %;干旱胁迫条件下氮运转效率在N2处理时最高为46.37 %,而对照组CK(N0)最低(40.47 %)。磷的转化效率在正常浇水条件下变化范围为24.76 %~29.66 %,其中,对照组CK(N0)磷转化效率最高;在干旱胁迫条件下磷的转化效率变化范围为 22.22 %~25.02 %,在N2处理时最高,CK(N0)处理时最低。
2.3.4 幼苗氮素利用率 不同水肥条件下,新银合欢幼苗氮素利用率存在较大差异。正常浇水时,N1处理氮素当年利用率最高为7.83 %;N3处理最低为6.19 %。干旱胁迫条件下氮素当年利用率在N2处理时最高为5.42 %;N1最低(2.32 %)。从根茎叶各器官的氮素当年利用率情况看,施氮处理后幼苗吸收的氮素都主要用于叶片产量的增加(表5)。
2.4 水肥互作对新银合欢幼苗各因素的影响
双因素方差分析表明水分与新银合欢幼苗总生物量,TN、可溶性糖、SOD、游离脯氨酸含量、丙二醛(MOD)、POD、CAT均存在极显著的正相关性(P<0.01),与叶绿素、TP和电导率呈显著性正相关(P<0.05)。而氮肥的用量除与幼苗TP含量无显著性相关外(P>0.05),与其它指标均呈显著性正相关(P<0.05)。而水分和氮肥的互作效应与所有指标都呈极显著正相关(P<0.01),这说明水肥的耦合效应对新银合欢幼苗在干旱条件下的生长和抗旱具有显著的作用(表6)。
2.5 不同水肥条件下幼苗各指标之间的相关性分析
由表7可知,生物量除与电导率不相关与MDA显著负相关(P<0.05)外,与其它指标之间都呈极显著正相关(P<0.01);全磷与全氮、叶绿素、游离脯氨酸显著相关(P<0.05),而与其他指标不相关(P>0.05);全氮除与电导率和丙二醛呈显著负相关外,与其它指标呈极显著正相关(P<0.01)。SOD、POD、CAT 3种酶呈极显著正相关(P<0.01);POD和CAT都分别与MDA呈极显著负相关(P<0.01),而SOD与MDA呈显著负相关(P<0.05)。

表5 新银合欢幼苗氮素利用率
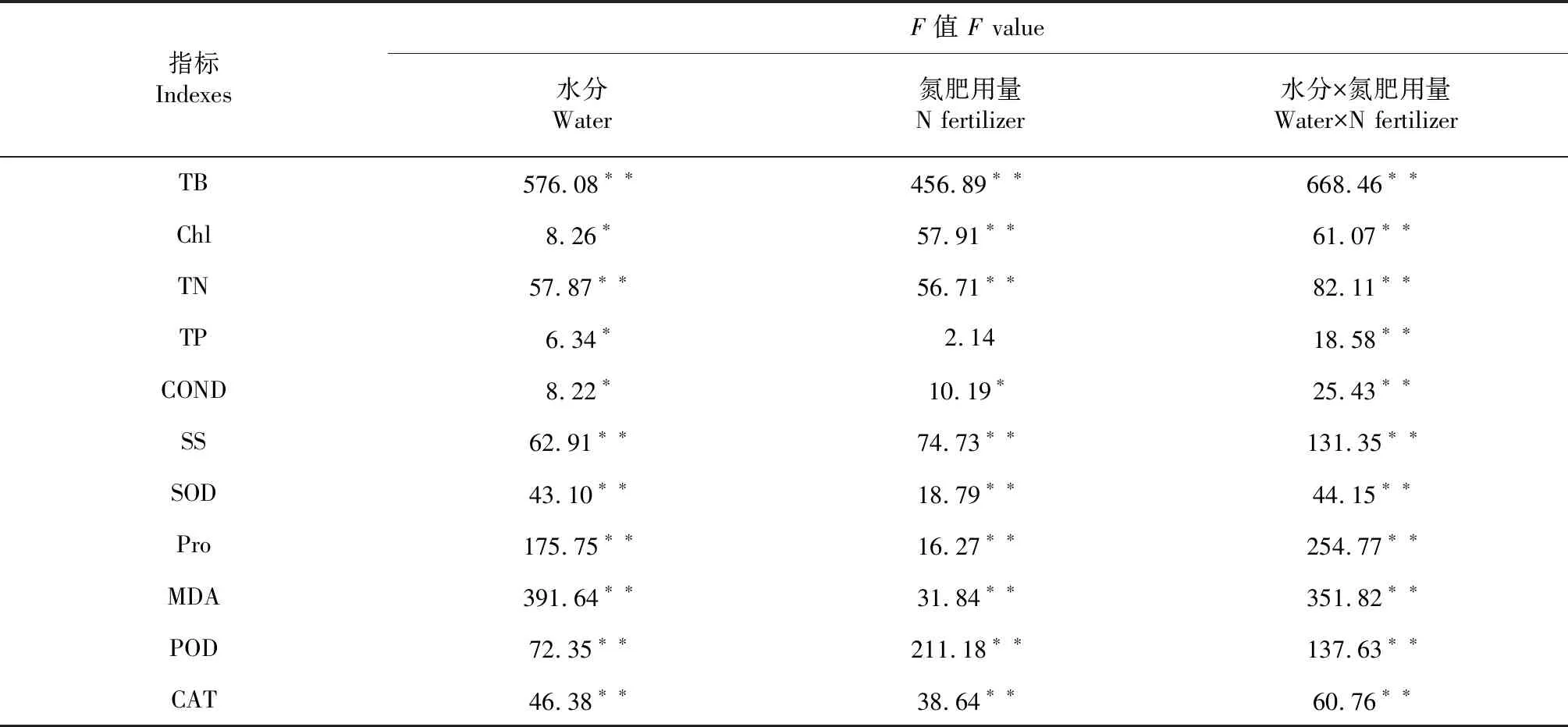
表6 水肥对幼苗各指标之间的影响
注:*表示显著水平(P<0.05);**表示极显著水平(P<0.01)。TB:总生物量;Chl:叶绿素;TN:全氮;TP:全磷;COND:电导率;SS:可溶性糖;SOD:超氧化物歧化酶;Pro:游离脯氨酸;MDA:丙二醛;POD:过氧化物酶;CAT:过氧化氢酶。下同。
Note: * represented significant difference (P<0.05); **represented extremely significant difference (P<0.01). TB: Total biomass; Chl: Chlorophyll;TN: Total nitrogen; TP: Total phosphorus; COND: Conductivity; SS: Soluble sugar; SOD: Superoxide dismutase; Pro: Free proline; MDA: Malondialdehyde;POD: Peroxidase; CAT: Catalase.The same as below.
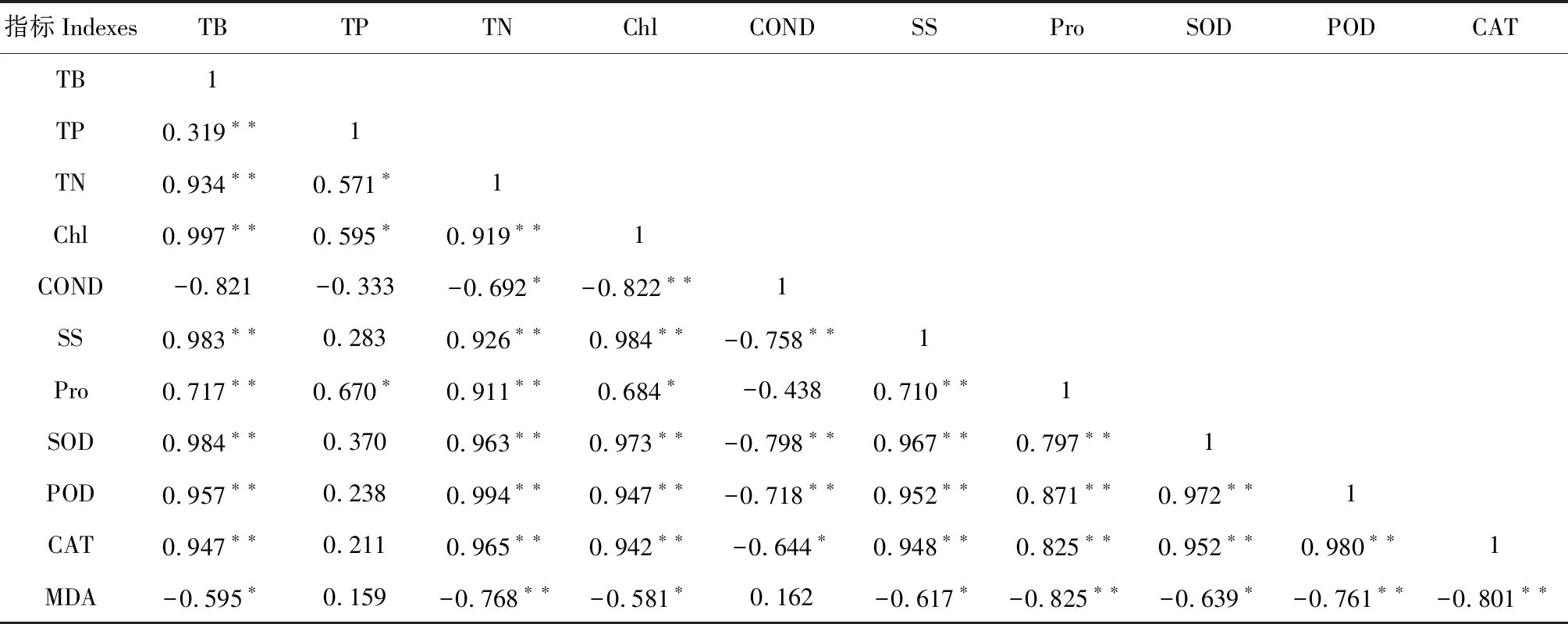
表7 新银合欢幼苗各指标之间的相关性
3 讨 论
3.1 水肥互作对抗氧化酶活性的影响

本研究中水分×氮肥的互作效应与SOD、POD、CAT 3种酶都呈极显著正相关(P<0.01),说明水氮的耦合效应对新银合欢幼苗在干旱条件下的生长和抗旱具有显著的作用;有研究表明,当植物遭受逆境胁迫(干旱、冻害、高温等)时,单一抗氧化酶或抗氧化剂不能抵御植物在其环境下受到的伤害,许多试验和研究发现在逆境条件下植物受伤害程度与SOD、POD和CAT等抗氧化酶活性提升成负相关[22]。本实验中SOD、POD和CAT活性之间呈显著正相关,表明3种抗氧化酶起着相互协调的作用。这与杨青川[26]等对扁蓿豆、紫花苜蓿的研究和王小乐[27]等对甘蔗的研究结果一致,说明这些抗氧化酶在新银合欢幼苗叶内可协同清除叶内多余的活性氧自由基,降低叶细胞渗透性的增加,减少离子大量泄漏及膜脂过氧化,从而保护了叶生物膜的结构和功能,降低或消除干旱胁迫对幼苗叶内细胞造成的伤害[23]。
3.2 水肥互作对幼苗氮磷吸收的影响
土壤中的水分和营养物质是影响植物各阶段生长发育和生理代谢的重要影响因子,两者之间有着密切的联系,研究表明土壤干旱制约着植物对氮磷等营养元素的吸收利用,而营养元素,特别是氮素不足又会极大的限制水分的作用[28]。本研究发现,施氮量相同时,正常浇水能促进新银合欢幼苗根系以上各形态指标上升,在干旱胁迫时适当施氮也能促进幼苗生长和总生物量及各器官生物量的提高,这与周利平[29]等的研究结果相同。适量的施氮可以减缓水分胁迫引起的氮代谢紊乱,使缺水植株的蛋白酶、肽酶及核糖核酸酶活性增强,从而维持较高的硝酸还原酶(NR)活性和蛋白质含量[30],有利于植物根、茎、叶等营养器官的发育和植株群体的协调发展,增强光合作用,因而有利于干物质的生产与积累[31]。本研究中土壤水分和氮肥之间存在明显的交互效应,当土壤含水量较低时,植物对氮的吸收也相对较低。这主要是由于干旱抑制了植物根系的生长,降低了根系的吸收面积和吸收能力,木质部液流粘滞性增大,从而降低了对营养物质的吸收和运输[32]。
本研究中施氮显著提高了新银合欢幼苗根、茎、叶的氮磷含量,且氮的分配排序为叶>茎>根,磷的排序为根>茎>叶,幼苗的氮养分主要储存在叶片中,而磷主要储存在根系中,这与杨贵明[33]等对桑树的研究结果相一致,也进一步表明叶是新银合欢幼苗重要的氮养分贮存库。正常浇水条件下新银合欢幼苗氮转运效率随着施氮量的增加而升高,而氮利用效率的变化趋势恰好相反。低氮条件下,氮的转运效率低而利用效率高;原因是低氮条件下少量的氮养分主要贮藏在根系中供根系生长,而向地上部的运输量减少,这与张耀华[34]等对橡胶苗研究结果一致。高氮条件下,根系吸收氮的同时向地上部运输量增加[34],因而氮的转运效率高而利用效率低。在干旱胁迫条件下,新银合欢幼苗氮、磷运转效率及氮素当年利用率均在N2(中氮1.72g/株)水平时最高,而超过此水平后其转运效率和利用率均下降,原因可能是适宜浓度的施氮提高了土壤水势和水分的有效性,使一部分原来对植物生长“无效”的水变得“有效”,而水分的有效性又会促进根系对营养物质的吸收和转移[35];而过量施用氮肥,反而会加重植物水分胁迫的程度,造成植物体内营养物质的亏损和作物减产[36]。
4 结 论
在水分充足条件下,增施氮肥可提高新银合欢幼苗的抗氧化酶活性和对氮、磷的吸收能力;干旱胁迫条件下施用中等浓度氮(1.72 g/株)可有效诱导新银合欢幼苗抗氧化保护酶系统活性增强,从而增强其抗旱性,延缓叶片衰老,同时可显著增加幼苗对氮磷营养物质的吸收,提高氮素的转运及利用效率。
