黄河三角洲滩涂—湿地—旱地土壤团聚体有机质组分变化规律*
2019-04-25刘兴华章海波代振飞付传城骆永明
刘兴华 章海波 李 远 代振飞 付传城 骆永明,4†
(1 中国科学院海岸带环境过程与生态修复重点实验室(烟台海岸带研究所),山东烟台 264003)(2 中国科学院大学,北京 100049)(3 浙江农林大学环境与资源学院,土壤污染与生物修复重点实验室,浙江临安 311300)(4 中国科学院土壤环境与污染修复重点实验室(南京土壤研究所),南京 210008)
全球土壤碳库储量约为2 500 Pg,是大气碳库的3倍,同时也是陆地生态系统最大且周转最慢的碳库[1]。目前,对内陆地区土壤,如森林土壤[2]、草地土壤[3]、耕地黑土[4]、耕地红壤[5]、湿地土壤[6]、潮滩盐碱土[7]等土壤有机碳研究已有较多报道。海岸带处于海陆交错地带,在全球碳循环中占有重要地位。黄河三角洲是世界河流三角洲中海陆变迁最活跃的地区之一,也是我国造陆速度最快的河口三角洲[8]。该区域同时受到黄河泥沙冲积、海相沉积等自然过程影响,以及滩涂盐渍土改良、水产养殖、油田开发等人类活动的影响,由海向陆依次形成了无植被滩涂、盐生植被湿地和农用地等系列土地利用类型[9]。滩涂土壤受海水浸渍影响,土壤盐分含量高、有机质含量低、土体发育不明显;随着耐盐植被生长,滨海湿地土壤有机质累积明显,一些土壤表层甚至形成泥炭层[9]。修建海堤围垦的农用地土壤,经过长期耕作洗盐,土壤盐分大大降低,逐渐向潮土演化,同时受耕作活动的影响,土壤有机质积累速度和周转过程与湿地明显不同[10]。目前对海岸带从滩涂到农用地演变过程中土壤有机质组成尤其是颗粒态有机质的变化及土壤有机碳库周转影响的了解还很少。
土壤有机碳库由活性有机碳库和惰性有机碳库组成[11-12]。Cambardella和Elliott[13]通过湿筛分组的方法将得到的粒径大于53 μm的有机质组分定义为颗粒态有机质(Particulate Organic Matter,POM),归属活性碳库。结合在土壤团聚体表面的半分解动植物残体、真菌菌丝体、孢子和粪球等有机质称为游离颗粒态有机质(Free Particulate Organic Matter,fPOM),而存在于土壤团聚体内部的称为团聚体内部结合的颗粒态有机质(Intraaggregate Particulate Organic Matter,iPOM)[14]。因受到土壤团聚体的保护程度不同,在相同土壤环境条件下iPOM也呈现出与fPOM不同的组成比例和稳定性[15]。与颗粒态有机质不同,矿物结合态土壤有机质(Mineral-associated Soil Organic Matter,mSOM)粒径通常小于53 μm,多由腐殖化程度较高的有机物质组成,同时受到土壤黏粒和粉粒的物理化学作用保护,属于土壤有机碳的“惰性库”[16]。通过湿筛法和密度分离法得到的这些土壤有机质组分,基本上能够反映原状土壤中有机碳的存在形态和特征,为认识土壤有机碳周转特征奠定了基础[17]。同时,不同有机质组分稳定性碳同位素特征也存在差异,利用碳稳定同位素分馏特征可以更准确地分析不同有机质组分中有机碳来源[18]、有机碳库的稳定性[5]和碳库周转[14]等特征。
因此,本研究旨在前期对黄河三角洲土壤碳氮调查工作的基础上[9],综合运用土壤有机质的物理分组方法和稳定性碳同位素技术,进一步分析黄河三角洲滨海滩涂无植被荒地—盐生植被湿地—旱地演变过程中土壤颗粒态有机质含量及其稳定同位素的变化特征,探讨滨海地区不同土地利用方式下土壤有机碳的稳定性和来源,为估算自然与人类活动影响下黄河三角洲土壤碳库储量提供基础数据和科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄河三角洲地区,属温带大陆性季风气候,四季分明,年均气温为11.9℃,年均降水量592 mm,年均蒸发量为1 944 mm,土壤类型以滨海潮土、滨海潮盐土为主[19],成土母质以黄河冲积母质和海相沉积物为主。黄河三角洲是我国三角洲中海陆变迁最活跃的地区,在黄河频繁改道、黄河冲积、海相沉积等自然过程和农业垦殖等人类活动影响下,由海向陆逐渐形成了无植被滩涂和养殖区、盐生植被湿地和旱地生态系统的演变梯度。其中,湿地主要植被类型有碱蓬(Suaeda heteropterKitag)、柽柳(Tamarix chinensisLour.)、芦苇(Phragmites communisTrin.)以及入侵种互花米草(Spartina alterniflora)。互花米草原本是主要分布在长江三角洲地区的典型湿地植被,20世纪90年代入侵黄河三角洲后迅速蔓延[20],在黄河三角洲自然保护区潮滩处已形成一定规模。旱地主要的粮食作物为小麦和玉米,经济作物主要为棉花,种植年限均超过50年。
1.2 样品采集
2015年10月在研究区采集土壤样品。选择沿黄河北侧由海到陆分布的3种土地利用类型,即无植被荒地(BS)、盐生植被湿地(WS)和旱地(FS)。其中,无植被荒地包括未利用滩涂和养殖底泥;湿地根据植被类型不同分为碱蓬、柽柳、芦苇和互花米草湿地;旱地包括棉田、麦田和玉米地。采样点分布图及其基本信息见表1。土壤样品使用不锈钢铲采集表层(0~20 cm)样品,每个采样点采用梅花布点采集5个样点,混匀带回实验室;一部分样品保存原样用于有机质分组,其余样品采用四分法取样,混匀、在室内风干,去除植物根系、动物残体、碎石、贝壳等,过10目尼龙网筛(2 mm),保存,用于基本理化性质的分析或前处理。
1.3 土壤基本理化性质和稳定性碳同位素分析
土壤样品基本理化性质的测定参考文献[21]的方法。其中土壤全盐含量采用重量法测定,每个采样点样品做3次重复,并计算其标准差;土壤pH采用电导法测定(水土比2.5∶1);土壤颗粒组成分析前去除土壤有机质和碳酸盐,之后采用激光粒度仪(Marlvern Mastersizer 2000F)测定,黏粒的粒径范围为<0.002 mm,粉砂为0.002~0.02 mm,砂粒为0.02~2 mm;土壤全氮、有机碳采用元素分析仪(Elementar Vario Micro cube)测定,乘以换算系数1.724得到有机质含量;土壤稳定性碳同位素采用同位素质谱分析仪(MAT253,Thermo Fisher)测定。在有机碳和稳定性碳同位素测定前,需要去除无机碳,即取约1 g样品于玻璃离心管中,加入5 mL 0.5 mol·L-1的盐酸,间断性涡旋震荡8 h直至无气泡以充分去除无机碳,重复2次,离心倒掉上清液;再用超纯水离心洗涤多次去除剩余盐酸,直至洗涤液为中性;50℃烘干后,待测。各指标在测定时,均做3次平行,测量标准偏差均小于0.5%。
1.4 土壤颗粒态有机质分组
土壤有机质分组参考文献中的方法[22]。首先,通过湿筛将土壤分为大团聚体和微团聚体。即称取20g挑除植物根系、贝壳、碎石等的土样于100 mL离心管中,缓慢加水浸润,水土比约为2∶1,此过程防止风干土中快速加水造成团聚体孔隙内气压不稳,团聚体崩裂,破坏其稳定性。然后将离心管倒置在2 mm筛(2 mm筛下层依次放置配套的250μm和53μm筛)水面以下,直至土样完全沉入到筛中。上下移动筛子,3 cm·次-1,25次·min-1。通过湿筛分离得到粒径为250~2 000μm大团聚体和53~250μm的微团聚体样品,再进一步分离颗粒态有机质;<53μm组分离心收集后用于分析矿物结合态有机质组分(mSOM),以上样品经50℃烘干,待用。每个采样点土壤样品做3次重复,准确称量烘干后的大团聚体和微团聚体质量,用于计算团聚体百分比及颗粒态有机质分配比。
在土壤团聚体分离的基础上,采用密度为1.85 g·cm-3的NaI溶液对大团聚体和微团聚体进行密度分级,即取5 g团聚体样品于100 mL离心管中,固液比为1∶7,上下颠倒1 min,静置30 min后,过滤分离上层轻组有机质,下层样品重复上述步骤,直至完全分离轻组有机质,得到大团聚体(250~2 000μm)和微团聚体(53~250μm)的游离颗粒态有机质(fPOM)。下层重组有机质用5 g·L-1的六偏磷酸钠分散16 h后湿筛,得到团聚体内部结合的颗粒态有机质(iPOM),各组分样品标记为:fPOM(250~2 000 μm)、iPOM(250~2 000 μm)、fPOM(53~250 μm)、iPOM(53~250 μm)以及mSOM。分离得到的这些组分均用超纯水反复洗去盐分离子,50℃烘干。每个样品的分离过程均做3次重复,准确称量烘干后的各有机质组分质量,并计算其标准差。土壤颗粒回收率大于97.5%。
1.5 数据处理
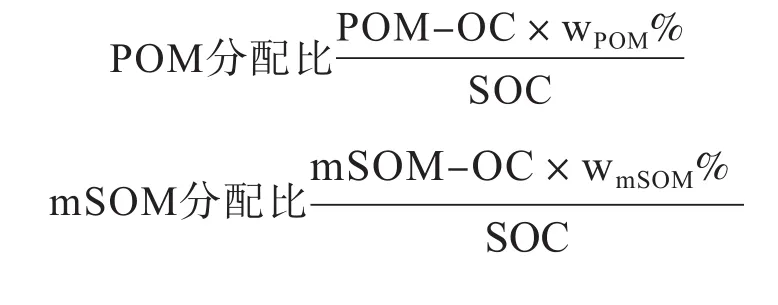
颗粒态有机质分配比的计算[23]:式中,POM-OC和mSOM-OC分别为颗粒态有机质(fPOM或者iPOM)和矿物结合态有机质中有机碳的含量,g·kg-1;wPOM%和wmSOM%分别为土壤中颗粒态有机质和矿物结合态有机质的质量分数(无量纲);SOC为土壤有机碳的含量,g·kg-1。
不同土壤有机质的陆源有机碳比例(f)采用基于有机碳同位素的二元端元模型[24]计算:

式中,δ13C海相为δ13C的海相端元值,δ13C陆相为δ13C的陆相端元值,δ13C样品为样品的δ13C值。陆地典型C3植物的δ13C值变幅为-32‰~-21‰,δ13C陆相采用其平均值为-27‰[24];近海生物包括海蜇、水蚤、浮游生物以及底栖生物等的δ13C值,变幅为-21.6‰~-20.3‰,δ13C海相采用其平均值,为-20.9‰[25]。
采用Microsoft Excel 2003软件和Origin 9.0软件对数据进行处理和绘图;用SPSS 20统计分析软件对数据进行差异显著性检验(LSD法,α=0.05)和相关性分析(单因素ANOVA法,α=0.05)。
2 结果与讨论
2.1 从潮滩到内陆土壤有机质与团聚体含量变化特征
从表2可以看出,不同土地利用类型下土壤有机质含量差异大。总体上,无植被荒地土壤有机质含量较低,均小于3.5 g·kg-1。向陆地延伸,随着湿地植被的生长,土壤有机质含量呈增加趋势;与无植被的荒地相比,湿地中碱蓬生长在潮间带,生物归还量低,而且植被凋落物容易被潮水带走[7],土壤有机质含量与无植被荒地相比变化不明显。柽柳湿地土壤有机质含量稍高,但小于5 g·kg-1,这可能由于柽柳生长密度和生物归还量较低,导致土壤有机质积累少[26]。互花米草湿地土壤有机质含量高达21.6 g·kg-1。自90年代初期互花米草入侵黄河三角洲地区[27],虽然同样生长在潮间带,但互花米草生物量大,而且促淤能力强,可以截留潮水携带的有机质[28]。芦苇湿地土壤有机质含量最高,达到45.2 g·kg-1,这主要是由于芦苇生长地势较高,受潮水作用相对较小,而且生物量大,植被凋落物归还量大,土壤固碳量高[29-30]。旱地土壤中,棉田土壤有机质含量最低,仅为6.9 g·kg-1,与棉田相比,麦田和玉米地土壤有机质含量有增高趋势,分别为19.7 g·kg-1和26.4 g·kg-1,这可能是由于小麦与玉米轮作过程中秸秆还田以及长期施肥增加了土壤有机质含量[31-32]。
研究区土壤pH变幅在7.6~8.8之间,普遍呈碱性。无植被荒地和盐生植被湿地土壤,由于受到海水影响,含盐量较高,多在10 g·kg-1以上,最高可达37.8 g·kg-1。旱地土壤,由于长期受到耕作、灌溉影响,盐分下降快,均在3.5 g·kg-1以下。
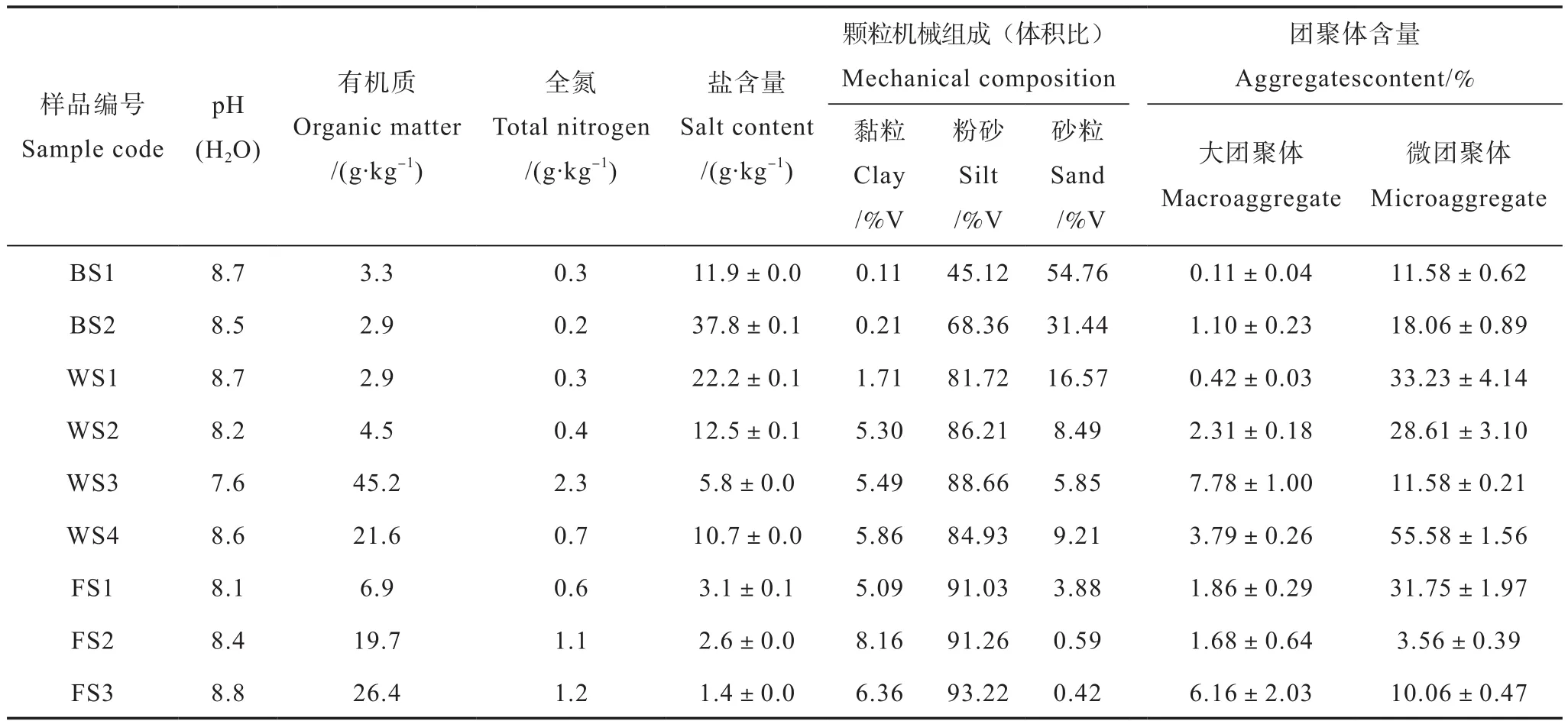
表2 供试土壤的基本理化性质及团聚体组成Table 2 Basic physico-chemical properties and aggregatecomposition of the tested soils
由表2可以看出,不同土地利用类型下土壤大团聚体和微团聚体的含量均存在较大差异。总体上,无植被荒地土壤大团聚体含量较低,滩涂土壤中几乎不存在大团聚体,养殖底泥中大团聚体含量仅为1.1%。与无植被荒地相比,盐生植被湿地土壤大团聚体含量呈增加趋势,芦苇湿地土壤大团聚体含量最高(7.78%)。湿地经农业垦殖后土壤大团聚体含量降低,棉田土壤和麦田土壤大团聚体含量均小于2%,但玉米地土壤大团聚体含量较高(6.16%)。有研究表明,短期玉米轮作对提高有机质含量和大团聚体形成都有良好的作用[33]。3种土地利用类型下土壤微团聚体含量均高于大团聚体含量。无植被荒地土壤微团聚体含量分别为11.58%和18.06%。盐生植被湿地中,互花米草湿地土壤微团聚体含量最高、平均为55.58%,其次为碱蓬和柽柳湿地,而芦苇湿地土壤微团聚体含量最低。滨海土壤旱地的耕作过程可能不利于土壤微团聚体的形成,与棉花单季作物土壤相比,麦田和玉米地土壤微团聚体含量明显较低。
土壤大团聚体含量与有机质含量呈显著正相关(r=0.91,P<0.05;图1),而与微团聚体含量的相关性不显著(r=0.30,P>0.05)。根据Six等[34]提出的团聚体周转概念模型解释,微生物利用新鲜输入的动、植物残体作为碳源,其分泌产物是团聚体形成的重要胶结物质;大团聚体中较大的颗粒态有机质分解、破碎成较小的颗粒态有机质,而后逐渐被黏粒和微生物产物包围,形成微团聚体;随着外界的干扰,稳定的大团聚体被破坏,逐渐释放出微团聚体。因而,大团聚体包被更多的有机质,而微团聚体中黏粒含量的增加是其与有机质含量相关性不显著的主要原因。
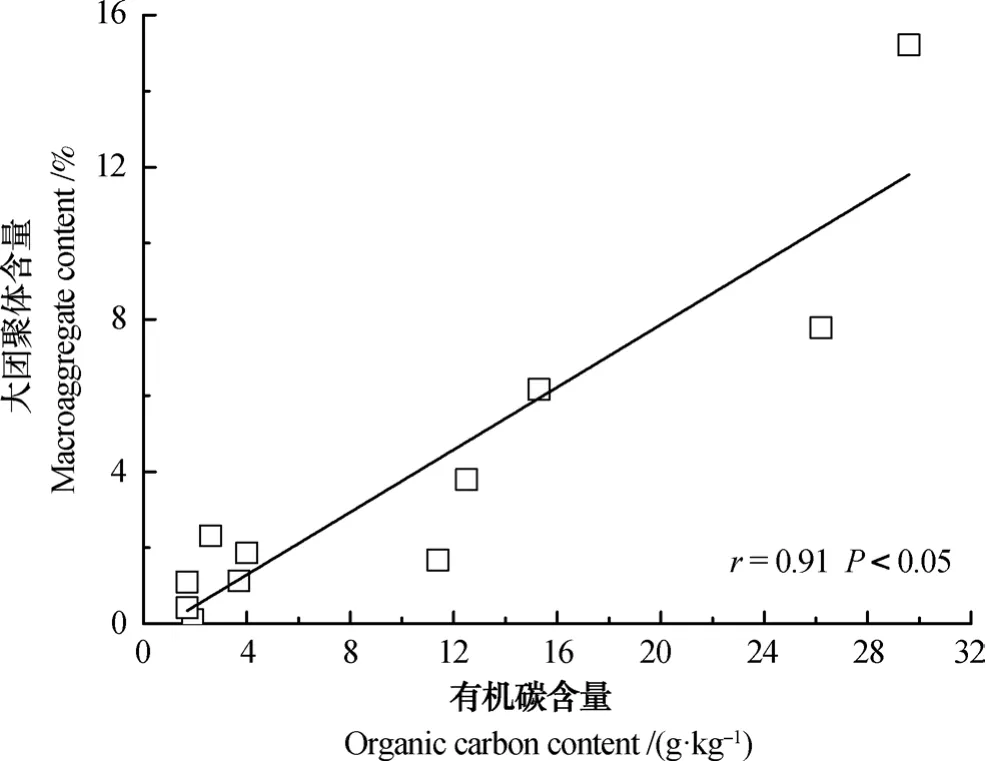
图1 黄河三角洲土壤大团聚体与有机碳含量之间的关系Fig. 1 Relationship between soil maroaggregates and organic carbon content in the soils of the Yellow River Delta
2.2 从潮滩到内陆土壤团聚体有机质中有机碳含量变化特征
颗粒态有机质大多是进入土壤中的动、植物半分解产物[34]。总体而言,在不同土地利用方式演变过程中,土壤颗粒态有机质和矿物结合态有机质的有机碳含量存在较大差异。不同土地利用类型土壤中游离颗粒态有机质(fPOM)的有机碳含量较高(图2A),变幅为162.5~410.0 g·kg-1;无植被荒地土壤大团聚体结合的fPOM的有机碳含量与微团聚体结合的fPOM的有机碳含量差异不显著;随着湿地植被的生长,不同粒径团聚体结合的fPOM的有机碳含量表现出显著差异(除芦苇湿地外),碱蓬和柽柳湿地土壤微团聚体结合的fPOM含量显著高于大团聚体结合的fPOM,而互花米草土壤团聚体结合的fPOM的有机碳含量则呈相反的趋势。这主要是由于碱蓬和柽柳湿地土壤大团聚体周转速度慢,较大的动植物残体等颗粒态有机质有充足的时间破碎为较小的有机质碎片,富集在微团聚体表面[34],而互花米草生长时间短,大团聚体表面容易粘附更多的有机质组分[35]。由盐生植被湿地向旱地过渡,两种土壤团聚体结合的fPOM的有机碳含量呈降低的趋势,小麦-玉米轮作地区土壤大团聚体和微团聚体结合的fPOM的有机碳含量趋于稳定,且二者的含量无显著差异。
图2B可知,在3类土地利用类型中iPOM的有机碳含量普遍低于98.2 g·kg-1,大团聚体iPOM的有机碳含量显著高于微团聚体,而且盐生湿地大团聚体iPOM的有机碳含量明显高于无植被荒地;盐生植被湿地向旱地过渡的过程中,大团聚体iPOM的有机碳含量表现出明显的降低趋势,而微团聚体iPOM的有机碳含量则呈现逐渐增加的趋势。这主要是由于旱地开垦过程破坏了大团聚体的稳定性,大团聚体破碎后释放出更多的微团聚体。微团聚体iPOM由于受土壤黏粒矿物保护作用和微生物产物的胶结作用,与大团聚体iPOM相比,其内部结合的有机质生物可利用性较低[34]。
与颗粒态有机质不同,矿物结合态有机质的有机碳含量普遍较低。与无植被荒地的相比,湿地植被的生长增加了mSOM的有机碳含量,尤其是芦苇和互花米草湿地土壤中mSOM的有机碳含量较高,分别为17.9 g·kg-1和18.9 g·kg-1;过渡为旱地后,mSOM的有机碳含量相对稳定,棉田和小麦-玉米轮作区土壤mSOM的有机碳含量为3.9~7.7 g·kg-1(图2C)。

图2 不同利用类型土壤中颗粒态有机质(fPOM、iPOM)和矿物结合态有机质(mSOM)中的有机碳含量Fig. 2 Organic carbon contents in the soil particulate organic matter (fPOM and iPOM) and mineral-associated organic matter (mSOM)in the soils of the Yellow River Delta relative to land use
2.3 从潮滩到内陆土壤团聚体有机质分配特征与稳定性
研究区土壤有机质分配特征如图3所示。整体上,研究区多数土壤中fPOM分配比高于iPOM。在由无植被荒地向旱地演变的过程中,颗粒态有机质分配比总体上呈现出先增加、后降低的趋势。无植被区未利用滩涂和养殖底泥中颗粒态有机质分配比分别为11.9%和12.8%,均以矿物结合态有机质为主。
盐生植被湿地土壤颗粒态有机质分配比占35%以上,其中互花米草湿地的分配比最高,达到75.2%。土壤中颗粒态有机质主要来源于不同分解阶段的动植物残体,其含量和组成受植被生长和环境因素的共同影响。湿地植被普遍具有较长的生长季、较高的净光合作用速率以及相对较大的生物量和凋落物归还量。因此,有机质的输入量大[36],为土壤微生物活动提供了充足的碳源,促进了团聚体的形成。同时,湿地土壤-植物生态系统在长期淹水环境下,有机质腐殖化速度相对缓慢,有利于土壤团聚体的稳定。较高含量的团聚体(表2)表面粘附更多的fPOM,同时团聚体的保护作用也有利于iPOM的积累,这与侯翠翠等[37]的研究结果相一致。此外,不同的湿地生态系统有机质的积累和分解过程还受到水分、盐度等因素影响[26]。例如,碱蓬分布地势低洼、具有较高的土壤盐分含量[38],不利于微生物分解新鲜植物残体,难以形成较多的大团聚体(表2),同时土壤黏粒含量较低(1.71%),砂粒含量较高(16.57%),不利于团聚体内部颗粒态有机质的稳定存在[39],因而其颗粒态有机质分配比较低。
随着滨海盐碱土壤的开垦,土壤中颗粒态有机质分配比逐渐降低。棉田土壤中颗粒态有机质为54%,而玉米地和麦田土壤颗粒态有机质均在20%以下。这主要是由于频繁的人为耕作扰动破坏了土壤团聚体结构[18],使土壤颗粒态有机质失去了团聚体的保护而多以游离态形式存在[40],更容易被微生物矿化分解。玉米、小麦等粮食作物种植的农田土壤耕作管理活动较棉花等单季作物更为频繁,大团聚体周转快,形成稳定的微团聚体机会少[39](表2)。因而,麦田和玉米地土壤的颗粒态有机质分配比远低于棉田土壤。
2.4 从潮滩到内陆土壤团聚体有机质稳定性碳同位素特征与海陆来源
研究区土壤团聚体有机质组分中稳定性碳同位素特征如图4所示。不同有机质组分的δ13C值呈现出fPOM <iPOM <mSOM的趋势。随着有机质矿化和腐殖化程度的加深,13C贫化的有机质优先降解,13C富集的有机质优先累积[32]。这些趋势表明,研究区土壤有机碳周转过程中,fPOM更容易被生物体优先利用,iPOM次之,mSOM最难被利用。这与Six[39]的研究结果一致,即土壤颗粒态有机质对环境条件的改变更为敏感,特别是容易受到耕作等人为活动的影响而发生矿化,属于土壤中的非稳定态的有机质组分,而矿物结合态有机质是土壤中的稳定性有机质组分,可反映土壤的固碳潜力。
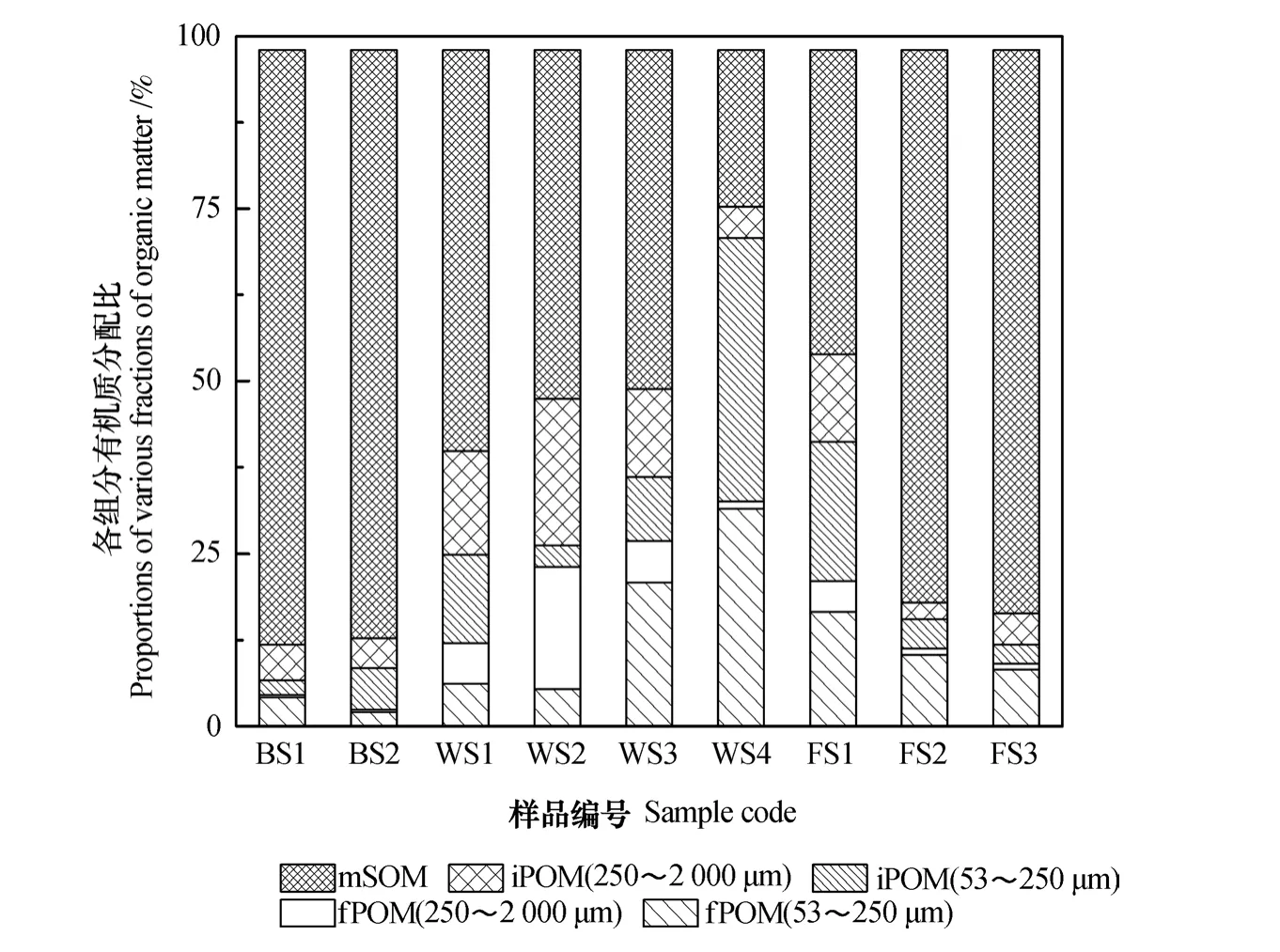
图3 不同采样区土壤中颗粒态有机质分配的百分比Fig. 3 Proportions of various fractions of soil POM in the soils relative to sampling area
同时,在无植被荒地、盐生植被湿地和旱地3种利用方式下,土壤大团聚体颗粒态有机质组分的δ13C值小于微团聚体颗粒态有机质组分的δ13C值,即δ13CiPOM(250~2000μm) <δ13CiPOM(53~250μm),δ13CfPOM(250~2000μm) <δ13CfPOM(53~250μm)。可见,微团聚体颗粒态有机质的生物可利用性小于大团聚体颗粒态有机质。Six等[39]报道了类似结果,即随着粒径减小,δ13C值越大,生物可利用性越低。结合图3中不同有机质组分的分配比特征可以得出,无植被荒地土壤有机质组分以矿物结合颗粒态为主,属于稳定碳库;向陆地延伸,盐生植被湿地土壤颗粒态有机质分配比较高,其有机质的生物可利用性较高,属于相对不稳定的有机质碳库;而在旱地开垦过程中,土壤颗粒态有机质分配比降低,土壤稳定碳库的比例相对较高。
不同土地利用类型下土壤各组分有机质的δ13C值呈如下趋势(图4):盐生植被湿地<旱地<无植被荒地。通常,陆地典型C3植物δ13C值处于-34‰~-23‰范围[24],海相生物的δ13C值处于-21.6‰~-20.3‰范围[25]。由于无植被荒地没有植被生长,并且该区分布于潮间带受海潮影响,因此,相比于旱地和湿地土壤,无植被荒地受到海源有机质的影响更大,表现为各组分中的δ13C值相对较高(图4)。通过同位素端源模型计算可得(图5),无植被荒地土壤不同组分有机质陆源有机质贡献率为25%~60%,并呈fPOM> iPOM>mSOM的变化趋势。而且大团聚体(250~2 000 μm)的颗粒态有机质的陆源贡献率大于微团聚体(53~250 μm)的颗粒态有机质,这一结果与章伟艳等[27]研究的河口土壤粗颗粒有机质易富集陆源植物碎片的结论一致。矿物结合态有机质组分则主要以海源有机质为主,与游离态颗粒态有机质不同。盐生植被湿地长期处于淹水环境下,土壤有机质的腐殖化速度相对缓慢[37]。随着旱地的开垦,在土壤有机质熟化过程中,有机质腐殖化程度提高,因而旱地土壤的有机质δ13C值大于湿地土壤。
3 结 论
总体而言,黄河三角洲由潮滩到内陆分布的无植被荒地、盐生植被湿地和旱地土壤的盐分含量逐步下降,有机质含量有所增加,但与植被类型、种植作物及其残留有关,无植被荒滩土壤的最低,芦苇湿地土壤的最高。土壤团聚体中有机质存在游离颗粒态有机质(fPOM)、团聚体内部结合的颗粒态有机质(iPOM)和矿物结合态有机质(mSOM),其有机碳含量分配及其稳定性与土地利用方式及植被类型有关。植被生长可以增加湿地土壤中的活性碳库,耕作可使旱地碳库更趋稳定。黄河三角洲滨海土壤,尤其是其大团聚体结合的活性碳库来源以陆源输入为主。

图4 不同土地利用方式下土壤颗粒态有机质和矿物结合态有机质中碳稳定同位素分布图Fig. 4 Biplots of δ13C of soil particulate organic matter and mineral-associated organic matter relative to land use

图5 无植被荒地土壤不同组分有机质的陆源有机碳百分比Fig. 5 Contributionof terrestrial organic carbon to different fractions of soil organic carbon in barren area
