酿酒酵母孢子作为GLP-1载体的研究
2019-04-24黄丹娣张慧杰冯诗倪中西秀树高晓冬
黄丹娣, 张慧杰, 闫 听, 冯诗倪, 中西秀树, 高晓冬
(江南大学 生物工程学院,江苏 无锡 214122)
糖尿病是目前危害人类健康的主要疾病之一,它是一种有遗传倾向的内分泌代谢疾病,以高血糖和高血糖继发的脂肪、蛋白质、水、电解质紊乱为特征。最新数据显示糖尿病患者的人数将在2030年上升到3.6亿[1]。而在我国,近年来随着人们生活水平的提高,糖尿病的发病率迅速提高,患病人数已经超过了1亿人,其中,绝大多数为二型糖尿病患者。糖尿病可引起包括心脑血管疾病在内的多种并发症,因此,糖尿病的防治已成为人们最为关注的热点之一。
二型糖尿病又被称为非胰岛素依赖糖尿病,患者本身仍产生胰岛素,然而由于胰岛β细胞功能低下和胰岛素抵抗,患者对胰岛素的利用却大打折扣[2]。目前,二型糖尿病患者的治疗手段主要包括使用胰岛素和降糖药物等。虽然这些药物能够有效降低血糖,但是也存在一些问题,例如会导致患者低血糖以及体重的增加等。此外,这些药物只能延缓病情,无法恢复胰岛β细胞功能,从而无法从根本上使病情得到控制。因此,开发安全有效的糖尿病治疗药物,已成为现代科学家的研究热点[3]。
胰高血糖素样肽-1(Glucagon-like peptide-1,GLP-1)含有37个氨基酸,是已发现的促胰岛素分泌作用最强的肠肽类激素。GLP-1的生物活性形式有2种,分别为GLP-1(7-37)和GLP-1(7-36)酰胺。GLP-1可以从多个方面发挥作用,如抑制胃排空,抑制胰高血糖素的分泌,降低血糖等[4],并且能够恢复胰岛β细胞功能。更为重要的是,GLP-1的作用具有葡萄糖浓度依赖性,仅在葡萄糖浓度较高时才发挥作用,因此不会导致由于使用过度而导致的低血糖问题。GLP-1已成为未来治疗二型糖尿病的安全有效的多肽药物[5]。然而天然的GLP-1(7-36)容易被二肽基肽酶(DPP-IV)降解为无活性的GLP-1(9-36),且由于其相对分子质量较小从而易被肾清除,因此导致GLP-1在人体内半衰期不足2 min,这大大限制了其临床应用。因此,科学家正致力于研究如何延长其在体内的半衰期从而提高其生理活性。目前的研究主要集中在以下几个方面:筛选GLP-1类似物,例如艾塞那肽,利拉鲁肽[6];对天然GLP-1进行结构改造,如GLP-1的N端修饰、脂肪酸修饰GLP-1等[7];将GLP-1与生物大分子蛋白偶联,如人清蛋白[8],IgG重链结合蛋白IgG-Fc[9]等,以及将GLP-1串联表达等,均取得了一定的效果。此外,传统的多肽类药物以及胰岛素需要注射使用,这让病人痛苦不堪。研究发现,GLP-1可以以活性形式被肠道直接吸收进入血液,通过血液循环直接与胰岛细胞表面的GLP-1受体结合并最终促进胰岛素的释放[10]。因此,若能够通过一些方法延长其在体内的半衰期,提高其利用率,并选择合适的载体构建口服GLP-1药物,将为有效治疗糖尿病提供更有力的方法,并具有巨大的科学意义和市场潜力[11]。
酿酒酵母是单细胞真核生物,其完整的基因组在1996年便已完成测序。酿酒酵母培养简单,生长迅速,且易于进行基因改造,被最早应用于酵母基因克隆和表达宿主菌及人源化蛋白[12]。酿酒酵母的细胞有2种生活形态,单倍体和二倍体。二倍体细胞可在营养匮乏的条件下减数分裂生成孢子,一个母细胞可分裂成4个孢子。酿酒酵母孢子易于获得,安全无毒,可以食用。酿酒酵母的孢子壁与母细胞相比有明显的不同。如图1所示,成熟的孢子壁从内到外分别是甘露糖层、β-葡聚糖层、壳聚糖层和二酪氨酸层。而酿酒酵母从内到外仅有β-葡聚糖层、甘露糖层。孢子特有的二酪氨酸层具有保护孢子抵御外界环境压力的能力。因此,在口服过程中,孢子壁可以保护孢内表达的外源蛋白抵抗一定的蛋白酶降解及胃酸低pH的影响。
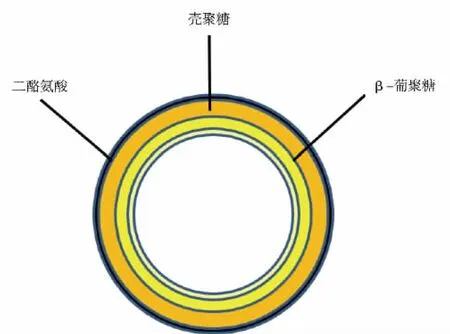
图1 酿酒酵母孢子壁的结构Fig.1 Structure of yeast spore wall
本研究拟利用酿酒酵母孢子构建GLP-1可口服给药体系。通过串联表达GLP-1,突变GLP-1部分碱基等方式对GLP-1进行改造,并同时将改造后的GLP-1表达到酿酒酵母孢子内以提高GLP-1抗酶解能力和体内半衰期。通过模拟肠胃液处理实验,体外刺激小鼠胰岛β细胞分泌胰岛素等实验证实了所构建的GLP-1给药体系的有效性。
1 材料与方法
1.1 实验材料
1.1.1 菌株和质粒 酿酒酵母AN120野生型菌株,dit1Δ缺陷型菌株,小鼠胰岛β细胞株MIN6细胞(江南大学食品学院孙佳教授馈赠),大肠杆菌DH5α。
pBlueScriptⅡSK(+)质粒,pRS424-TEFpr为本实验室保存质粒,pRS424-TEFpr-8*GLP-1-GFP和pRS424-TEFpr-ss-8*GLP-1-GFP为本研究所构建的质粒。
1.1.2 培养基 LB培养基:酵母提取物10 g,胰蛋白胨10 g,氯化钠 10 g,琼脂粉 20 g(固体),加水至1 L,121℃高压灭菌20 min。
SD-Trp培养基:无氨基酵母氮源(YNB)6.7 g,琼脂粉20 g(固体),加水至900 mL,121℃高压灭菌20 min,使用前加入2 g缺少色氨酸(Trp)的氨基酸粉末以及100 mL灭菌的20 g/dL的葡萄糖溶液。
YPAD培养基:酵母提取物10 g,蛋白胨20 g,腺嘌呤0.03 g,琼脂粉20 g(固体),加水至900 mL,121℃高压灭菌20 min,使用前加入100 mL灭菌的20 g/dL的葡萄糖溶液。
YPACe培养基:酵母提取物10 g,蛋白胨20 g,腺嘌呤0.03 g,琼脂粉20 g,乙酸钾20 g,加水至1 L,121℃高压灭菌20 min。
产孢培养基:乙酸钾20 g,加水至1 L,121℃高压灭菌20 min。
1.2 实验方法
1.2.1 大肠杆菌质粒的构建 GLP-1的活性区域含有36个氨基酸,其氨基酸序列从NCBI中获得,如图2所示。为了防止其被二肽基肽酶(DPP-IV)和胰蛋白酶(Trypsin)降解,我们对其非保守序列的第2位、第20位、第28位的氨基酸进行了修改优化,已有文献证明这3个位点的改变并不影响GLP-1的生理活性。同时,为了在孢子内大量表达GLP-1并防止GLP-1被降解,将其4串联表达,并在序列首尾设计了相应的酶切位点,以方便后面的基因操作。此外,为了防止同源重组,对不同串联序列的一些碱基也做了微调,整个4*GLP-1序列也根据酿酒酵母偏爱密码子进行了优化,并由上海生工生物公司合成。
合成的4*GLP-1序列首尾两端各设计了2个酶切位点,分别为BamHⅠ、SmaⅠ、EcoRⅠ以及EcoRⅤ。大肠杆菌质粒表达载体的构建流程如图3所示。由于pRS424-TEFpr质粒本身有2个EcoRⅤ酶切位点,无法用于直接构建8*GLP-1重组质粒,因此选择pBlueScriptⅡSK(+)作为出发质粒。首先为了方便检测目的蛋白的表达和定位,以pRS424-TEFpr-GFP为模板,通过PCR的方法将绿色荧光蛋白GFP编码序列连接到载体,分别在5’和3’端加入EcoRⅠ以及XhoⅠ酶切位点。引物序列如下:
上游引物:
5’-CCGGAATTCAAAGGAGAAGAACTTTTCAC-3’下游引物:
5’-CCGCTCGAGTTATTTGTATAGTTCATCCATGC-3’
PCR获得目的条带之后,通过酶切连接的方法,将GFP基因序列连接到pBlueScriptⅡSK(+)载体上,转化大肠杆菌DH5α,挑取4-6个单菌落于LB-AMP液体培养基中,于37℃摇床过夜培养,随后通过菌液PCR鉴定阳性转化子,再提取阳性转化子的质粒,进行酶切验证。
接着为构建pBlueScriptⅡSK (+)-4*GLP-1-GFP,用BamHⅠ、EcoRⅠ双酶切4*GLP-1条带和pBlueScriptⅡSK(+)-GFP重组质粒,连接产物转化大肠杆菌DH5α,于LB-AMP培养基挑取阳性转化子。为构建重组质粒pBlueScriptⅡSK(+)-8*GLP-1-GFP,用 SmaⅠ、EcoRⅤ双酶切目的条带,用 SmaⅠ单酶切重组质粒pBlueScriptⅡSK (+)-4*GLP-1-GFP,获得两端为平末端的片段,连接转化大肠杆菌DH5α,挑取一定数量的单菌落进行酶切验证。并挑取阳性单菌落于LB-AMP液体培养基中37℃过夜培养以扩增重组质粒,重组质粒用小提质粒试剂盒提取。
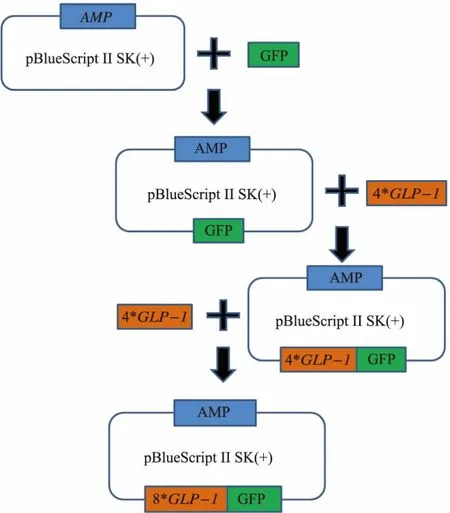
图3 质粒构建流程Fig.3 Construction of plasmid pBlueScriptⅡSK (+)-8*GLP-1-GFP
1.2.2 酿酒酵母质粒表达载体的构建 为使目的蛋白能够在酿酒酵母孢子内表达,首先,需构建酿酒酵母转化质粒pRS424-TEFpr-8*GLP-1-GFP,通过BamHⅠ、XhoⅠ双酶切重组质粒将8*GLP-1-GFP序列切下来再连接到pRS-424-TEFpr表达载体上,连接产物转化大肠杆菌,为使目的蛋白8*GLP-1能够定位在酿酒酵母孢子壁上,需在目的蛋白前添加信号肽序列ss片段。因ss片段大小不足100 bp,本文采用将TEF启动子与ss序列一同PCR的方法,获得TEFpr-ss片段以替代重组质粒上的TEF启动子序列。以pRS424-TEFpr-ss-3HA为模板,通过PCR的方法获得TEFpr-ss序列,分别在两端引入SacⅠ和BamHⅠ酶切位点。引物序列如下:上游引物:
5’-GCTGGAGCTCATAGCTTCAA-3’
下游引物:
5’-CCGGATATCGGAAACAGGATTACAG-3’
最终获得pRS424-TEFpr-8*GLP-1-GFP以及pRS424-TEFpr-ss-8*GLP-1-GFP重组质粒 (图4-5),构建好的质粒由上海生工测序。
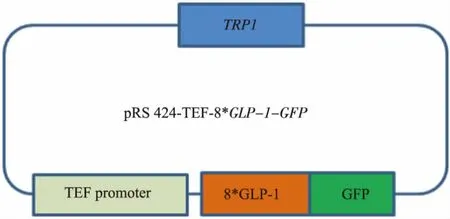
图4pRS424-TEFpr-8*GLP-1-GFP重组质粒Fig.4Plasmid pRS424-TEFpr-8*GLP-1-GFP
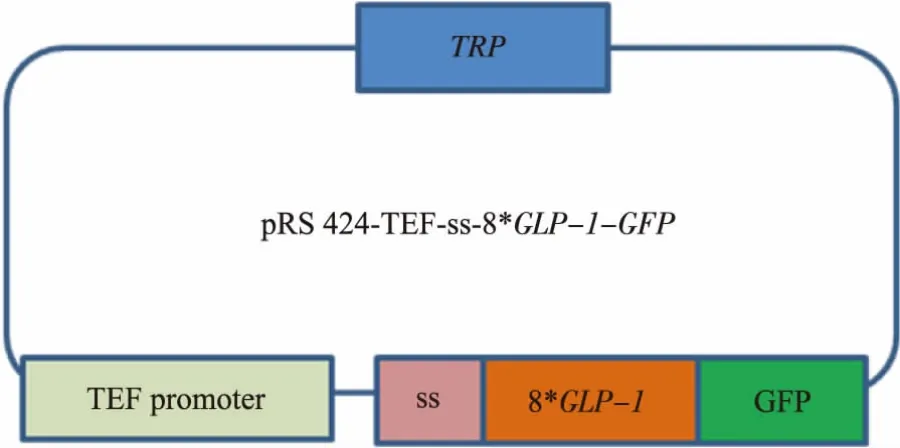
图5pRS424-TEFpr-ss-8*GLP-1-GFP重组质粒Fig.5 Plasmid pRS424-TEFpr-ss-8*GLP-1-GFP
1.2.3 GLP-1重组蛋白在酿酒酵母孢子的表达以及孢子的纯化 将得到的重组质粒通过一步转化法转入到酿酒酵母后,涂相应缺陷平板,2天后长出的单菌落即为成功转入重组质粒的酵母。挑取一个单菌落接种于5 mL SD-Trp培养基的试管中,并于30℃、220 r/min的摇床过夜培养。12 h后取1 mL菌液转入盛有50 mL的YPACe液体培养基的250 mL三角瓶中,于30℃、220 r/min摇床预产孢24 h;6 000 r/min离心2 min收集菌体,用产孢培养基重悬,并全部转入250 mL的三角瓶中(无菌操作),于30℃,220 r/min摇床培养 24 h;取 5~10 μL产孢培养基中的菌液,于显微镜下观察酵母产孢情况,正常可以看到视野中有大量四联球形态的子囊细胞。
为了使单个孢子能够从子囊孢子中释放出来,需对重组酵母的子囊孢子进行进一步处理,8 000 r/min离心收集孢子,并重悬于2 mL原生质体buffer中,充分混匀后加入约 100 μL 的 lyticase(1 mg/mL),并置于37℃的摇床处理3 h,随后超声破碎以获得单个孢子。为进一步除去孢子部分破碎的子囊壁和裂解酶,以获得纯净的孢子。需对超声破碎的孢子进行纯化处理:首先需配制体积分数50%、60%、70%、80%梯度的Percoll溶液,配方如下:5mL Pecoll、4mL体积分数0.5%Triton X-100,1 mL 2.5 mol/L蔗糖;6 mL Pecoll,3 mL 体积分数 0.5%Triton X-100,1 mL 2.5 mol/L 蔗糖;7 mL Pecoll,2 mL 体积分数 0.5%Triton X-100,1mL 2.5 mol/L 蔗糖;8 mL Pecoll,1 mL体积分数 0.5%Triton X-100,1 mL 2.5 mol/L蔗糖。接着将超声破壁过的孢子悬浮液用5 mL体积分数0.5%Triton X-100洗涤一遍,并溶解在1 mL的体积分数0.5%Triton X-100中,将体积分数80%、70%、60%、50%梯度的Percoll溶液各取 1 mL,沿离心管壁缓慢依次加入10 mL离心管中,最后加入1 mL溶解在体积分数0.5%Triton X-100中的孢子液;4℃、15 000 r/min离心约30 min后,梯度溶液会分层,用移液枪吸取孢子悬浮的一层于干净离心管中,用5 mL 0.6 mol/L NaCl溶液洗涤1次;接着用1 mL体积分数0.5%Triton X-100洗涤2次;最后将纯化后的孢子置于冷冻干燥机中(-50℃,72 Pa)24 h后取出备用。
1.2.4 GLP-1重组蛋白的检测 为了检测目的蛋白是否成功表达,需提取重组酵母孢子内的全蛋白进行Western blot检测,具体方法为:离心收集约5 mL培养基的菌体于1.5 mL离心管中,用dd水洗2遍后用500 μL S buffer重悬,再加入5 μL的溶壁酶(lyticase),于30℃酶解30 min。离心收集沉淀并用 S buffer洗2遍后,加入400 μL lysis buffer重悬,再加入适量玻璃珠以及4 μL PMSF (100 mmol/L),于冰上破碎,具体步骤为冰上静置1 min,再于振荡器振荡1 min,如此循环15次后,于4℃、1 000 g离心10 min,取上清液3 000 g离心10 min进一步去除细胞碎片。所得的上清液即为提取的酿酒酵母全蛋白。全蛋白用Western blot进一步检测目的蛋白是否表达,Anti-GFP mouse和 Goat Anti-mouse lgGHRP分别作为一抗和二抗。
在Western blot检测到目的条带后,通过荧光显微镜检测荧光信号来进一步确定GLP-1在孢子中的表达位置。首先重接种酿酒酵母并产孢,产孢过程同上,获得四联球孢子后,收集约5 mL菌液的孢子,并用dd水洗2遍后重悬,调节一定的浓度,取5 μL于载玻片上,并尽快置于荧光显微镜下,使用100×油镜观察检测GFP荧光信号,获得的荧光照片由NIS-Element AR软件分析。
1.2.5 酿酒酵母孢子给药体系的体外活性检测在确定目的蛋白成功表达后,接下来要对酿酒酵母孢子给药体系的活性进行检测。为模拟人体内部消化环境,首先用模拟肠胃液对孢子载药体系进行处理。处理流程图如图6所示,首先,称取10 mg孢子粉,溶于10 mL模拟胃液中,于37℃、180 r/min摇床消化处理2 h,其中模拟肠胃液的配方为:2.0 g氯化钠、3.2 g胃蛋白酶、7 mL HCL,溶解于 1 L ddH2O。随后于模拟胃液中加入NaHCO3并调节pH至7.5,再加入胰酶至终质量浓度为10 g/L,配制成模拟肠液,同样于37℃摇床处理4 h。随后为使蛋白酶失活,将样品置于金属浴中100℃热处理5 min,然后迅速冷却至室温。离心管于3000r/min离心20min,收集上清液后置于真空冷冻干燥机中冻干备用。
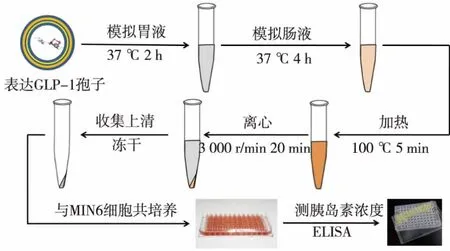
图6 给药体系体外活性检测过程Fig.6 Detection of in vitro activity of the drug deliverysystem
为检测经模拟肠胃液处理后的消化产物是否具有促胰岛素释放的作用,需将冻干产物与MIN6细胞共培养并检测上清液中的胰岛素浓度。将MIN6细胞接种于96孔板上,接种密度为2×104个/孔,分为空白对照组和实验组,每组3个平行样。培养24 h后,取冻干的消化产物,重悬于100 μL的细胞培养基中,再取10 μL加入96孔板的样品孔中,与MIN6细胞共培养2 h,随后取上清液,并按照小鼠胰岛素ELISA试剂盒的操作步骤进行胰岛素水平的测定。
2 结果与分析
2.1 重组质粒的构建
为了能够将GLP-1表达到酿酒酵母内,需要构建pRS424-TEFpr-8*GLP-1-GFP以及pRS424-TEFpr-ss-8*GLP-1-GFP重组质粒,为使重组蛋白能够定位在酿酒酵母孢子壁上,需在目的蛋白前添加信号肽序列ss片段。因ss片段大小不足100 bp,本文采用将TEF启动子与ss序列一同PCR的方法,获得TEFpr-ss片段以替代重组质粒上的TEF启动子序列。以pRS424-TEFpr-ss-3HA为模板,通过PCR的方法获得TEFpr-ss序列,分别在两端引入SacⅠ和BamHⅠ酶切位点。最终获得pRS424-TEFpr-8*GLP-1-GFP以及pRS424-TEFpr-ss-8*GLP-1-GFP重组质粒,构建好的质粒首先经过SacⅠ和 BamHⅠ双酶切验证(图 7),图 7 (a)在 2号泳道有一条约1 490 bp的条带,与序列8*GLP-1-GFP的长度一致。图7(b)亦在500 bp(序列TEFprss)处检测到条带。在酶切验证后重组质粒由上海生工进行进一步的确认。

图7 重组质粒酶切电泳Fig.7 Agarose gel electrophoresis of recombinant plasmid
2.2 8*GLP-1-GFP重组蛋白的表达
为检测目的蛋白是否成功表达,我们将pRS424-TEFpr-8*GLP-1-GFP和pRS424-TEFpr-ss-8*GLP-1-GFP重组质粒转化到酿酒酵母AN120野生型菌株,dit1Δ缺陷型菌株内,产孢获得相应重组酵母孢子并提取蛋白。利用Western blot检测目的蛋白条带,从图 8 可看出,在 55×103(8*GLP-1-GFP)左右有明显的条带。在野生型中,相对于胞内表达的重组体系,表达在孢子壁上的目的蛋白量明显较多,可能是由于表达到孢子壁上的重组蛋白可防止被内部酶解。而在二酪氨酸缺陷型孢子中,情况正好相反,可能是由于其缺少最外层的二酪氨酸,从而导致大量的重组蛋白分泌到胞外。
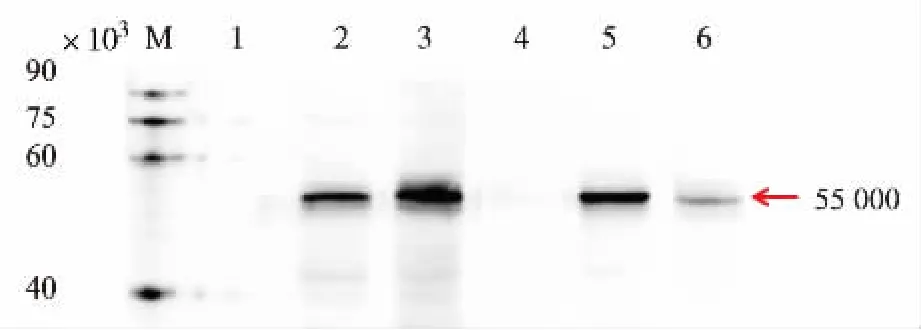
图8 8*GLP-1-GFP重组蛋白表达的Western blot验证Fig.8 Western blotting analysis of 8*GLP-1 expressed in yeast spore
2.3 8*GLP-1-GFP重组蛋白的在酿酒酵母孢子的定位
我们进一步通过GFP的荧光定位来确定GLP-1 在孢子中的位置。 图 9 中(b)、(e)为胞内表达,可以检测到明显的荧光,(c)、(f)为胞子壁表达,其荧光强度明显低于胞内表达,且似乎有少量泄露到子囊壳内。荧光定位实验进一步证明了重组蛋白8*GLP-1被成功表达到酿酒酵母孢子内。
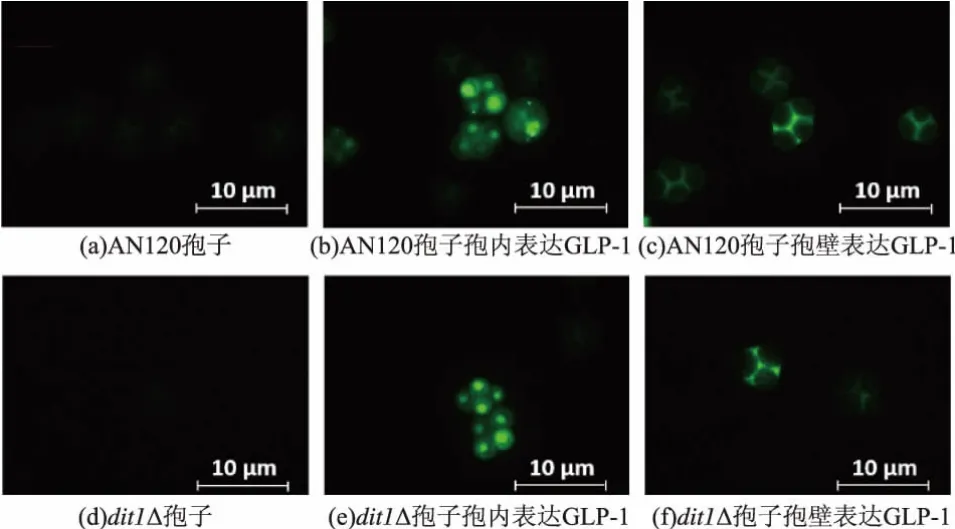
图9 8*GLP-1-GFP重组蛋白在孢子内的荧光定位Fig.9 Localization of 8*GLP-1-GFP in yeast spore
2.4 酿酒酵母孢子载药体系的体外活性检测
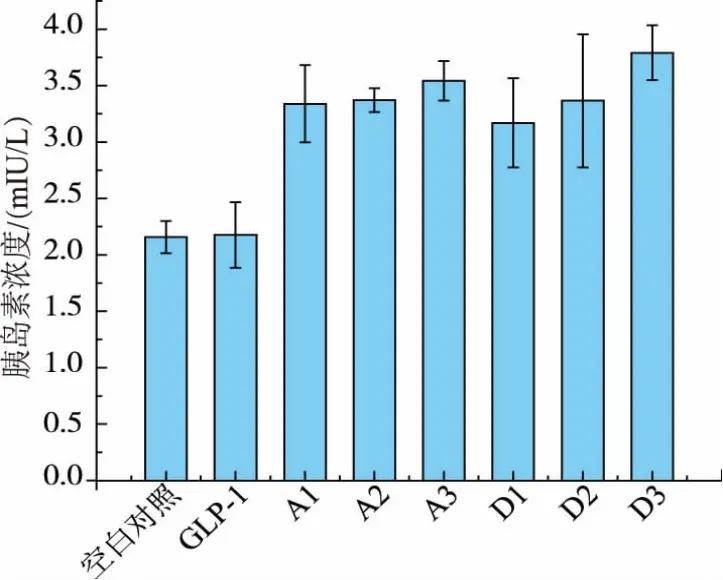
图10 GLP-1刺激MIN6细胞分泌胰岛素结果Fig.10 Insulin secretion stimulated by GLP-1
在确定目的蛋白成功表达后,我们进一步对酿酒酵母孢子GLP-1载药体系的体外活性进行检测。将经模拟肠胃液处理后的载GLP-1孢子与MIN6共培养,并用ELISA方法检测胰岛素浓度。如图10所示,与单纯的GLP-1相比,表达了GLP-1重组蛋白的孢子能够显著促进胰岛素的分泌,同时,胞子壁表达与胞内表达的效果差别不大。说明无论是表达到孢子壁还是表达到孢内,孢子都可以对所表达的GLP-1具有一定的保护作用,并提高其促胰岛素活性。
3 结 语
糖尿病作为影响现代人生命健康的一大疾病,至今仍无有效的解决方法。GLP-1及其类似物已成为研究治疗二型糖尿病的热点。本文首先对GLP-1进行基因改造及串联表达,然后选择酿酒酵母孢子作为表达和运输载体构建GLP-1口服给药体系。本文构建了重组GLP-1质粒并成功在酿酒酵母孢子内表达,形成表达8串联GLP-1的酿酒酵母孢子载药体系。该体系在经过模拟肠胃液处理后能有效刺激MIN6细胞分泌胰岛素,证明了所构建的GLP-1给药体系的有效性。综上,本研究为进一步开发治疗糖尿病的口服药物提供了新的思路。
