湿地植物丝瓣剪秋罗幼苗对温度和土壤养分的表型可塑性响应
2019-04-17唐占辉
包 宇, 杨 苗, 唐占辉
(1. 国家环境保护湿地生态与植被恢复重点实验室, 长春 130117; 2. 东北师范大学 环境学院, 长春 130117)
表型可塑性是指同一种基因型个体通过对不同环境做出应答而产生不同表型的特征,是生物适应环境的一种被动表现[1-2]。自然环境中植物生长发育所需资源在环境中并非均质,如果一种基因型只存在一种表型的话,那么该物种就很难在异质的环境中表现出较高的适合度[3-5],植物通过生物量分配等表型可塑性来增加对资源的利用效率, 同时也为自身提供了一种有效地适应环境机制[6]。湿地处于陆地生态系统和水生生态系统的过渡区域,从而兼备两种生态系统的多种复杂生态因素,导致湿地植物特别是幼苗阶段对环境因子的变化具有不同的响应规律。特别是湿地植物幼苗对外界环境因子的表型可塑性变化,对其种群更新、生存和发展具有重要的意义[7-9]。
吉林省东部金川泥炭地是龙岗山脉中段的山间谷地形成的一块典型的泥炭沼泽湿地,该湿地曾受到较大人为干扰,近年来被加以保护。汤袁等人研究了该湿地的圆叶茅膏菜(Droserarotundifolia)在不同养分和生境条件下的生态可塑性特征,发现圆叶茅膏菜的年龄结构、腺毛密度、叶形态特征都有一定的可塑性规律[10]。丝瓣剪秋罗(L.wilfordii)是该湿地分布的一种石竹科(Caryophyllaceae)剪秋罗属(Lychnis)的多年生草本植物[11],野外调查发现其野外种群数量近年来下降,但对其保护和研究却远远落后。目前,对于剪秋罗属植物的研究主要集中在园林栽培、组织培养及多倍体育种等方面[12-15],而它们的幼苗生长发育对环境的响应,开展研究较少。本研究选择的对象为丝瓣剪秋萝,其幼苗生长对湿地环境因子(温度、土壤养分)的响应还没有相关研究。考虑到植物幼苗对环境因子的变异响应是保护湿地植物、恢复湿地生态工作需要解决的关键问题之一,因此本研究以湿地植物丝瓣剪秋罗幼苗为研究对象,研究在不同温度和土壤养分处理下,丝瓣剪秋罗幼苗生长状况、生物量积累和分配等特征。研究结果对认识湿地植物幼苗表型可塑性对环境因子变化的响应及日后湿地恢复工作具有重要意义,为更好地保育湿地植物丝瓣剪秋萝提供参考依据,也对深入了解剪秋萝属植物的生长发育特性具有重要的参考价值。
1 材料与方法
1.1 材料
本研究中丝瓣剪秋罗的种子是2016年冬天购于华夏种业公司并保存在干燥温室中,2017年4月初播种育苗。
1.2 方法
1.2.1 种子萌发和幼苗移栽
随机选取饱满且大小一致的丝瓣剪秋罗种子,将种子放入以泥炭土为基质的花盆(19 cm×14.5 cm×9.5 cm)中,在光照培养箱内做萌发实验。当幼苗长至5 cm时候,严格挑选长势良好且生长一致的幼苗移植到以蛭石作为基质的塑料花盆中(排除非处理因素影响),幼苗移植后经过缓苗阶段,待植物材料完全恢复正常后,再一次淘汰掉不符合条件的植株后,进行实验处理,样本量总共为54株植物幼苗。花盆底部具有排水孔,为防止土壤渗漏,花盆排水孔下方垫有细尼龙网,整体置于托盘上。控制实验期间幼苗均在可控的植物培养箱内生长。
1.2.2 实验处理方式
实验设置分为温度处理和营养处理(每个处理各27株)。温度处理设置3个梯度:分别为高温(30℃)、中温(20℃)和低温(10℃)。土壤养分处理设置3个梯度:分别为高营养(全强度改良霍格兰营养液)、中营养(1/2强度改良霍格兰营养液)、低营养(1/10强度改良霍格兰营养液)。以上处理方式下每个处理的样本量为9株植物,将9株幼苗分种在3个花盆(每盆3株幼苗),温度处理用控温自动调节功能设置;养分处理按照每个花盆每次加30 mL相应的营养液,每隔4 d加1次。实验期间,保持花盆基质的湿润状态,以保证幼苗能有效的吸收养分。每隔10 d取样测量1次,每次取1盆,将盆土倒出用水小心冲掉土壤以获得完整3株幼苗,进行下述指标的测量。
1.2.3 测量指标
测量指标时将幼苗分成地上、地下部分,清洗后用吸水纸吸干,在65℃烘干48 h,用1/10 000的电子天平称量,计算各项生长参数,各项参数计算方法如下[16]:
株高(cm)=从植物露出地面的基部开始,至植物顶点的高度;
总生物量(g)=根生物量(干重)+地上生物量(干重);
根冠比=根生物量(干重)/地上生物量(干重);
相对生长率=(LnMt2—LnMt1)/(t2—t1),其中M指幼苗干生物量,t表示不同时间g/(g·d);
表型可塑性指数=处理中某一性状的最高值减去最低值除以最高值。
1.3 数据分析
采用SPSS 19.0统计分析软件进行数据分析。在比较前,先检验数据是否符合正态分布。对于服从正态分布的数据,使用单因素方差分析(One-Way ANOVA)检验不同处理对植物各生长参数影响的显著性;对于不服从正态分布的数据,首先进行数据的转换,如果转换后数据服从正态分布,使用单因素方差分析检验相关变量间是否存在显著差异。若仍不服从正态分布,则使用非参数检验中的K个独立样本检验法,上述所有显著性检验的显著度均设为0.05。
2 结果与分析
2.1 不同温度下丝瓣剪秋罗幼苗表型可塑性
2.1.1 植株高度可塑性
丝瓣剪秋罗幼苗的株高在盆栽实验开始后第10天时,高温条件下幼苗增高较其他条件下明显,低温条件下株高增长最小;20 d时,高温条件较中温条件株高相差0.8 cm,低温条件下株高增长最小;30 d时,高温条件株高较其他条件下增高显著,并且低温条件下株高增高最小(图1)。不同温度处理条件下,第10天、第20天及第30天时幼苗株高均存在显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05),温度对丝瓣剪秋罗幼苗株高的影响显著。同时在10、20和30 d时均能观察到温度越高,幼苗株高越高的现象。
2.1.2 生物量积累和根冠比可塑性
在不同温度处理下,丝瓣剪秋罗幼苗的总生物量在3个不同时间点均有显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05)。第10天、第20天及第30天时,高温条件下幼苗总生物量均较其他条件下增加显著,并且低温条件下总生物量增加最小(图2)。结果表明幼苗总生物量在3个时间点均随温度升高而增加。温度对丝瓣剪秋罗植物幼苗的总生物量影响显著。
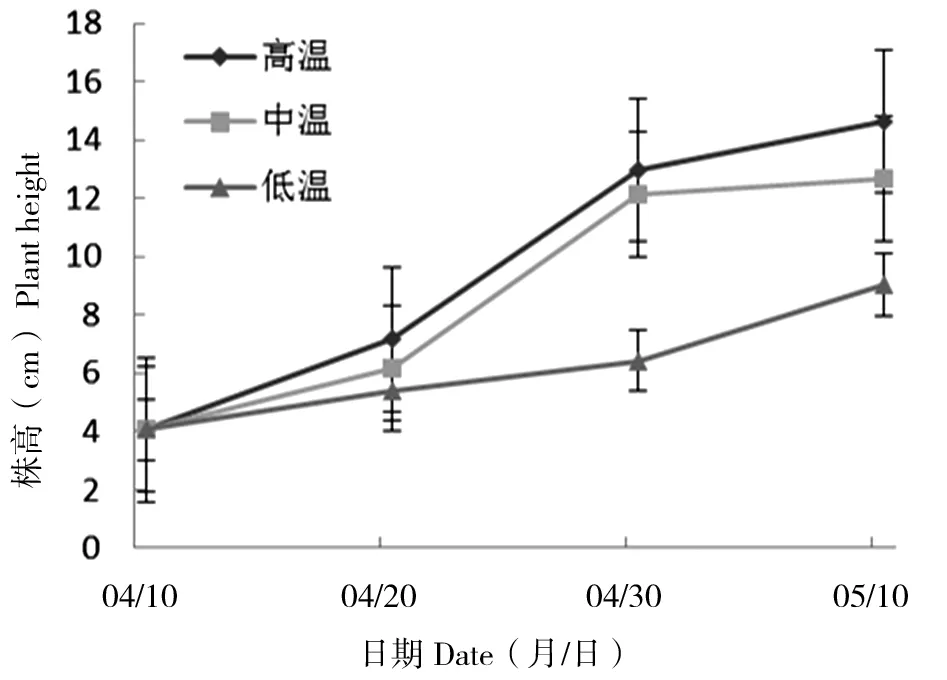
图1 丝瓣剪秋罗幼苗在3种不同温度处理下植株高度随生长时间的动态变化Figure 1 The temporal dynamics of plant height during thirty days under three temperature treatments

图2 丝瓣剪秋罗幼苗在3种不同温度处理下植株总生物量随生长时间的动态变化Figure 2 The temporal dynamics of plant biomass during thirty days under three temperature treatments
第10天、第20天和第30天时,高温条件下幼苗根冠比较其他条件下小,并且在低温条件下根冠比随时间变化下降最为缓慢,根冠比最高(图3)。丝瓣剪秋罗幼苗的根冠比在不同温度处理下,除第10天外,在第20天和第30天均存在显著差异(P20 d<0.05,P30 d<0.05)。不同温度处理条件下,幼苗根冠比表现为随温度升高而下降,温度对于丝瓣剪秋罗幼苗的根冠比的影响在后期更为明显。
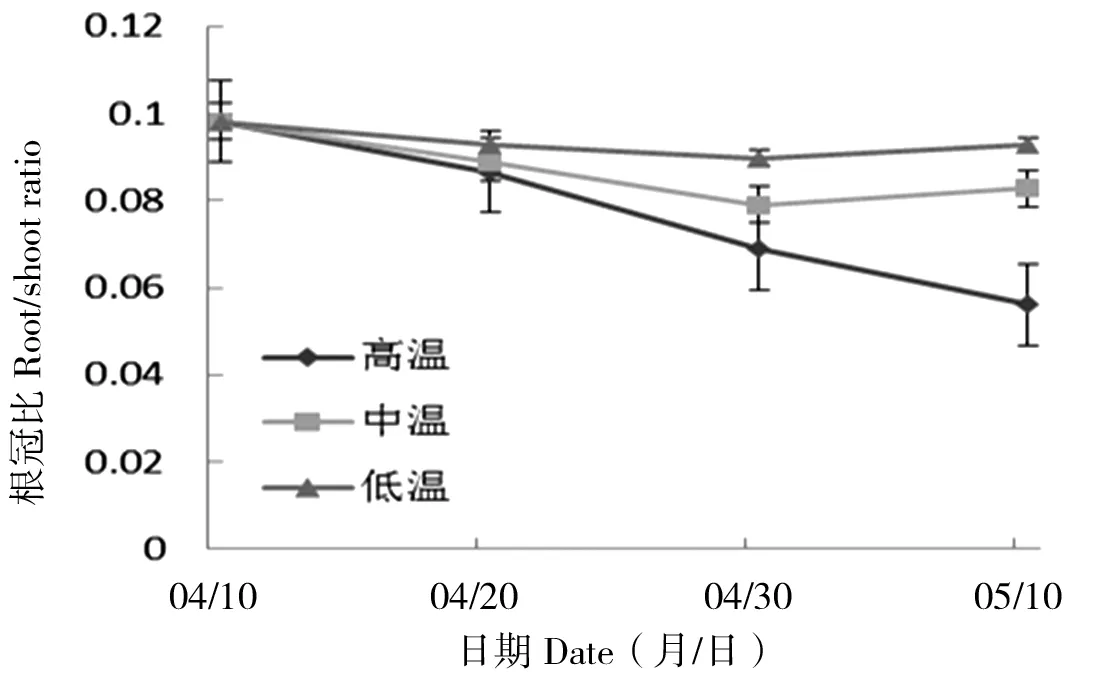
图3 丝瓣剪秋罗幼苗在3种不同温度处理下根冠比随生长时间的动态变化Figure 3 The temporal dynamics of plant root/shoot ratio during thirty days under three temperature treatments
2.1.3 相对生长率可塑性
幼苗相对生长率随温度升高而增大,第10天、第20天及第30天时,高温条件下幼苗相对生长率均大于其他条件下幼苗相对生长率,并且在低温条件下相对生长率最低(图4)。在第10天、第20天和第30天时,丝瓣剪秋罗幼苗的相对生长率在不同温度条件下均存在显著的差异,(P10 d<0.05,P20 d<0.05,P30 d<0.05),温度对于丝瓣剪秋罗幼苗的相对生长率影响非常显著。
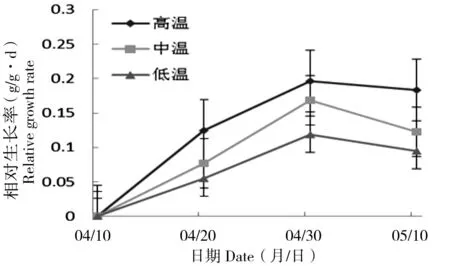
图4 丝瓣剪秋罗幼苗在3种不同温度处理下相对生长率随生长时间的动态变化Figure 4 The temporal dynamics of plant relative growth rate during thirty days under three temperature treatments
2.2 不同土壤养分条件下丝瓣剪秋罗幼苗表型可塑性
2.2.1 植株高度可塑性
在10 d、20 d和30 d时幼苗株高随土壤养分浓度升高而增加;高养分条件下幼苗平均株高均比其他条件下高,并且低养分条件下株高最低(图5)。在不同土壤养分条件下,丝瓣剪秋罗的幼苗株高在第10天、第20天和第30天时,均存在显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05),土壤养分对丝瓣剪秋罗幼苗的株高影响显著。
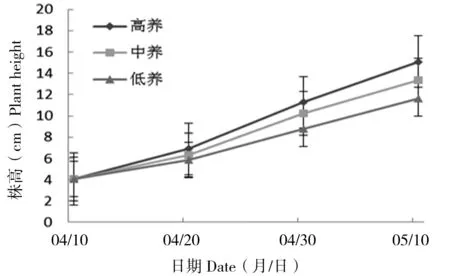
图5 丝瓣剪秋罗幼苗在3种不同土壤养分条件下植株高度随生长时间的动态变化Figure 5 The temporal dynamics of plant height during thirty days under three nutrient treatments
2.2.2 生物量积累和根冠比可塑性
在盆栽实验进行后的第10天、第20天和第30天时观测到丝瓣剪秋罗幼苗总生物量随土壤养分浓度升高而增加的趋势;高养分条件下幼苗总生物量均较其他条件下增加显著,并且低养分条件下总生物量增加最小(图6);不同养分条件下,其幼苗总生物量均存在显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05)。
丝瓣剪秋罗幼苗的根冠比随土壤养分浓度升高,有降低的趋势。第10天、第20天和第30天时,高养分条件下幼苗根冠比较其他条件下小,并且在低养分条件下根冠比下降最为缓慢,根冠比最高(图7)。在3个时间点的不同土壤养分条件下,幼苗根冠比均存在显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05)。

图6 丝瓣剪秋罗幼苗在3种不同土壤养分条件下植株总生物量随生长时间的动态变化Figure 6 The temporal dynamics of plant biomass during thirty days under three nutrient treatments
2.2.3 相对生长率可塑性
丝瓣剪秋罗幼苗相对生长率随土壤养分浓度升高而增加。第10天和第30天时,高养分条件下幼苗相对生长率均大于其他条件下幼苗相对生长率;第20天时,相对生长率最高的是高养分处理,相对生长率最低的是低养分处理(图8)。幼苗的相对生长率在3个时间点不同土壤养分条件下,均存在显著差异(P10 d<0.05,P20 d<0.05,P30 d<0.05)。
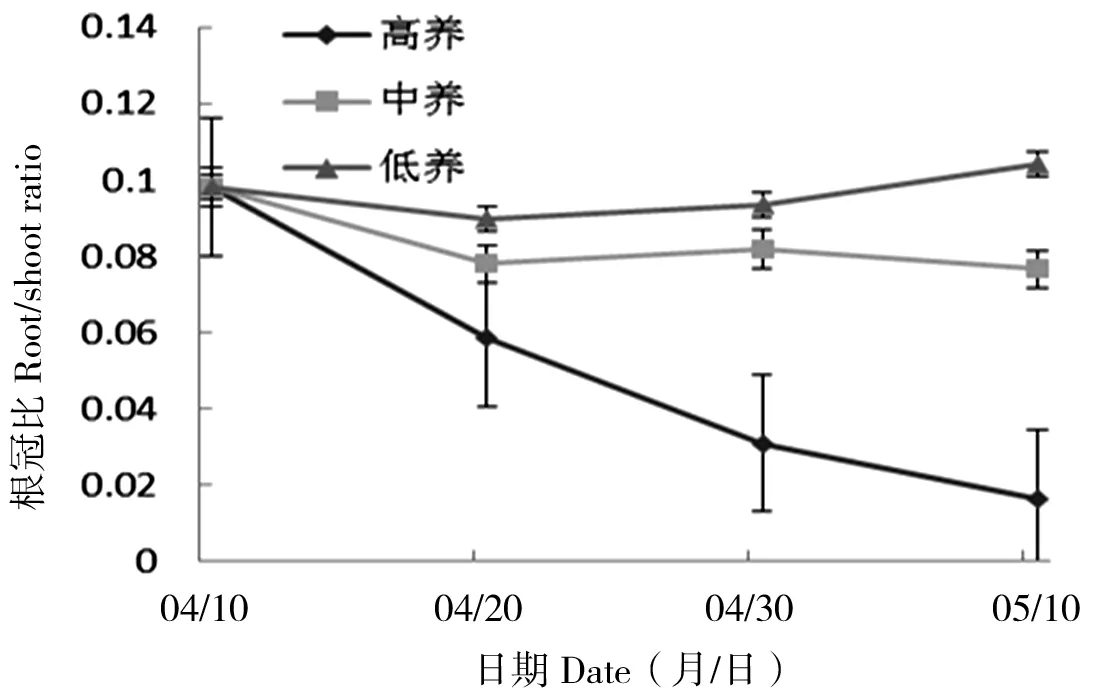
图7 丝瓣剪秋罗幼苗在3种不同土壤养分条件下根冠比随生长时间的动态变化Figure 7 The temporal dynamics of plant root/shoot ratio during thirty days under three nutrient treatments
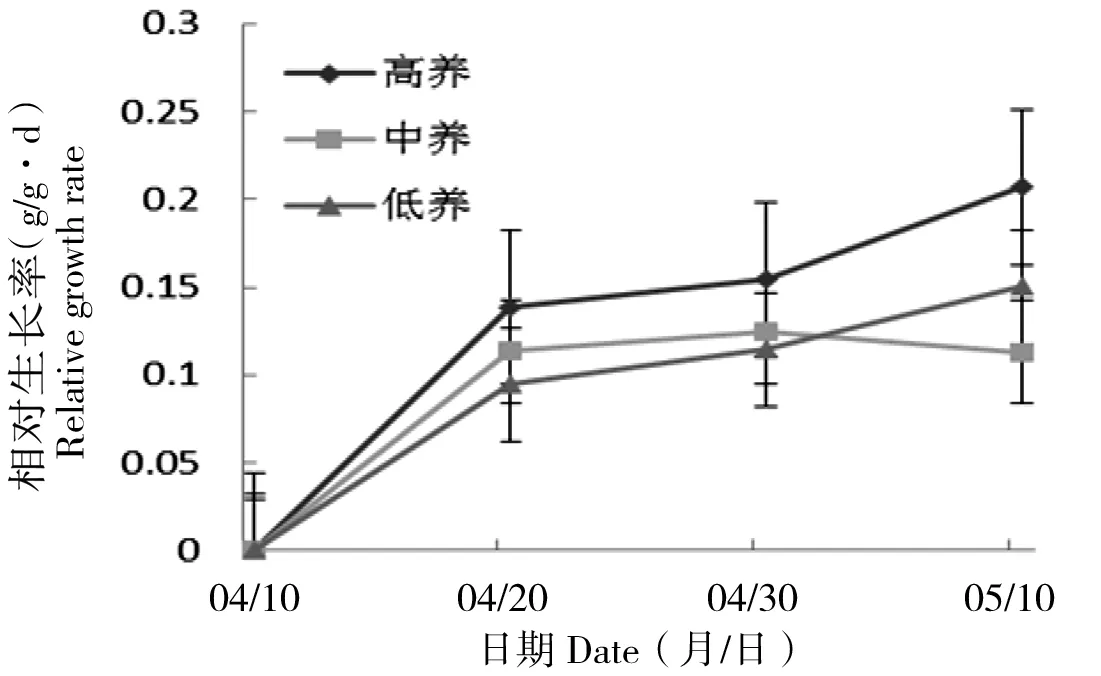
图8 丝瓣剪秋罗幼苗在3种不同土壤养分条件下相对生长率随生长时间的动态变化Figure 8 The temporal dynamics of plant relative growth rate during thirty days under three nutrient treatments
2.3 丝瓣剪秋罗幼苗的表型可塑性指数
在不同温度处理下,第10天时,相对生长率可塑性最强;第20天和第30天时,总生物量可塑性最强;分别取各项参数的表型可塑性指数均值表明:相对生长率可塑性最强,其次是总生物量可塑性与株高可塑性,可塑性最低的为根冠比可塑性(表1)。在不同土壤养分处理条件下,其中根冠比可塑性最强,其次是相对生长率可塑性与总生物量可塑性,可塑性指数最低的为株高(表2)。PI(温度)=0.5004>PI(养分)=0.4709,表明丝瓣剪秋罗幼苗对于温度的可塑性更强。
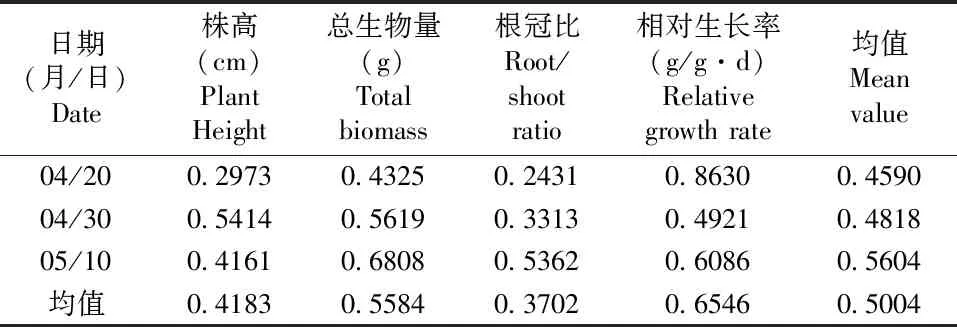
表1 不同温度条件下丝瓣剪秋罗幼苗的表型可塑性指数Table 1 Phenotypic plasticities of seedlings of L. wilfordii under different temperature conditions
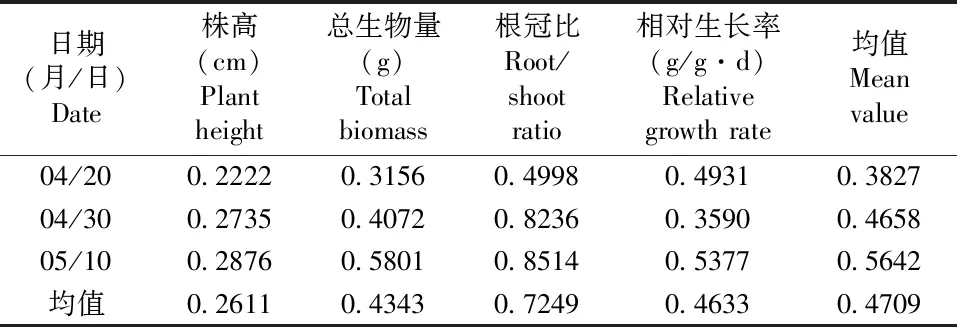
表2 不同土壤养分条件下丝瓣剪秋罗幼苗的表型可塑性指数Table 2 Phenotypic plasticities of seedlings of L. wilfordii under different nutrient conditions
3 讨论
3.1 丝瓣剪秋萝幼苗对温度条件的表型可塑性响应
植物所采取的生活史策略主要是通过能量的合理分配、各个生命过程的协调,使物种的繁殖、存活及适合度达到最大[17]。当环境资源条件改变时,植物幼苗通过形态特征的改变来适应生境条件的变化,如幼苗高度、生物量积累和分配以及相对生长率发生改变,以维持幼苗在不同环境下的存活和生长[18]。实验结果表明,在不同温度处理条件下,30℃和20℃处理下的植物幼苗长势都较好,且生长明显快于10℃的处理组。30 d内丝瓣剪秋罗幼苗的株高、总生物量、相对生长率随温度梯度升高而显著增加。温度对植物生长的影响主要是通过调节生化物质含量和保护酶活性来改变各种代谢过程而产生的一种综合效应[19]。10℃处理的丝瓣剪秋萝幼苗株高、总生物量、相对生长率相对其他两组处理较小,研究期内丝瓣剪秋罗幼苗的根冠比呈下降趋势,并且温度越高,根冠比越小。可能是因为在低温条件下,蒸腾作用和呼吸作用减弱等因素限制了水分和养分的吸收,而为了保证根部对水分和养分的吸收,将更多的资源分配给根部,表现为根冠比增大。30℃的处理组各生长参数均高于其他处理组,生长发育均好于其他处理组。范增丽对两种淫羊藿属植物(Epimedium)的研究表明,增温显著影响植株的生长进程,在增温20℃的条件下,两种淫羊蕾属植物的株高,叶片长、宽及叶面积的增量均大于对照和增温5℃[20]。这与对高寒草甸6种常见禾本科植物幼苗生长和生物量分配的影响研究和不同温度下雌雄葎草营养生长期的生长特性等研究结果相似[17,21,24-25]。本研究发现丝瓣剪秋罗幼苗的相对生长率对温度的可塑性响应最强,幼苗相对生长率是植物生活史特征的重要方面,也是评价幼苗生长对环境响应的重要指标[22]。在相似资源条件下, 具有较大相对生长率可塑性的草本植物种具有较大的入侵性能和拓殖能力[23],丝瓣剪秋萝野外居群生长在湿地中,推测其幼苗相对生长率可塑性最强,可能是其居群更新、成功定植的决定因素。
3.2 丝瓣剪秋萝幼苗对土壤养分条件的表型可塑性响应
在不同土壤养分处理下,丝瓣剪秋罗幼苗的高度、总生物量、相对生长率随土壤养分梯度升高而增加,在一定范围内土壤养分越高,高度、总生物量、相对生长率越大。研究期内丝瓣剪秋罗幼苗的根冠比随幼苗生长发育的进行逐渐降低,丝瓣剪秋萝幼苗在低养分时的根冠比最大,在高养分时根冠比最小,符合平衡生长假说,该假说认为植物在资源受限时会把资源分配给能更多获取受限资源的器官[26]。有很多研究结果表明增加养分将会使根生物量减小,叶生物量增大[27],养分充足时,幼苗把更多的资源用于地上部分各器官的生长,以利于叶片等增加从而获取更多的光能用于光合作用。而在低养分时,幼苗具有较多的根生物量,用于吸收更多的水分和养分,提高生存能力[18]。本研究中,养分较缺乏的低养分组,幼苗把资源更多地分配给根部以满足自身最大限度地获取养分,导致根冠比较高养分组增大。
4 结论
丝瓣剪秋罗幼苗对温度变化的表型可塑性指数高于对土壤养分变化的表型可塑性指数,意味着丝瓣剪秋萝幼苗时期对温度条件的变化较为敏感,对温度条件变化表现为积极的可塑性响应。植物在异质环境中,不论表现为消极的可塑性响应还是积极的可塑性响应,表型可塑性能够使植物改变表型特征,最大限度地提高自身适合环境的适合度。本研究中丝瓣剪秋萝幼苗在高温和中温处理下较低温处理各项参数显著升高,在高养分条件下的各项生长参数也优于中低养分条件。根据实验结果,丝瓣剪秋萝幼苗阶段生长发育最适温度为20℃~30℃。日后可以通过调控土壤中养分浓度和外界温度,来提高丝瓣剪秋萝幼苗对湿地复杂环境的适应能力,进而可将其应用于湿地的生态保育与恢复。
