细条天竺鲷群体线粒体DNA控制区序列比较分析❋
2019-04-16方润东周永东李鹏飞高天翔
方润东, 宋 娜, 周永东, 李鹏飞, 高天翔
(1. 中国海洋大学水产学院,山东 青岛 266003; 2. 浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江 舟山 316021; 3. 浙江省海洋水产研究所,浙江 舟山 316021; 4. 浙江海洋大学水产学院,浙江 舟山 316021)
细条天竺鲷(Apogonlineatus)属鲈形目(Perciformes)、天竺鲷科(Apogonidae)、天竺鲷属(Apogon),主要分布于中国的黄渤海和东海,台湾及南海亦有分布[1-4],为底栖性小型鱼类,多栖息于泥砂质海底,为常见的小型鱼类,是一些食肉型经济种类的重要食物来源。细条天竺鲷常栖息于近岸中下层,磷虾、十足类和端足类是其主要摄食的对象[5]。由于细条天竺鲷很少被食用、没有任何直接的经济价值,和众多小型鱼类一样,并未受到足够重视。小型鱼类既是食肉型经济鱼种捕食的对象,又可以摄食其他鱼类产的卵,可以在一定程度上调控某些经济鱼类的种群规模[6]。因此随着当前渔获物组成的明显改变,这类小鱼对于维持海洋生态系统食物链正常有序的能量流动和物质交换具有不可替代的重要作用[6-8]。因此,开展对细条天竺鲷种质资源状况及现有遗传丰富度的调查研究,能够正确了解并有效保护这一重要的海洋鱼类。迄今为止,有关细条天竺鲷的研究仅仅局限于耳石形态、食性、资源分布及生态价值等基础工作[9-13]。虽然细条天竺鲷具有较高的生态价值,但尚未见其群体遗传学方面的研究报道。
为了实现渔业资源开发的可持续性,需要建立有效的渔业管理制度。根据群体建立不同的单元可提高渔业管理效率,而群体遗传结构对渔业管理单元的划分具有关键作用[14]。线粒体DNA(Mitochondrial DNA, mtDNA)作为分子标记对检测群体遗传结构非常敏感,而且线粒体DNA呈母性遗传且不发生重组,其中控制区进化速度较快,是研究遗传多样性和群体遗传结构较为理想的工具[15-17]。目前利用mtDNA控制区基因检测海洋生物遗传多样度组成及群体遗传特征已成为显著有效的方法,曾先后被应用于日本鳀(Engraulisjaponicus)、斑尾复虾虎鱼(Synechogobiusommaturus)、香鱼(Plecoglossusaltivelis)[18-20]等海水鱼的遗传丰富度和遗传特征的探讨。除此以外,核苷酸序列中的变异个数和分布方式能帮助我们推测鱼类种群的进化历程,进而探讨产生和维持这些多态的机制[19]。进入更新世冰期后,全球剧烈的气候波动导致海平面下降,海洋环境变化明显[21]。海洋鱼类适宜的栖息环境大量减少导致种群数量急剧下降,幸存下来的个体集中到避难所。冰期过后,气温的回升使海平面回升到之前高度,幸存者在度过冰期后由原有避难所重新移殖到其他适宜生存的环境中[22]。由此可认为冰期气候波动对海洋环境的改变深刻影响到海洋鱼类的群体遗传特征[23]。本研究利用mtDNA控制区序列检测细条天竺鲷群体的遗传多样性及遗传结构,探讨影响其种群遗传结构形成和变化的因素,从而为保护细条天竺鲷现有生物资源及适度的开发利用提供可靠的理论依据。
1 材料与方法
1.1 实验样品
本研究所用的细条天竺鲷群体分别采自威海乳山、青岛胶南、上海以及舟山近海海域,共114尾细条天竺鲷个体,基本覆盖了中国沿海细条天竺鲷的主要分布范围(黄渤海到东海,见表1、图1)。物种的鉴定依据Mabuchi等[4]。
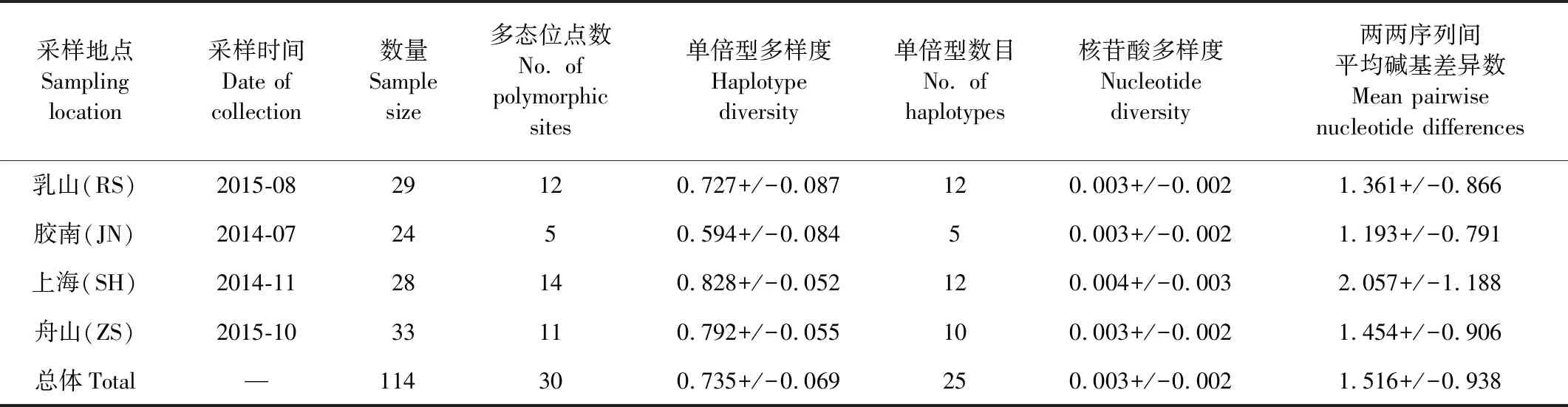
表1 细条天竺鲷的采样信息以及一些遗传多样性指数Table 1 Sample information of Apogon lineatus including several genetic diversity indices
图1 细条天竺鲷的采样地点Fig.1 Sampling location of Apogon lineatus
1.2 DNA提取与目的片段扩增
剪取样本一小块背部肌肉,并采用标准的苯酚-氯仿法提取基因组DNA。扩增所用引物为控制区通用引物[19],正向序列为5’-CCCACCACTAACTCCCAAAGC-3’,反向序列为5’-CTGGAAAGAACG-CCCGGCATG-3’。
PCR反应在体系包括: 1 μL DNA模板,17.25 μL去离子水,2.5 μL 10×PCR buffer,2 μL的dNTPs,0.15 μL Taq聚合酶,正反向引物物各1 μL。热反应循环为:94 ℃预变性5 min,94 ℃变性45 s,50 ℃退火45 s,72 ℃延伸1 min, 共35次循环;72 ℃再延伸10 min。为防止样品污染,每次试验均设置阴性对照。将纯化、回收后的PCR产物送到上海桑尼生物技术有限公司,采用ABIPrism3700自动测序仪双向测通产物序列。
1.3 数据处理
使用 Dnastar 软件包下的Seqman软件对测序结果进行比对分析并加以校正;遗传多样性指数计算在ARLEQUIN 3.11软件上完成[24]。遗传多样性指数包括:单倍型多样度(Haplotype diversity),群体多态位点数(No of polymorphic sites),核苷酸多样度(Nucleotide diversity)和平均两两碱基差异数(Mean pairwise nucleotide differences)。计算单倍型之间的关系和核苷酸之间的差异数,并手工构建分支网络树。以斑鳍天竺鲷(Apogoncarinatus)为外群,使用Mega3.0软件基于K2P模型构建系统树。
使用群体间分化指数Fst值衡量两两群体间遗传分化程度[25]。在计算两两群体间Fst值时,进行10 000次重抽样检测其显著性。当进行多重比较时,在Excel中使用Bonferroni方法校准P值[26]。采用确切P检验,检查不同群体间能否发生随机交配。
核苷酸不配对分布分析和中性检验用来检测细条天竺鲷群体历史动态。中性检验包括Tajima’sD检验[27]和Fu’Fs检验[28]。D值和Fs为显著的负值,通常表明群体经历过扩张事件。核苷酸不配对分布分析同样用来检测是否存在群体扩张[29]。使用广义非线性最小方差法估算群体扩张时间τ值和群体扩张参数θ0,θ1。τ值是用突变单位表示的群体扩张的时间。利用公式τ=2ut可把τ值换算成真实的扩张时间,其中t为群体扩张实际经历的时间,u为整个基因片段的变异速度。θ0代表扩张前的群体扩张参数 ,θ1反之。
2 结果
2.1 遗传多样性
细条天竺鲷所有个体经PCR扩增后获得长504 bp的目的片段。对所有测序得到的原始序列进行比对校正、剔除转运RNA后,获得4个群体共114个个体长度为476 bp的控制区第一高变区序列,作为用于后续数据分析的目的序列。在所有目的序列中发现10处简约信息位点和26处变异位点,其中转换23处,颠换3处。4种碱基在目的序列中占比分别为:A为32.39%,C为18.62%,G为17.19%,T为31.80%, A+T占比(64.19%)高出C+G占比(35.81%)很多,表现为AT偏好。细条天竺鲷控制区A+T含量较高,与其他海洋鱼类控制区A+T占比高,C+G占比低的特征类似。除此之外,114条控制区序列共包含了30个单倍型,其中仅有4个单倍型被不同群体共享,其余26个均为各群体所独有(见表2)。综合单倍型多样度及核苷酸多样度来看,上海和舟山群体的遗传多样性高于乳山和胶南群体的遗传多样性。
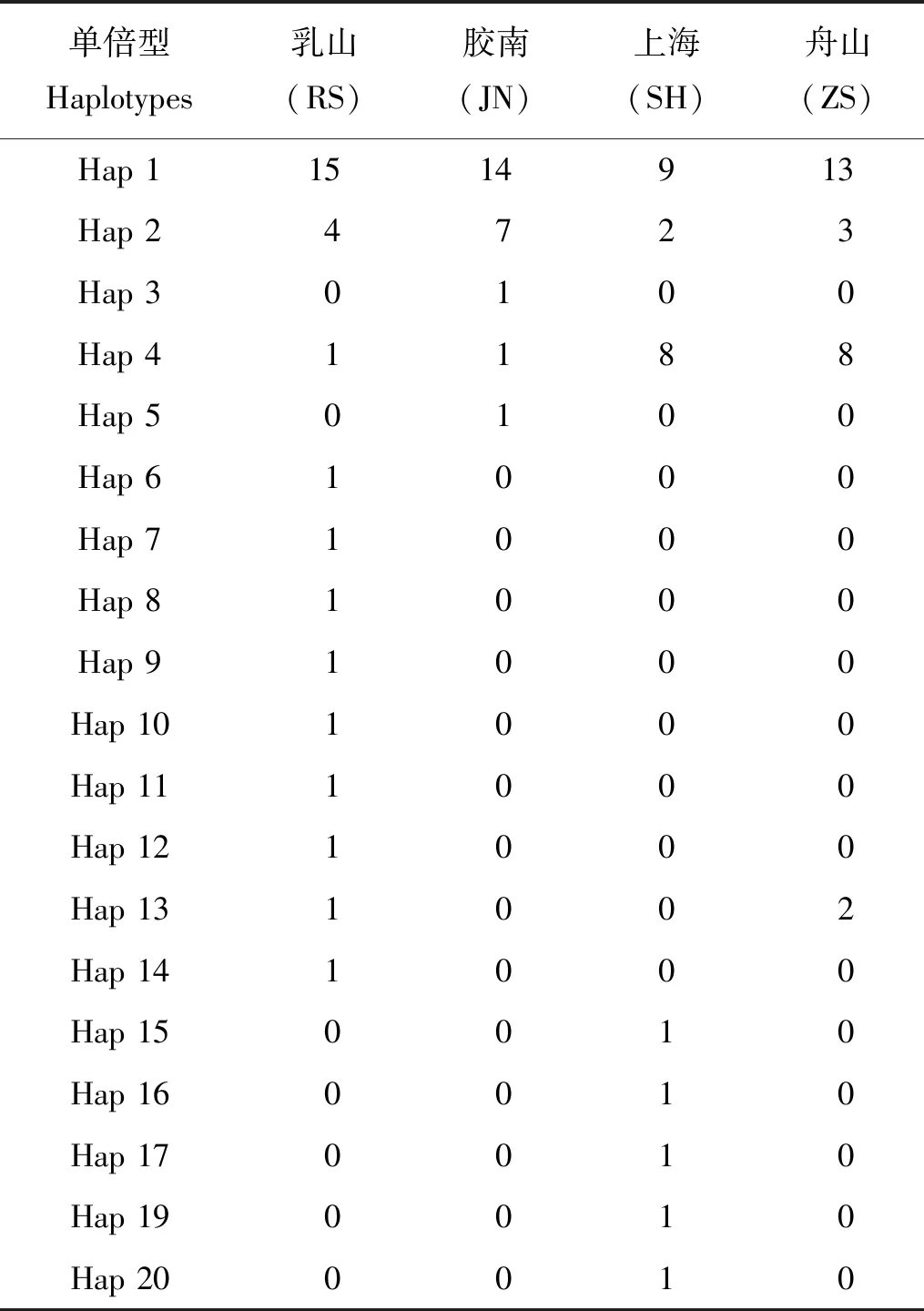
表2 细条天竺鲷4个地理群体的单倍型分布Table 2 Haplotypes in four Apogon lineatus populations
续表2
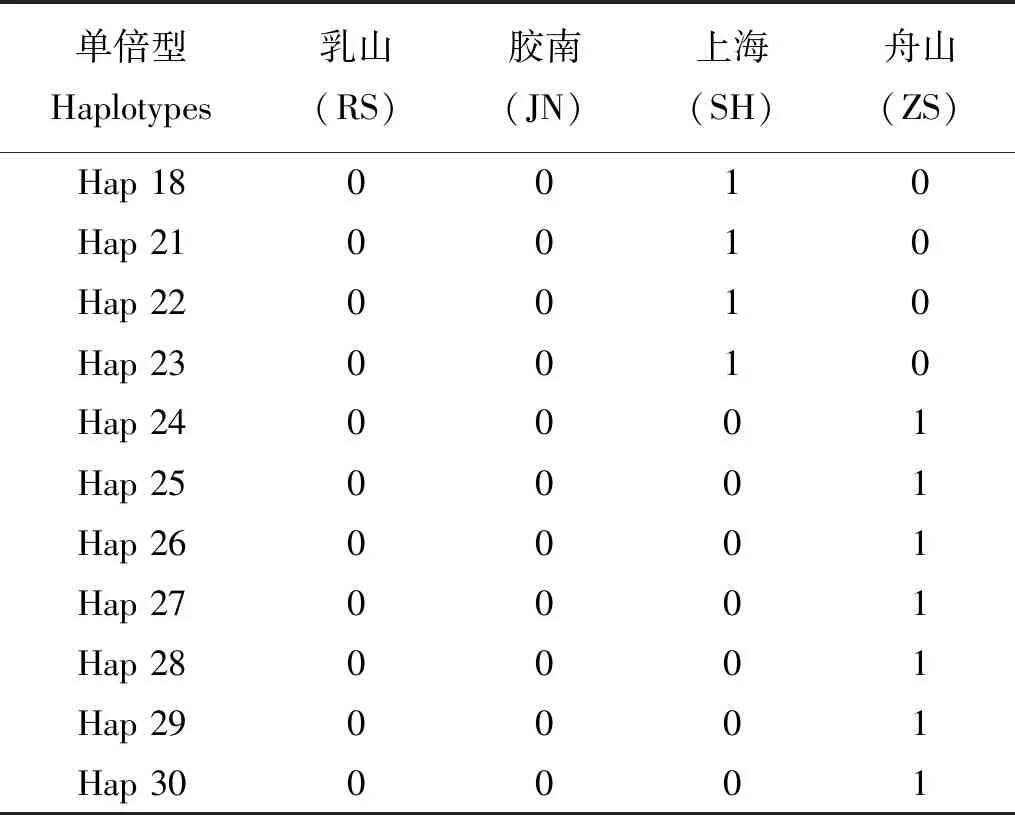
单倍型Haplotypes乳山(RS)胶南(JN)上海(SH)舟山(ZS)Hap 180010Hap 210010Hap 220010Hap 230010Hap 240001Hap 250001Hap 260001Hap 270001Hap 280001Hap 290001Hap 300001
2.2 群体遗传结构
以斑鳍天竺鲷为外群,利用30个细条天竺鲷控制区单倍型构造邻接关系树(Neighbor-joining tree,NJ树),结果表明不同群体的单倍型均无规律地散布到单倍型NJ树上。不存在与地理群体显著对应的系统分支,由此我们认为细条天竺鲷不同地理群体间并无显著的谱系结构。细条天竺鲷与斑鳍天竺鲷遗传差异显著(见图2)。

(比例尺为0.01,代表遗传距离远近,分支处数字为超过50%的自展支持率。Scale bar=0.01 substitutios per site. Numbers at notes are bootstrap values.The numbers lower than 50% are not shown.)
图2 基于细条天竺鲷30个控制区单倍型构建的邻接关系树, 所用外群为斑鳍天竺鲷A.carinatus
Fig.2 NJ tree forA.lineatusbased on 30 haplotypes with an outgroupA.carinatus
由30个单倍型构成的分支网络树(见图3),其分支结构与NJ树的构造相类似,没有发现某一单倍型分支都来自同一地理群体的现象。分支网络树的构造为星爆形,由此可推测细条天竺鲷群体历史上曾发生扩张事件。
细条天竺鲷两两群体间Fst值的范围为-0.015到0.067(见表3),并且不同群体间遗传差异都不显著(经校正的P值大于0.05),同时确切P检验的分析提示随机交配现象广泛存在于这4个群体之间。

图3 细条天竺鲷控制区单倍型分支网络树Fig.3 Minimum spanning trees showing genetics relationship in A. lineatus

乳山(RS)胶南(JN)上海(SH)舟山(ZS)乳山(RS)—0.8660.0200.117胶南(JN)0.017—0.0110.045上海(SH)0.0340.067—0.607舟山(ZS)0.0180.062-0.015—
2.3 群体历史动态
较低的核苷酸多样度和单倍型分支网络树的星爆形构造,暗示了细条天竺鲷群体历史上可能发生过扩张事件。核苷酸不配对分布分析的结果(见图4)表明,细条天竺鲷核苷酸不配对分布表现为近正态分布单峰形,同时观测值的分布与群体扩张模型预期分布较好地重合。利用中性检验对这一推测进行验证(见表4),Tajima’sD值和FS值均小于0,并且所有群体的统计检验显著(P<0.05)。因此综合以上两种方法的验证,我们认为细条天竺鲷群体历史上发生过扩张事件。
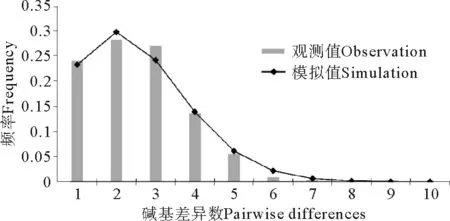
图4 细条天竺鲷控制区单倍型核苷酸不配对分布Fig.4 Mismatch distributions of control region haplotypes of A. lineatus
τ值(图4中的峰值)是用突变单位表示的群体扩张的时间,利用公式τ=2ut可换算成真实的扩张时间t,其中u为整个基因片段的变异速度。本研究根据5%~10%/MY的控制区分歧速率和τ值2.973,估算出细条天竺鲷的群体扩张时间大约为62 450~124 900 a前,处于更新世晚期。
3 讨论
具有较高遗传多样性的物种,生存能力和对环境变化的适应能力越强[30]。对物种遗传多样性的研究可以帮助预测物种的发展动向和分析物种的进化潜力。
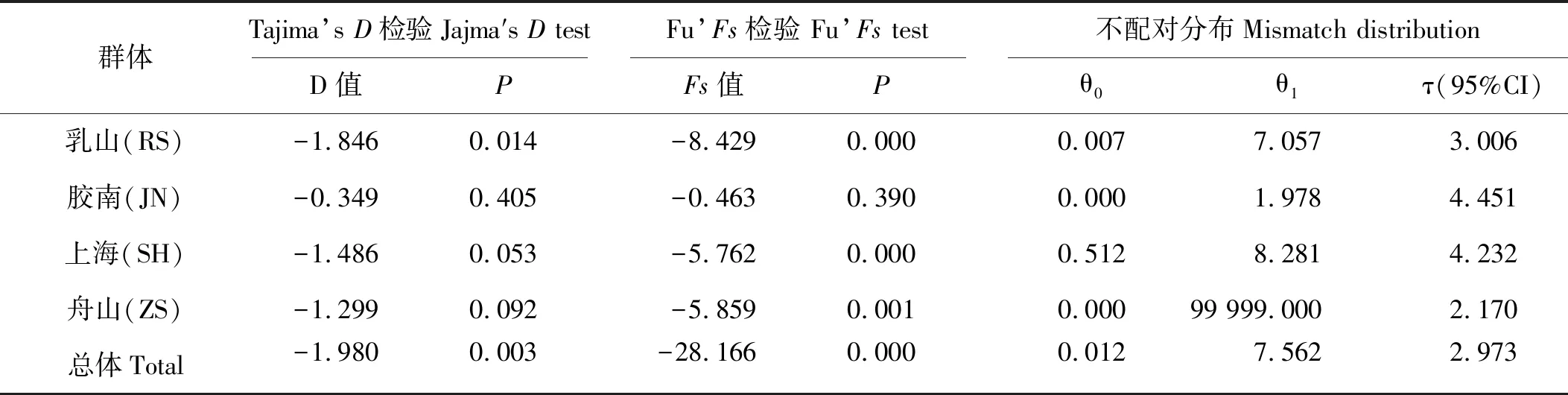
表4 细条天竺鲷群体Tajima’s D、Fu’Fs统计值和核苷酸不配对分布分析结果Table 4 Tajima’s D, Fu’Fs and mismatch distribution parameter of A. lineatus
本研究中,细条天竺鲷作为其分布区主要小型鱼类优势种和肉食性经济鱼类的重要饵料鱼种,与已研究报道的鱼类相比其遗传多样性属于中等偏低的水平[19-20]。鉴于高有效种群尺度是维持物种高水平遗传多样性的原因[31],有效种群较小是造成其低遗传多样性的重要原因,细条天竺鲷资源量可能较少,因此其资源状况需要引起渔业资源管理部门的重视。
海洋生态系统由于其开放性缺乏明显的地理和物理障碍,并且广泛存在的洋流运动能够使鱼卵、幼体和成鱼扩散到更远的范围。海洋鱼类群体间的基因交流一般不会因为地理距离增大而中断,这保证了不同地理群体仍有相似的遗传结构[31-34]。Fst值又称群体分化指数,用来描述群体间遗传差异大小,差异越大则该值越大,反之越小。细条天竺鲷4个群体的遗传分化指数Fst值介于-0.015~0.067之间,同时统计检验不显著,因此可以判断不同地理群体间遗传结构非常相近。本研究通过对细条天竺鲷线粒体DNA控制区序列分析,表明细条天竺鲷分布在我国黄海和东海的4个群体间没有明显的地理分化,可能是由一个大而稳定的种群经过长期进化而来。出现这种情况的原因可能是:乳山海域到舟山海域之间不存在影响细条天竺鲷基因交流的明显地理障碍,虽然细条天竺鲷成体不作长距离迁移,但是其漂浮卵、浮游幼体和成体都可能因为中国沿岸流活动而扩散较远的距离[21,35]。我国近海的黑鲷(Acanthopagrusschlegeli)和台湾海峡的斑鰶(Konosiruspunctatus)等也被证实在较大区域内没有明显的地理分化[36-37]。
更新世全球气候波动表现为冰期和间冰期相互更迭。以大概10万a为一次间隔,这样的气候更替持续了80万a,海平面也随之发生升降[38]。第四纪末冰盛期,海平面下降了120~140 m,细条天竺鲷的群体数量和分布受到了巨大影响,适宜其栖息的环境大量消失导致种群数量急剧下降,幸存下来的个体集中在避难所。冰期过后,气温的回升使海平面回升到之前高度,细条天竺鲷在度过冰期后由原有避难所重新移殖到其他适宜生存的环境中,之前孤立的种群也随着隔离海域的重新连接开始逐渐交流[39-40]。细条天竺鲷上海、舟山群体的单倍型多样度及核苷酸多样度显著高于乳山、胶南群体,我们推测可能是因为中国东海海盆是其在更新世冰期的避难所之一[18]。核苷酸不配对分布在经历过近期群体扩张事件的群体中一般会呈现明显的单峰分布[29,41]。核苷酸不配对分布的结果提示细条天竺鲷发生了群体扩张,Tajima’sD值和FS值均小于0,并且所有群体的统计检验显著(P<0.05),亦验证了这一推测。我们将5%~10%/MY的控制区分歧速率用于细条天竺鲷控制区,得出其群体扩张事件大约发生在在62 450~124 900年前,属于晚更新世冰期。由此可以推测细条天竺鲷的群体扩张事件与更新世气候的波动有关。
本研究涉及的地理群体只有乳山、胶南、上海和舟山共4个群体,而鉴于细条天竺鲷在我国是分布于黄渤海及东海的常见小型鱼类,因此想要探讨细条天竺鲷在中国沿海分布范围内的遗传结构我们还需要分析更多的群体以及样本并利用包括核基因等分子标记方法,以期更好地了解这一物种的遗传分化,为评估其资源状况及在海洋生态系统中的价值提供科学依据。
