东祁连山金露梅灌丛不同恢复期碳储量和土壤养分变化
2019-04-04徐长林宋美娟汪鹏斌鱼小军
贺 翔,徐长林,宋美娟,汪鹏斌,鱼小军
(甘肃农业大学草业学院 / 草业生态系统教育部重点实验室 / 甘肃省草业工程实验室 /中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
金露梅(Potentilla parvifolia)灌丛是高寒落叶灌丛的典型代表,广泛分布于青藏高原海拔2 700-4 500 m的山地阴坡、半阳坡、潮湿滩地及高海拔的山地阳坡[1]。在青藏高原以及我国其他高大山系中约有金露梅灌丛 1.16 × 107hm2,分布面积居世界之最,仅次于矮生嵩草(Kobresia humilis)草甸面积[2]。和其他灌丛一样,金露梅灌丛含有丰富的植物资源,同时也是野生动物的栖息地以及家畜的放牧地,并且具有良好的持水能力[3],对水土保持及涵养水源[4]具有重要的生态意义。
随着全球气候变暖以及人类生产生活的改变,灌丛生态系统也发生了较多变化。近年来针对金露梅灌丛的分布[5]、生物量[6-7]、CO2排放[8]及放牧影响[9-10]等方面已有研究。土壤养分是反映草地生产性能及土壤潜在肥力的重要指标,也是制定土壤改良利用方向及措施的重要依据[11]。关于土壤氮、磷、钾元素的研究主要集中在植被结构及其地上-地下生物量分配[12]、大区域或农田生态系统[13]、退化草地[14]、不同植被类型[15]等方面。目前,诸多学者对灌丛植被特征及变化有较多研究,而有关金露梅灌丛土壤理化性质及生态功能的动态研究尚少。因此,为明晰植被恢复过程中土壤特性的变化特征,以位于东祁连山的甘肃天祝高寒金露梅灌丛为研究对象,对比了不同恢复期金露梅灌丛碳储量和土壤养分含量变化情况,为高寒灌丛生态系统管理提供科学依据。
1 材料与方法
1.1 样地概况
试验地设在甘肃省天祝县抓喜秀龙乡附近的金露梅灌丛,地理坐标为 37°11′-37°13′ N,102°23′-102°29′ E,海拔 2 960 m。该地区水热同期,无绝对无霜期,年均气温-0.1 ℃,最热月7月,均温为 12.7 ℃,最冷月 1 月,均温为-18.3 ℃,≥ 0 ℃的年积温为 1 380 ℃·d。年均降水量 416 mm,多集中于7-9月,年蒸发量为1 592 mm。土壤以亚高山草甸土、亚高山黑钙土等为主[16]。灌丛土层薄,约 25 cm,土壤 pH 为 7.0~8.2[11]。
1.2 试验设计
运用“空间代替时间”的方法[17],依据前人的研究基础,选择1997年监测的金露梅灌丛样地相关数据为未恢复治理期 (no recovering, NR),2018 年7月在1997年的原样地(此样地20年一直处于自然演替)附近再次监测,所得数据代表恢复后期(late stages of restoration, LR)。依据任继周的《草业科学研究方法》[18],对周边金露梅灌丛群落植被状况进行调查,依调查结果在微地貌及生境条件基本一致的灌丛群落选择布设2个100m×100m样地,代表1997-2018年20年期间的恢复初期(initial stages of restoration, IR)和恢复中期 (medium-term of restoration, MR)。各样地植被基本情况如表1所列。
1.3 测定方法
野外采样:于2018年8月底分别在各样地内的10m×10m样方中随机挖取金露梅灌丛5丛,分别装袋带回实验室,按叶、枝、茎、根分别收集,并在灌丛下取0.5m×0.5m样方测定草本,重复3次,收集其地上、地下部分植物量,装袋带回实验室,且根系泥土用流水清洗干净,然后将所有样品置入电热恒温鼓风干燥箱于105 ℃杀青30min,再于 65 ℃下烘干 24 h后称重,粉碎,过 0.15 mm网筛,装入棕色广口瓶以用于碳含量的测定。

表1 试验样地灌丛植被基本情况Table 1 Shrub vegetation situations in test sampling plots
2018年8月初,分别在3个样地的对角线附近均匀选择3个5 m × 5 m样方,在样方中选择具有代表性观测点一处,即每个样地3个点,在测定前一天,将PVC环嵌入土壤中,其嵌入深度为2 cm,清除环内的植物和凋落物,经24 h平衡后,采用土壤碳通量自动测量系统(Li-8100)进行土壤呼吸测定。选择天气晴朗、气候条件稳定的时间,测定时间一般为08:00-12:00,每个点测定时间约为2 min,所有测定点必须在 4 h 内完成。连续测定 8 d,每月共测两次。为了消除日动态变化的影响,每次按不同顺序进行测定。
室内分析:2018年8月中旬采用五点取样法进行土壤取样,利用直径3.5 cm土钻,取0-10、10-20和20-30 cm土层土壤,5次重复,取400~500 g土样带回实验室,捡出土壤中碎石子、草根等,阴干后研磨细,过0.25 mm筛网,测定土壤有机质(重铬酸钾-外加热法,g·kg-1)、全氮(半微量凯氏定氮法,g·kg-1)、全磷(H2SO4-HClO4消煮-钼锑抗比色法,g·kg-1)、全钾(NaOH熔融-火焰光度法,g·kg-1);过1 mm筛网,测定速效氮(碱解扩散法,mg·kg-1)、速效磷(高锰酸钾氧化-葡萄糖还原法,mg·kg-1)、速效钾(NH4OAc浸提-火焰光度法,mg·kg-1)[20]。
土壤容重采用环刀法测定,在每个样地边界处挖掘一个土壤剖面,用直径5 cm土壤环刀按0-10、10-20和20-30 cm土层取土壤容重样品,5次重复,带回实验室,置入电热恒温鼓风干燥箱于105 ℃烘干24 h,称重并计算土壤容重。
1.4 统计分析
利用 Microsoft Excel 2010 进行数据整理,并绘图,利用 SPSS 20.0 对所有数值以“平均值 ± 标准误”进行单因素方差分析。
2 结果
2.1 金露梅灌丛碳储量
随着金露梅灌丛恢复期的延长,金露梅的叶、枝、茎和根的碳储量呈增大趋势,各恢复期间差异显著(P<0.05);金露梅灌丛下草本地上部分的碳储量基本保持不变,差异不显著(P>0.05);草本地下部分的碳储量随着恢复期延长呈下降趋势,各恢复期间差异显著(P<0.05);整个灌丛系统的碳储量呈增大趋势(图1)。
2.2 土壤物理性质变化
2.2.1 土壤呼吸
金露梅灌丛丛间土壤呼吸速率随恢复期延长总体呈升高趋势,NR、IR、MR间和 IR、MR、LR间无显著差异(P>0.05),但LR显著高于NR(P<0.05);丛内土壤呼吸速率呈先降后升趋势,NR 显著高于 IR、MR、LR(P< 0.05)(图 2)。
2.2.2 土壤水分
金露梅灌丛经20年恢复,土壤含水量也表现出不同程度的差异,其丛间土壤含水量,0-10 cm土层中,NR、IR、MR 之间无显著差异 (P>0.05),三者均显著低于 LR(P<0.05);10-20cm 土层中,MR和LR无显著差异(P>0.05),均显著高于NR和IR(P<0.05);20-30cm 土层中,IR、MR、LR 无显著差异 (P>0.05),MR 和 LR 显著高于 NR(P< 0.05)(图3)。金露梅灌丛丛内土壤含水量变化趋势与丛间土壤含水量变化基本一致,总体均呈上升趋势 (图 3)。
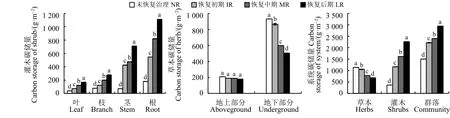
图 1 金露梅灌丛碳储量动态变化Figure 1 Dynamic changes in the carbon storage of Potentilla parvifolia
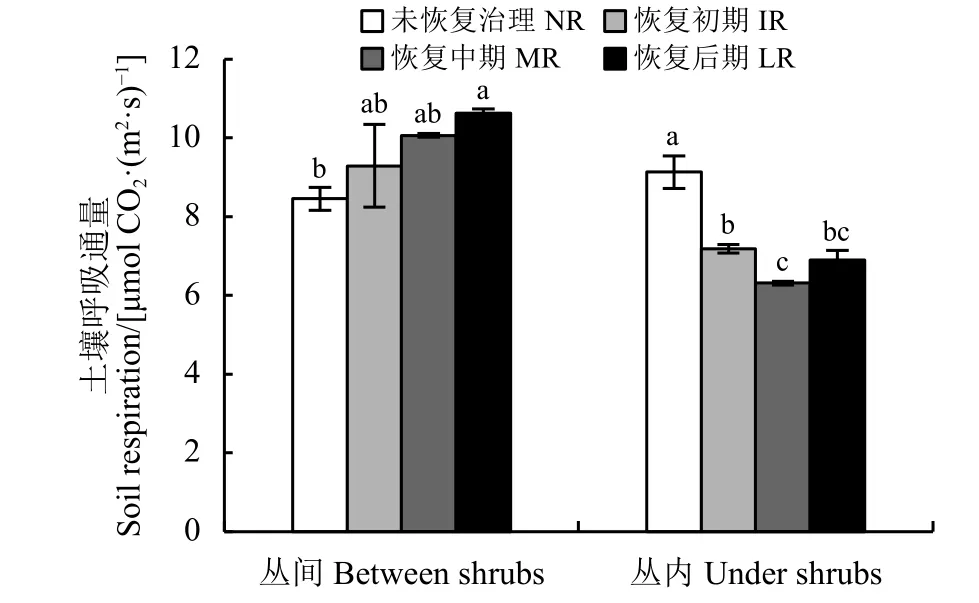
图 2 金露梅灌丛土壤呼吸动态变化Figure 2 Dynamic changes in the soil respiration of Potentilla parvifolia
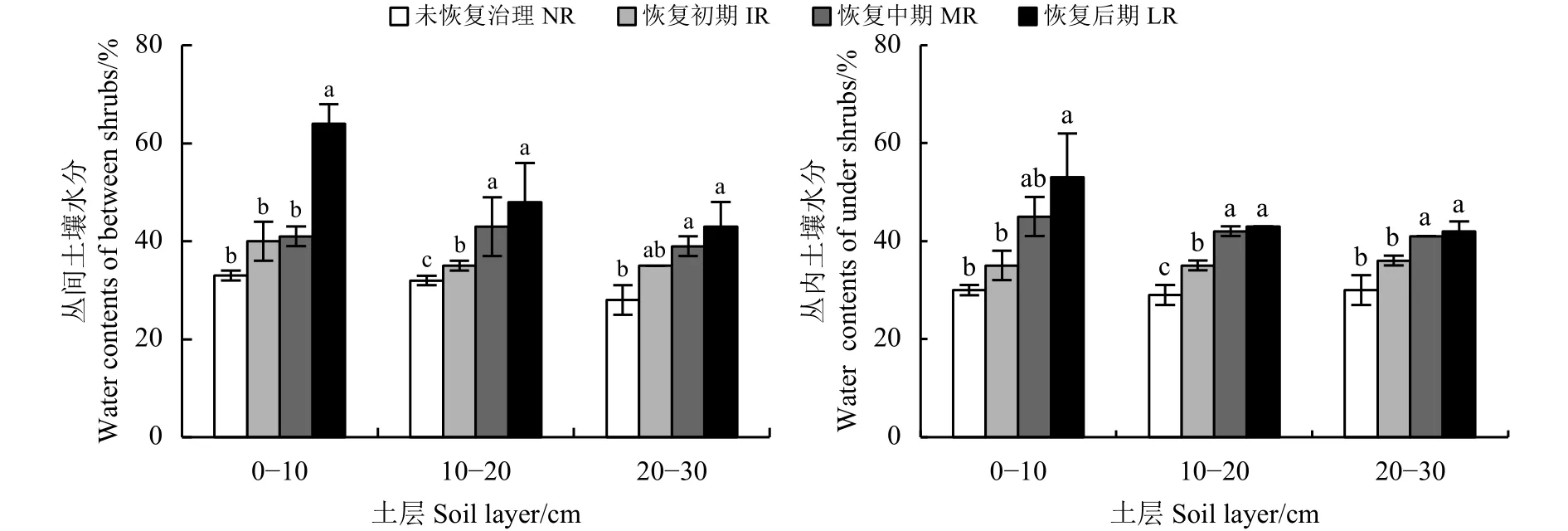
图 3 金露梅灌丛土壤含水量动态变化Figure 3 Dynamic changes in the soil water contents of Potentilla parvifolia
2.2.3 土壤容重
金露梅灌丛随恢复期延长,丛间土壤容重0-10 cm土层中呈下降趋势,LR显著低于NR和IR(P<0.05);10-20 cm土层中,呈先降后升趋势,但4个恢 复 期 之 间 无 显 著 性 差 异 (P>0.05);20-30 cm土层中,也呈先降后升趋势,IR和MR无显著差异(P>0.05),MR 显著低于 NR 和 LR(P<0.05)(图 4)。
丛内土壤容重0-10 cm土层中呈下降趋势,IR、MR、LR 之间无显著差异 (P>0.05),MR、LR 显著低于 NR(P<0.05);10-20 cm 土层中,表现为 MR <IR≈ LR<NR,差异显著 (P<0.05);20-30 cm 土层 中 , MR 显 著 小 于 NR 和 LR(P< 0.05), NR、IR、LR 无显著差异 (P>0.05)(图 4)。
2.3 土壤养分含量变化
2.3.1 土壤有机质的变化
随着金露梅灌丛的逐渐恢复,丛间各层土壤有机质含量均呈先升后降的单峰曲线,0-10 cm土层中,峰值为 MR, NR和 LR显著低于 MR、IR(P< 0.05);10-20 cm 土层中,峰值为 IR, MR 和 LR 显著低于 NR、IR(P<0.05);20-30 cm 土层中,表现为IR>NR>MR>LR(表 2)。
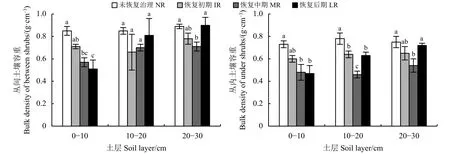
图 4 金露梅灌丛土壤容重动态变化Figure 4 Dynamic changes in the soil bulk density of Potentilla parvifolia
随着金露梅灌丛的逐渐恢复,丛内各层土壤有机质含量呈先升后降的单峰曲线,峰值均为IR。0-10 cm 土层中,NR、LR 显著低于 IR、MR(P<0.05);10-20 cm土层中,MR、LR显著低于 NR、IR(P<0.05);20-30 cm土层中,LR显著低于 NR、IR、MR(P<0.05)(表 3)。综上,除表层土壤有机质含量保持一致外,深层土壤有机质含量在LR降低很明显,与NR相比,约降低64.2%。
2.3.2 土壤全氮和速效氮的变化
随着金露梅灌丛的逐渐恢复,丛间土壤全氮含量0-10 cm土层中,MR、LR显著低于NR、IR(P<0.05);10-20 cm土层中,IR、MR、LR均显著低于 NR(P<0.05);20-30 cm 土层中,LR 显著低于NR、IR、MR(P<0.05)(表 2)。丛内土壤全氮含量0-10 cm土层中,恢复期之间无显著性差异(P>0.05);10-20、20-30 cm土层中,全氮含量均呈降低趋势 (表 3)。
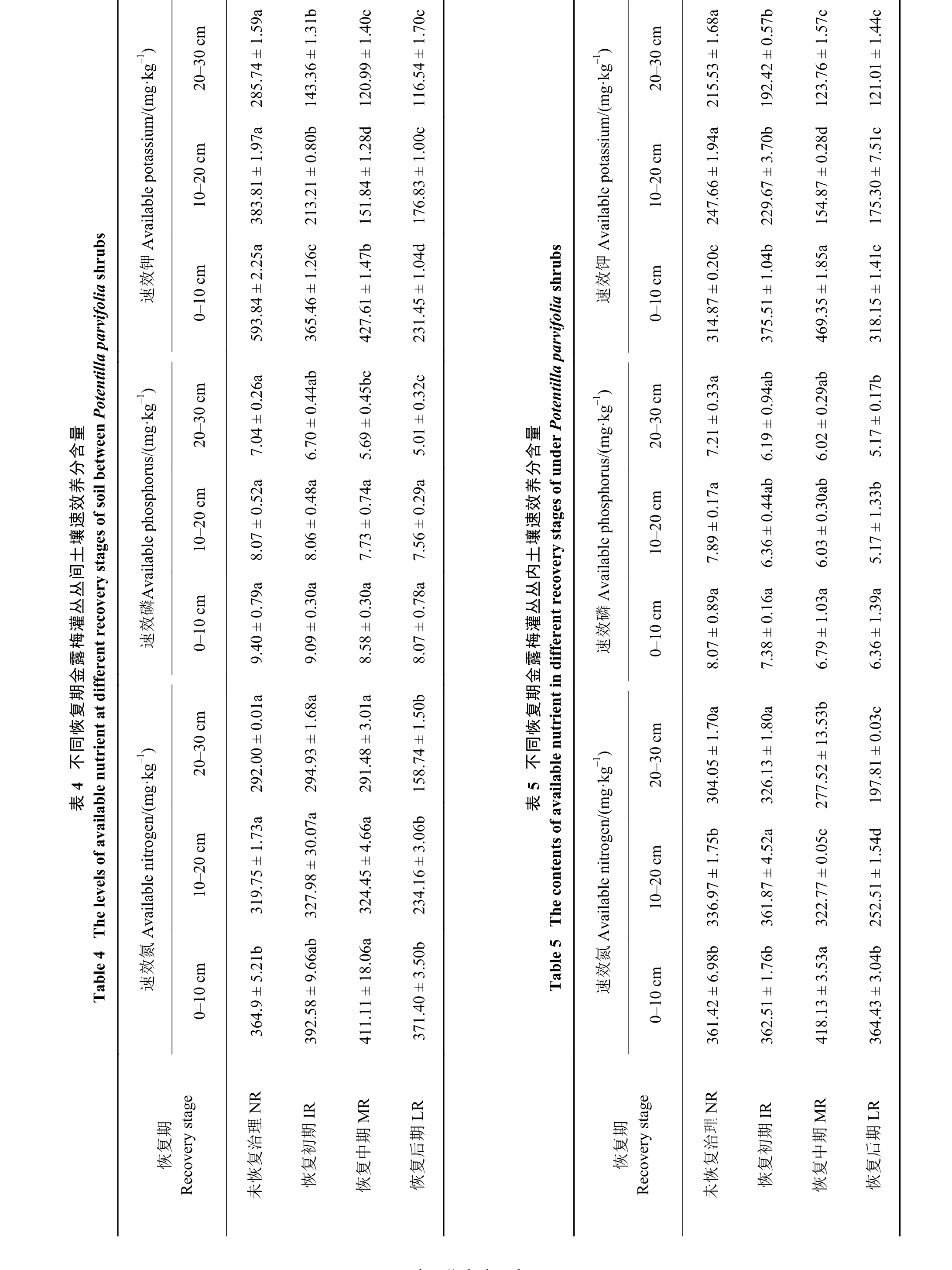
随着金露梅灌丛的逐渐恢复,丛间土壤速效氮含量呈先升后降的单峰曲线,0-10 cm土层中峰值为 MR, NR、LR 显 著 低 于 MR(P<0.05);10-20、20-30 cm土层中,峰值为IR, LR显著低于NR、IR、MR(P<0.05),后三者差异不显著 (P>0.05)(表 4)。丛内土壤速效氮含量0-10 cm土层中,MR显著高于 NR、IR 和 LR(P<0.05);10-20 cm 中,表现为IR>NR>MR>LR,差异显著(P<0.05);20-30 cm 中,MR、LR 显著低于 NR、IR(P<0.05)(表 5)。
2.3.3 土壤全磷与速效磷的变化
金露梅灌丛土壤全磷含量在丛间0-10 cm土层中,MR 显著高于 NR、IR、LR(P<0.05);10-20、20-30 cm土层中,IR显著高于 NR、MR、LR(P<0.05)(表 2)。丛内土壤全磷含量 0-10、10-20 cm土层中,MR 显著低于 NR、IR、LR(P<0.05);20-30 cm 中,MR、LR 显著低于 NR、IR(P<0.05)(表 3)。
金露梅灌丛土壤速效磷含量表现为NR>IR >MR>LR。丛间土壤 0-10 cm、10-20 cm 土层中,4 个恢复期之间无显著性差异 (P>0.05);20-30 cm土层中,LR 显著低于 NR、IR(P<0.05)(表 4)。丛内土壤0-10 cm土层中,恢复期之间无显著性差异 (P>0.05);10-20、20-30 cm 土层中,LR 显著低于 NR(P<0.05)(表 5)。
2.3.4 土壤全钾和速效钾的变化
土壤全钾含量表现为 NR>IR>LR>MR。丛间土壤0-10 cm土层中,恢复期之间差异显著(P<0.05);10-20、20-30 cm中,IR和LR显著低于NR(P<0.05),显著高于 MR(P<0.05)(表 2)。丛内土壤0-10 cm土层中,NR和IR显著高于MR、LR(P<0.05);10-20、20-30cm土层中,MR显著低于NR、IR、LR(P<0.05)(表3)。
丛间土壤速效钾含量0-10 cm土层中表现为NR>MR>IR>LR,差异显著 (P<0.05);10-20 cm土层中表现为:NR>IR>LR>MR,差异显著 (P<0.05);20-30 cm土层中,MR、LR显著低于NR、IR(P<0.05)(表 4)。丛内土壤速效钾含量 0-10 cm 土层中,MR 显著高于NR、IR、LR(P<0.05);10-20 cm土层中,表现为 NR>IR>LR>MR,差异显著 (P< 0.05);20-30 cm土层中,NR显著高于 IR、MR、LR(P<0.05)(表 5)。

3 讨论
碳是生命物质中的主要元素之一,是有机质的重要组成部分。草地是个巨大的碳库,在中国陆地生态系统中扮演着重要角色。方精云等[21]研究表明目前中国草地生物量碳密度在294.00~348.10gC·m-2,中国草地生态系统碳库大小为29.10PgC,还迫切需要研究中国陆地生态系统固碳潜力及植物碳库动态变化,尤其是灌丛生态系统碳储量动态变化。本研究表明,从1997年至2018年金露梅灌丛系统碳储量从 1 502.28g·m-2增长到 2 944.22g·m-2,增大2倍左右,随着金露梅灌丛恢复期的延长,灌木的碳储量呈增长趋势,而灌丛下草本层碳储量逐渐降低,但灌丛密度越大,其贡献也越大,使整个系统碳储量呈上升趋势,这与李红琴等[4]报道的金露梅灌丛植被主要以草本固碳为主的观点相悖。金露梅各器官碳储量和灌木层平均碳储量均增大,其主要由灌丛面积和生物量增加,即灌丛恢复引起的,这是我国陆地生态系统碳储量增加的主要原因[22]。本研究包括了灌木层和草本层,使对灌丛生态系统碳储量的估算更为准确。若灌丛密度小,则草本占有较多,其对系统碳储量的贡献不容疏忽,而赵串串等[12]的研究仅限于灌木层,忽略了草本层,在一定程度上低估了灌丛生态系统的碳储量。
土壤呼吸是土壤中碳返还大气的主要途径,也是目前全球大气中碳增加的一个重要来源[23]。土壤呼吸会受诸多因素的影响,如土壤有机物、微生物、基质和环境状况等[24]。本研究表明,丛间土壤呼吸速率随金露梅灌丛恢复期的延长而逐渐升高,丛内土壤呼吸则有下降趋势,其主要原因归结于土壤有机质,丛内凋落物量大,有机质含量较高,抑制了土壤呼吸[25]。其次,金露梅灌丛的冠幅越来越大,丛内的总辐射[26]远不如丛间,使丛内土壤呼吸速率降低,丛间升高。金露梅灌丛呈进展演替,灌丛的面积逐渐大于丛间草地的面积,整个系统的土壤呼吸却基本保持不变。本研究发现,0-10 cm土层容重呈下降趋势,且小于1.0,说明土壤较疏松,孔隙度较大,具有良好的持水性,这与张平等[27]研究的毛管孔隙度越大,土壤中有效水的贮存容量越大的结果相一致。
土壤有机质是指示土壤健康的关键指标,其含量受土地利用方式、人为耕作管理措施等因素的影响[28-29]。本研究发现,随植被恢复期的延长,金露梅灌丛生物量增大,凋落物量也随之增多,凋落物分解,使土壤有机质含量增多,但生物量增大到368 g·m-2左右时,有机质含量却有下降趋势,这与胡克林等[30]研究认为随着植物产量的不断提高,部分地区的有机质入不敷出,呈下降趋势的结果相一致。土壤中的氮素主要来源于土壤表层植物残体和土壤动物残体及微生物等有机质的积累[31]。植被覆盖可有效减少土壤全氮的流失,其减少作用随覆盖度的增大而增加[32],这与Weier[33]研究结果相一致。土壤全磷含量与有机碳消涨趋势是一致的[34]。本研究中,金露梅灌丛丛间土壤全磷含量随土层深度增加而减少,灌丛内土壤全磷含量则随土层深度增加呈先减少后增加趋势。在不同的覆被类型下,土壤全磷含量随土壤深度增加而增加或减少[35]。本研究与土壤全磷含量随植被恢复时间的增加而增加[35-36]的研究结果相悖。因为,金露梅灌丛持续恢复,全磷包括其他部分养分含量增加到一定程度不再继续增加,反而有降低趋势。
土壤速效养分是植物生长发育所必需的基本元素直接来源,其含量变化规律与土壤类型、地形地貌及成土母质的分布情况有一定的联系[37]。而对于磷,Osono和Takeda[38]研究发现,木质素与磷的质量比率高时,磷会被固定在枯枝落叶中,当比值小时,磷将从枯枝落叶中释放以增加土壤养分含量。本研究结果表明,金露梅灌丛经过长时间的恢复,丛内枯枝落叶将大量积累,枯落物增加,导致部分磷固定在枯枝落叶中而未被释放,造成土壤速效磷含量很低,与周厚诚等[36]认为速效磷含量在各群落中皆表现出随土壤深度增加而降低的趋势,且降低速率较小的观点相符,而与Jiao等[39]研究的有效磷含量与有机质和全氮含量具有显著正相关的结果相悖。
综上,金露梅灌丛经过20年的恢复,金露梅灌丛为陆地生态系统提供了提高碳储量和土壤水分等贡献,发挥了其促进碳循环、保持水土以及涵养水源的生态功能。同时土壤有机质、全磷以及速效氮、速效磷、速效钾等养分含量有减少趋势,降低了土壤的肥沃程度,使生态系统的生产力受到影响。因此需要进一步研究灌丛合理的放牧利用率和调控机制,使灌丛和草本植物协调共生、和谐生长,提升土壤特性,优化生态环境效益,提高草牧业生产能力。
4 结论
从1997年至2018年,金露梅灌丛灌木碳储量呈增大趋势,金露梅灌丛下草本地上部分碳储量基本保持不变,地下部分碳储量呈下降趋势,灌丛群落的碳储量处于增大趋势。随着金露梅灌丛逐渐恢复,丛间土壤呼吸速率随灌丛恢复期的延长而增大,丛内土壤呼吸速率则减小;土壤含水量均呈增大趋势,0-10 cm土层容重呈减小趋势,10-30 cm土层土壤容重先减后增。金露梅灌丛丛间和丛内0-30 cm土壤有机质和速效氮含量先增后减;全磷和速效磷含量逐渐减小;全钾含量先减后增,速效钾含量总体呈减小趋势。
